预约演示
更新于:2026-01-30

Avery Biomedical Devices, Inc.
更新于:2026-01-30
概览
关联
100 项与 Avery Biomedical Devices, Inc. 相关的临床结果
登录后查看更多信息
0 项与 Avery Biomedical Devices, Inc. 相关的专利(医药)
登录后查看更多信息
6
项与 Avery Biomedical Devices, Inc. 相关的新闻(医药)2025-12-26
0:14
大家好。我叫 David Allis(大卫·阿利斯)。我在纽约的洛克菲勒大学担任教授并领导实验室,我们实验室研究染色质生物学与表观遗传学。我很高兴能参与 iBiology 这个精彩的系列讲座。
0:29
希望今天在我这一到两场讲座里,能在某些方面让你有所收获。什么是表观遗传学(epigenetics)?我们为什么要关心它?以及,关于表观遗传学究竟有什么可能是真的——以至于“决定我们的不只有 DNA 本身”?我们的遗传信息是否不止 DNA?我想我们很多人从小都接受这样的观念:
0:49(标题:关于遗传的一个突破性发现)
真正的遗传学,牢牢建立在 DNA 分子之上。
0:54
我用这张幻灯片“暴露一下年龄”:在 1944 年的洛克菲勒大学,人们甚至还不知道 DNA 就是遗传信息的宏分子载体。
1:06
这项工作由 Avery、MacLeod 和 McCarty 完成。幻灯片左侧是教科书式的观点:基因、DNA 分子(沃森和克里克描述其结构的分子)构成了遗传的基础。
1:31
你可以看到黄色框里:基因上游存在一些定义明确的 DNA 元件——我们通常称之为转录因子、转录调控因子——它们可以是正向或负向的调控,向基因“下指令”,让基因要么表达、要么沉默。
1:49
所以这里的想法是:难道基因(DNA)就是全部吗?
1:55(标题:人类基因组计划)
2001 年的一项里程碑式成果,是对整个人类基因组进行测序。
2:03
它得到了大量关注(也理所应当),因为我们第一次获得了从碱基到碱基的“蓝图信息”,也就是遗传信息。人类基因组计划之所以令人兴奋、引起巨大关注,有很多原因:
2:24
基因也许决定(或至少强烈影响)性状——对像我这样年纪不小的人来说,弄清楚它也许能帮助我们改善一些问题;基因决定疾病——这关系到很多人;此外,遗传分析可能帮助我们诊断疾病、并潜在发展治疗策略,也就是今天所说的个体化医疗或精准医疗。
2:46
这些都是人类基因组计划带来的好处。
2:53
但与此同时,人类基因组计划也留下了一些“挥之不去”的问题,我想用它们来为表观遗传学铺垫:
3:01
第一,测出来的人类基因数量出乎意料地少——大约只有 21,000 个。看起来不少,但实际上偏少,尤其当你想到:青蛙、鱼、蠕虫的基因数也差不多。直觉上很难接受我们竟然不需要比这些“生物小家伙”更多的基因。
3:28
第二,个体差异从何而来?尤其是一对双胞胎:如果同卵双胞胎在遗传上完全一致,但外貌或表型并不完全相同,那么差异来自哪里?
3:46
第三,在人的一生中,我们受到环境、饮食等外界因素影响——这些会不会影响“基因组”的功能输出?从严格意义上说,基因组序列相对固定,这似乎无法解释这种层面的变化。
4:07
第四,当我们用越来越精准的方式去研究人类疾病时,是否每一种情况都能找到一个明确的遗传损伤/突变来解释它?如果答案是:我没找到错误、没找到序列改变——那还能用什么性质来解释?
4:26(标题:表观遗传“景观”:基因 + 环境 = 表型)
这就引出了我第一讲的主题:表观遗传学。
4:33
这个词由 Conrad Waddington(康拉德·瓦丁顿)在 1942 年提出。他因一张著名的“地形图”而广为人知:一个小球沿着山坡滚落。作为发育生物学家,他关心的问题是:为什么在胚胎早期,同一套基因组可以产生不同的细胞类型?
4:55
小球沿着地形图左侧或右侧滚下,最终落在不同的“沟槽”里;他设想这就对应不同的外在表现、不同的表型——但关键是:都来自同样的遗传物质。
5:09
“epi-”这个希腊前缀意为“在……之上”“附加于……”,也就是在遗传学之上、或在遗传信息之外“再加一层”的东西。
5:24(标题:什么是表观遗传学?为什么它令人兴奋?)
我给你一个简化理解的方法:把一个基因想成一个英文单词——那是遗传学,是“字母本身”。
5:37
但在英语里,同样的单词你可以用不同方式读:轻声读、重读;加粗、下划线,意义会改变。字母没变,单词没变,但“读法”变了,意义就变了——这就像表观遗传。
6:01
如果你更偏技术一点:基因像电脑硬件,表观遗传像电脑软件。硬件一样,但你往系统里写入的“程序”不同,最终结果就不同。
6:14
这是一张 2011 年 ChemBioChem 封面图,画了一个拼图,想表达表观遗传学与生物学众多领域之间令人兴奋的联系。你可以挑自己喜欢的拼图块:发育生物学是表观遗传学的起点;而疾病(左下角的白色拼图块)同样重要——尤其是那些无法仅靠遗传突变解释、却可能由表观遗传异常解释的疾病。
7:02
另一个比喻:快到情人节了,来个蛋糕。蛋糕本体可以一样,但你在上面抹不同的糖霜/奶油(含糖、不含糖等),味道就会大不一样。蛋糕没变,但“覆盖层”改变了风味。糖霜通常就在蛋糕上面——这也呼应“epi-”的含义:在遗传之上再加一层。
7:33(标题:Epi-genetics:基因组之外“附加的一层”)
回到真实生物学:一个早期胚胎的细胞,最初都还“未定向”,并不清楚将来要变成什么。随着发育推进,同一套基因组会分化出不同细胞类型:血液、神经、骨、皮肤等。
7:57
一旦分化完成,细胞“知道自己是谁”,并在分裂时把身份传给子代细胞:血细胞产生血细胞,等等。
8:15
当然,我们也知道干细胞是很热门的领域:体内保留了一小群“储备细胞”,具有多能性,仍可走向不同命运。另一个热点问题是:能否把一个完全分化的细胞重新编程,让它“倒回去”,回到更具多能性的状态?
8:39
但我想强调的是:无论是什么细胞类型,它们拥有的遗传物质(DNA序列)是相同的。
8:47
而且很遗憾的是,当你“重编程”或在这层表观基因组(epigenome)上发生错误时,细胞可能被推回到更未分化的状态,增殖能力增强。现在已经有大量例子显示,这类变化会让细胞呈现癌样表型——癌症当然是非常可怕的疾病之一。
9:13
那么,如何从机制层面真正理解表观遗传,而不只是细胞生物学现象?
9:27
这张图可能让你觉得我疯了(笑)。这是我以前实验室成员 Sean Taverna(后来在约翰·霍普金斯医学院任教)画的漫画。DNA 分子像刚洗完澡出来,面临选择:拿一条写着 “histone(组蛋白)” 的毛巾,还是拿一条写着 “herstone” 的毛巾。看起来大自然选择了 histone。【注:这里是英文双关,her + stone 模仿 histone 发音的玩笑】
我想表达的是:我说的“histone(组蛋白)”究竟是什么?
10:02(标题:染色质是基因组的生理存在形式)
如果你看细胞核的示意图,高等生物的 DNA 并不是“裸露的 DNA”,而是被蛋白质包装起来的。
10:16
它看起来像“串珠”一样:基本重复单位称为核小体(nucleosome)。核小体的核心是一组组蛋白,作为支架,DNA 大约绕着它缠绕近两圈。
10:36
这些“珠子串”还会进一步卷曲形成更高阶的结构(具体细节至今仍在研究中)。
10:43
如果你把它想成弹簧玩具(Slinky):当弹簧被拉开、染色质更“开放/去凝聚”时,基因更容易表达;相反,当弹簧被压紧、染色质高度致密时,基因更难表达。
11:10
即使是转录因子,在致密环境下也更难找到其 DNA 靶序列并发挥作用。
11:23(标题:电镜下的细胞核:常染色质 Eu 与异染色质 Het)
我也得给你看一张真实的细胞核:这是人细胞核的电镜图像,通过染色能看到致密与去凝聚的染色质。
11:32
你会看到核内有一些斑块状、非常致密的区域——称为异染色质(heterochromatin);其间散布着更开放、去凝聚的区域——称为常染色质(euchromatin)。
11:49
这就是一个真实的人细胞核。
11:57(标题:在染色质背景下将基因在“开/关”之间切换)
在某些条件下,你甚至可以把染色质从细胞核里“抽离/拉出来”(原转写里“flute it”应为“pull/tease it out”之意),在核外看到这些纤维。
12:11
下图可以看到经典的“串珠样”结构,像珍珠项链。相对开放的常染色质状态,似乎可以通过某种“分子开关”,被切换成上图那种更凝聚、更致密的状态(异染色质)。
12:32
因此对我们这种生化实验室来说,关键问题是:这个分子开关是什么?它的靶点是谁?是什么在控制常染色质与异染色质的切换?
12:51(标题:第二个突破性发现——组蛋白会发生化学修饰)
这就引到洛克菲勒的另一个开创性发现。我要致敬 Vince Allfrey。
13:03
这张图展示核小体的原子结构:不同颜色表示不同组蛋白,DNA 绕在外面。你还能看到从核小体伸出来的“尾部”(histone tails)。
13:24
Allfrey 在 1964 年提出:这些尾部会被加上化学修饰/化学标记,它们可以充当“开关”或“信号”,指示细胞让基因处于开启或关闭状态。
13:47
这就是他的设想:这些化学修饰可能就是我们刚才说的“开关”。
14:01
而任何化学修饰都需要酶系统去“添加”这个标记——我们把它们叫作 writers(书写者);也需要相反的酶系统去“去除”这些化学基团——我们叫作 erasers(橡皮擦)。两者对抗,形成开/关调控。
14:22
这是 1964 年提出的假说,距今已超过半个世纪。
14:29(标题:谨慎选择“模式系统/模式生物”)
那怎么找到这些酶?你可能不会想到这种模式生物。
14:36
上图是纤毛虫(ciliate)的一种示意图;下图是四膜虫(Tetrahymena)的真实染色照片。纤毛虫有个很独特的特性:它们一生中有两个细胞核。
15:04
大的那个叫大核(macronucleus),主要负责基因表达,它的染色质更像大量常染色质的集合。
15:16
小的那个叫小核(micronucleus),图中红色标出,非常致密、完全转录沉默。小核几乎不做转录,它主要保留用于性生殖周期相关的遗传传递。
15:50
我做博士后时对这个系统很着迷,虽然研究一个这么“奇怪”的生物看起来很另类。
15:58
我给你们看一张我当年的凝胶图(非常“史前”)。我尝试把大核和小核做生化分离,我加入的 Martin Gorovsky 实验室建立了这些方法。
16:17
凝胶上左侧是大核的组蛋白,右侧是小核的组蛋白。你不用理解凝胶细节,只要注意:大核组蛋白显示出更明显的“梯状/阶梯状”条带(我用绿点标出)。而小核里这种梯状形式少得多(虚线框处基本看不到)。
17:13
这些“梯子”来自电荷差异;每一级“阶梯”代表多加了一个化学基团——这正是 Allfrey 当年设想的化学修饰,其中一个重要的就是乙酰基(acetyl group)。
17:31
这给了我思路:我可以纯化大核,去分离负责把乙酰基加到组蛋白上的“writer”。
17:48
我建立了自己的实验室,吸引了几位勇敢的学生,其中之一是 Jim Brownell。
17:54(标题:活性凝胶检测)
这是 Jim 博士论文里的内容。他的想法非常巧妙:做一个“凝胶内活性测定”。
18:07
他把组蛋白(我们一直在讲的蛋白)“掺进”凝胶里(图中用蓝色表示;当然组蛋白本身不是真的蓝色)。作为对照,他还做了另一块同样的凝胶,但掺入的是非组蛋白蛋白质。
18:42
然后把四膜虫大核提取物上样电泳。在理想情况下,如果凝胶经过一系列处理(细节略),再加入反应所需的放射性辅因子——放射性乙酰辅酶 A(acetyl-CoA;乙酰基来自代谢)——孵育、洗涤后,就能在自显影里看到酶所在的位置:一个红色条带(箭头所指)。对照凝胶中则不应出现。
19:35
这是 Jim 第一次尝试就得到的结果:左边(含组蛋白的凝胶)出现了很强的条带;右边对照凝胶(他用的是 BSA,牛血清白蛋白,不是组蛋白)没有强条带。这说明“组蛋白 writer”不会对 BSA 起作用。
20:07
Jim 的厉害之处在于:他从 200 升四膜虫培养物中纯化出足够的 p55 条带蛋白,完成测序并克隆了基因。
20:20(标题:可逆乙酰化“开/关开关”——1996)
这成为我们实验室最知名的工作之一。Jim 那条神秘条带,最终被证明是组蛋白乙酰转移酶(histone acetyltransferase, HAT)。
20:34
有趣的是:在酵母里,这个蛋白原本就被认为是正向转录调控因子,但当时没人知道它是一个酶、并且具有 HAT 活性。这个发现极大推动了整个领域:大家开始认真对待“染色质这一层”的问题。
21:01
Jim 的论文发表于 1996 年 3 月。几乎同时(一个月后),哈佛大学 Stew Schreiber 团队(研究生 Jack Taunton)用非常巧妙的化学策略发现了能把乙酰基“去掉”的酶:组蛋白去乙酰化酶(histone deacetylase, HDAC)。
21:29
更令人惊讶的是:这个蛋白在酵母中早已被认为是转录共抑制因子(co-repressor)。于是“魔法发生了”:一个被认为促进转录的蛋白其实在给组蛋白写入乙酰化;一个被认为抑制转录的蛋白其实在擦除乙酰化。
21:57
因为这项工作是我们最为人熟知的内容之一,我想把真正的 Jim Brownell 介绍给你们:左边是“日常版”,他在我实验室里总戴着帽子进冷室——因为他追的是一个叫 HAT 的东西,而冷室也很冷(笑)。右边是他 1997 年获奖的样子:他赢得了“世界最佳博士论文国际奖”,被邀请去瑞典斯德哥尔摩与诺奖得主交流。我想问你:你觉得 Jim 哪边看起来更自在?我觉得他明显更适合当实验室里的“实验鼠”,而不是领奖台上的那种(笑)。
22:46
顺便一提:Jim 的 Cell 论文发表在 1996 年 3 月 22 日——那天刚好是我的生日,所以我觉得很酷。
22:59(标题:不同于 DNA 突变,表观遗传错误是可逆的,因此提供了大量有前景的药物靶点)
来快速总结一下。左边是经典遗传学:DNA 分子是遗传的核心,序列相对固定,不易被改变;基因调控因子通过识别 DNA 序列实现激活或抑制。
23:32
而我讲的是右边:更灵活、更流动的表观遗传层。各种化学基团可以被添加、也可以被去除,使基因组能快速响应外界变化。
23:53
关键差异在于:若 DNA 上发生遗传性突变(来自父母、可遗传给后代),我们往往缺乏有效办法去“修复序列”。如果它致病,我们就很被动。
24:11
但如果是表观遗传“景观”被错误设置,理论上可以通过干预把它“纠正回来”,因为 DNA 模板本身并未突变。问题在 DNA 之上。
24:32
因此,当 writers/erasers 被识别后,许多实验室开始将它们作为药物研发靶点。
24:41
其中一个非常成功的例子来自我现在所在的洛克菲勒大学“街对面”的纪念斯隆-凯特琳癌症中心(Memorial Sloan-Kettering):Paul Marks 团队开创性地把表观遗传药物用于临床试验。
24:48(标题:表观遗传癌症治疗:在治疗后数周逆转人群中的错误)
这是一个病人:左边是治疗前的胸腔影像,两张不同扫描。箭头指向胸腔内很大的肿瘤团块——这是喉癌转移至肺。
25:19
右边是治疗 8 周后同一病人的影像:肿瘤团块几乎消失或明显空泡化,对病人来说显然改善很多。
25:40
这些令人鼓舞的结果推动 Marks 实验室把工作发展到最终形成首批 FDA 批准、用于特定癌种治疗的表观遗传药物之一。
25:53(标题:癌症表观遗传学:在人体中逆转错误)
这里还有一位女性皮肤癌患者:上图是治疗前;下图是接受这种“表观遗传抑制剂/表观遗传药物”(epi-inhibitors)治疗后。视觉上很容易看出病灶显著改善。
26:10
这也在全球范围内引发对这些酶作为强大药物靶点的兴趣与深入研究。
26:24(标题:肿瘤学中的表观遗传靶点:组蛋白修饰靶点)
总结到这里:无论你最喜欢哪种修饰,都可以把它理解为一门“语言”。有 writers、有 erasers;现在我们还知道有些修饰可以被 **readers(读取模块)**识别——它们读到标记后,会结合到组蛋白尾部,从而引发染色质开放或关闭。
26:53
而且就当下而言,连这些 readers 也已经被成功开发出药物。
27:00
所以,“读、写、擦”三类环节都成为有吸引力的药物靶点,其中一些已经进入临床应用。
27:13(标题:遗传学如何揭示表观遗传在工作:果蝇位置效应斑驳 PEV)
我前面主要讲了激活相关的系统;现在讲沉默系统。Allfrey 在 1964 年还提出过另一类修饰:比如甲基化(methyl)。
27:38
乙酰化常被视为“开启”的标记;那甲基化呢?领域最初是怎么对甲基化产生兴趣的?一个经典的表观遗传现象提供了答案:不改变 DNA,却改变表型。
27:58
这是果蝇的眼睛。右侧两只果蝇眼睛颜色不同:一个很红,一个很白。发生了什么?
28:08
果蝇有个基因叫 white gene(白基因),位于 X 染色体末端。果蝇研究者常按突变表型给基因命名:white 基因正常活跃时反而会让眼睛变红(这确实有点反直觉)。
28:33
当染色体被 X 射线处理后,white 基因在不改变序列的前提下被“搬到”异染色质附近,它就会被沉默。
28:51
这种现象叫 位置效应斑驳(position-effect variegation, PEV):基因换了“邻居”,序列没变,但眼睛从红变白;有时呈现斑驳不均,因此叫 variegation。
29:14
由于这是果蝇系统,研究者可以筛选突变体:哪些突变会让 PEV 不容易发生。我要讲其中一个对表观遗传领域极其重要的突变:Su(var)3-9。
29:31
Su(var)3-9 的含义是“抑制斑驳(suppressor of variegation)”的突变。3-9 表示它定位在第 3 号染色体、突变体编号 9。关键不是编号,而是它的分子功能是什么。
30:09
我职业生涯里最喜欢的故事之一:右下角这位科学家 Thomas Jenuwein 获得证据表明,小鼠的 Su(var)3-9 同源蛋白可能是一种“甲基转移酶 writer”。他想与我们合作验证。
30:39
我们用类似于当年研究乙酰化 writer 的思路,与 Thomas 合作。这里展示的是我办公室的黑板(直到今天还在)。结论是:哺乳动物 Su(var)3-9 的确是甲基转移酶(methyltransferase)。
30:57
它的靶标是组蛋白 H3 的第 9 位赖氨酸(lysine 9)。也就是 H3K9。简直不可思议——这个基因从名字到功能居然“早就写对了”,尽管当年给它命名的遗传学家并不知道它到底做什么。
31:25(标题:逻辑相似,但“开/关”由两套系统实现)
现在人们通常这样理解:
31:31
(1)开启基因(ON)的一种典型方式:
用 HAT(组蛋白乙酰转移酶)把乙酰基写到组蛋白上;写上之后,会有蛋白质(读者,图中 X)识别乙酰化并结合,促进基因激活;当要关闭时,用 HDAC 把乙酰基擦掉。
32:17
(2)沉默基因(OFF)的一种典型方式:
先确保该赖氨酸位点没有乙酰化(可先用 HDAC 去乙酰化);然后由 HMT(组蛋白甲基转移酶,如 Su(var)3-9)写入甲基;接着读者蛋白(图中 Y)读到甲基标记后,促使染色质压缩并沉默。
33:04
当年做这张图时我们还不知道有没有“去甲基化酶”;后来 Yang Shi 等人发现甲基标记也能被酶促去除,所以这张图其实也已经有点过时。
33:22
但总体而言:乙酰化常与“开启”相关,甲基化常与“关闭”相关(当然具体取决于位点与修饰类型)。
33:36
下面给几个生物学例子。
33:43
这张漫画里,妈妈爸爸在忙,姐姐在逗弟弟:她说自己的表观基因组(epigenome)更复杂。某种意义上她说得对:女性有两条 X 染色体(两条粉色 X),男性是 XY。
34:07
自然界如何在 X 染色体剂量上实现平衡?答案是:在早期发育中把女性两条 X 里的其中一条关闭,这叫 X 染色体失活(X inactivation)。
34:25(标题:表观遗传沉默的“相同基因组”例子:三花猫)
你可能觉得我在编,但实验能看到:把女性哺乳动物细胞染色,如果用“甲基化相关的组蛋白抗体”染色,会看到局部红色亮斑——那就是被完全关闭、异染色质化的失活 X。
34:50
在男性细胞里做同样染色,一般看不到这种现象。
34:55
顺便说:你如果见过三花猫(右图),毛色斑块就是女性 X 失活的结果:与毛色基因相关的那条 X 在不同细胞群里随机失活,早期发育就决定了不同区域的毛色,所以三花猫几乎都是雌性。
35:30
我提到同卵双胞胎:他们遗传上完全相同,但现实中往往并不完全一样。研究发现:同卵双胞胎在表观基因组层面并不完全一致,因此成为“遗传相同、表观遗传不同”的很好的模型。
36:05
再往前一步:我小时候以为,如果我做些对自己不好的事(比如不良生活方式),至少只是在伤害自己;而“人如其食(we are what we eat)”也常被提起。
36:25
但表观遗传学革命后,有人把这句话改写成:“我们也在某种程度上是我们父母吃了什么的结果……甚至是更上一代的结果。”
36:32(标题:遗传完全相同的鼠,但母鼠饮食不同)
这里是一个直观例子:这些小鼠遗传完全相同,唯一变量是母鼠在怀孕期间的饮食不同(更具体地说,是会影响甲基化水平的饮食差异)。结果是:后代虽然基因相同,但表型差异巨大——体型、毛色都明显不同。这想想其实有点“吓人”。
37:22
再极端一些:Michael Meaney 团队研究更偏社会行为的问题:是否存在类似“好妈妈 vs 不那么好的妈妈”的动物模型?
37:40
他发现某些鼠系的母鼠会频繁舔幼崽、拥抱/照料更多(“好妈妈”),另一些则相反(“不怎么照料”)。
38:01
他进一步研究:把遗传相同的幼崽暴露给“好妈妈”或“差妈妈”,会发生什么?结果显示:脑区中的表观遗传标记会改变,进而影响基因表达,最终影响应激反应与可遗传的表型。
38:31
这也让人想到社会层面的现实意义:当养育环境差异巨大时,后果可能不仅是成长经历的悲剧,还可能通过表观遗传层影响下一代,甚至下一代的下一代。
38:55
这些要点你在总结图里应该已经看到了。
39:01
我想给你留下这样的印象:左侧是经典遗传学,这当然没有错——比如吸烟、过度日晒确实可能造成 DNA 突变并致病,我们当然希望从父母那里继承到“好的 DNA”。
39:26
但现在越来越清楚:遗传学并不能解释全部;基因组还存在更动态、更快速响应需求的机制层——表观遗传。
39:50
为了完整起见:不只是组蛋白会被化学修饰,DNA 本身也会(比如 DNA 甲基化),这些都是生化学家非常关注的热点。
39:58
writers、erasers、readers 这些概念,以及这些修饰与人类疾病(尤其癌症)的关联,使其成为药物开发的重点。原因之一是:基因组序列相对难“修”,表观基因组更可能被“纠正”。
40:27(标题:一本新的表观遗传学教材《EPIGENETICS》)
如果你对这个领域感兴趣……我不是在硬推书,但这是个复杂领域。以前 Cold Spring Harbor 找了少数几个人写一本教材,这是第二版,刚在去年出版。
40:49
我也想借此介绍编辑团队:最左边你可能认得 Thomas Jenuwein(我们做 Su(var)3-9 甲基化合作的那位)。此外还有 Monica Lachner、Danny Reinberg、我本人,以及 Marie-Laure Caparros。
41:10
这也说明:除了实验室里的同伴,能在全球范围内建立合作、一起做学术项目,也是很大的荣幸,而且很有趣(原转写 “run” 应为 “fun”)。
41:30
我的第一讲到此结束。最后如果不感谢很多人就太说不过去了:我展示过少数实验室成员的照片(如 Jim Brownell),但我整个职业生涯有无数杰出的学生、博士后与技术人员;许多项目也离不开重要合作者;iBiology 团队非常棒——我不习惯被拍摄,但他们对我非常友好;也感谢 Lasker Foundation 促成这次录制;感谢我的资助来源、洛克菲勒大学;更感谢各位投入时间聆听。非常感谢。
如果你希望我进一步“增强可读性”,我也可以在不改动原意的前提下:
把这一讲整理成中文讲义版(按逻辑分节:背景→问题→概念→机制→案例→临床意义→总结);
或者做一个术语表(HAT/HDAC/HMT、euchromatin/heterochromatin、PEV、H3K9、X inactivation 等)方便你复习。
2025-12-26
0:14
大家好。我叫 David Allis(大卫·阿利斯)。我在纽约的洛克菲勒大学担任教授并领导实验室,我们实验室研究染色质生物学与表观遗传学。我很高兴能参与 iBiology 这个精彩的系列讲座。
0:29
希望今天在我这一到两场讲座里,能在某些方面让你有所收获。什么是表观遗传学(epigenetics)?我们为什么要关心它?以及,关于表观遗传学究竟有什么可能是真的——以至于“决定我们的不只有 DNA 本身”?我们的遗传信息是否不止 DNA?我想我们很多人从小都接受这样的观念:
0:49(标题:关于遗传的一个突破性发现)
真正的遗传学,牢牢建立在 DNA 分子之上。
0:54
我用这张幻灯片“暴露一下年龄”:在 1944 年的洛克菲勒大学,人们甚至还不知道 DNA 就是遗传信息的宏分子载体。
1:06
这项工作由 Avery、MacLeod 和 McCarty 完成。幻灯片左侧是教科书式的观点:基因、DNA 分子(沃森和克里克描述其结构的分子)构成了遗传的基础。
1:31
你可以看到黄色框里:基因上游存在一些定义明确的 DNA 元件——我们通常称之为转录因子、转录调控因子——它们可以是正向或负向的调控,向基因“下指令”,让基因要么表达、要么沉默。
1:49
所以这里的想法是:难道基因(DNA)就是全部吗?
1:55(标题:人类基因组计划)
2001 年的一项里程碑式成果,是对整个人类基因组进行测序。
2:03
它得到了大量关注(也理所应当),因为我们第一次获得了从碱基到碱基的“蓝图信息”,也就是遗传信息。人类基因组计划之所以令人兴奋、引起巨大关注,有很多原因:
2:24
基因也许决定(或至少强烈影响)性状——对像我这样年纪不小的人来说,弄清楚它也许能帮助我们改善一些问题;基因决定疾病——这关系到很多人;此外,遗传分析可能帮助我们诊断疾病、并潜在发展治疗策略,也就是今天所说的个体化医疗或精准医疗。
2:46
这些都是人类基因组计划带来的好处。
2:53
但与此同时,人类基因组计划也留下了一些“挥之不去”的问题,我想用它们来为表观遗传学铺垫:
3:01
第一,测出来的人类基因数量出乎意料地少——大约只有 21,000 个。看起来不少,但实际上偏少,尤其当你想到:青蛙、鱼、蠕虫的基因数也差不多。直觉上很难接受我们竟然不需要比这些“生物小家伙”更多的基因。
3:28
第二,个体差异从何而来?尤其是一对双胞胎:如果同卵双胞胎在遗传上完全一致,但外貌或表型并不完全相同,那么差异来自哪里?
3:46
第三,在人的一生中,我们受到环境、饮食等外界因素影响——这些会不会影响“基因组”的功能输出?从严格意义上说,基因组序列相对固定,这似乎无法解释这种层面的变化。
4:07
第四,当我们用越来越精准的方式去研究人类疾病时,是否每一种情况都能找到一个明确的遗传损伤/突变来解释它?如果答案是:我没找到错误、没找到序列改变——那还能用什么性质来解释?
4:26(标题:表观遗传“景观”:基因 + 环境 = 表型)
这就引出了我第一讲的主题:表观遗传学。
4:33
这个词由 Conrad Waddington(康拉德·瓦丁顿)在 1942 年提出。他因一张著名的“地形图”而广为人知:一个小球沿着山坡滚落。作为发育生物学家,他关心的问题是:为什么在胚胎早期,同一套基因组可以产生不同的细胞类型?
4:55
小球沿着地形图左侧或右侧滚下,最终落在不同的“沟槽”里;他设想这就对应不同的外在表现、不同的表型——但关键是:都来自同样的遗传物质。
5:09
“epi-”这个希腊前缀意为“在……之上”“附加于……”,也就是在遗传学之上、或在遗传信息之外“再加一层”的东西。
5:24(标题:什么是表观遗传学?为什么它令人兴奋?)
我给你一个简化理解的方法:把一个基因想成一个英文单词——那是遗传学,是“字母本身”。
5:37
但在英语里,同样的单词你可以用不同方式读:轻声读、重读;加粗、下划线,意义会改变。字母没变,单词没变,但“读法”变了,意义就变了——这就像表观遗传。
6:01
如果你更偏技术一点:基因像电脑硬件,表观遗传像电脑软件。硬件一样,但你往系统里写入的“程序”不同,最终结果就不同。
6:14
这是一张 2011 年 ChemBioChem 封面图,画了一个拼图,想表达表观遗传学与生物学众多领域之间令人兴奋的联系。你可以挑自己喜欢的拼图块:发育生物学是表观遗传学的起点;而疾病(左下角的白色拼图块)同样重要——尤其是那些无法仅靠遗传突变解释、却可能由表观遗传异常解释的疾病。
7:02
另一个比喻:快到情人节了,来个蛋糕。蛋糕本体可以一样,但你在上面抹不同的糖霜/奶油(含糖、不含糖等),味道就会大不一样。蛋糕没变,但“覆盖层”改变了风味。糖霜通常就在蛋糕上面——这也呼应“epi-”的含义:在遗传之上再加一层。
7:33(标题:Epi-genetics:基因组之外“附加的一层”)
回到真实生物学:一个早期胚胎的细胞,最初都还“未定向”,并不清楚将来要变成什么。随着发育推进,同一套基因组会分化出不同细胞类型:血液、神经、骨、皮肤等。
7:57
一旦分化完成,细胞“知道自己是谁”,并在分裂时把身份传给子代细胞:血细胞产生血细胞,等等。
8:15
当然,我们也知道干细胞是很热门的领域:体内保留了一小群“储备细胞”,具有多能性,仍可走向不同命运。另一个热点问题是:能否把一个完全分化的细胞重新编程,让它“倒回去”,回到更具多能性的状态?
8:39
但我想强调的是:无论是什么细胞类型,它们拥有的遗传物质(DNA序列)是相同的。
8:47
而且很遗憾的是,当你“重编程”或在这层表观基因组(epigenome)上发生错误时,细胞可能被推回到更未分化的状态,增殖能力增强。现在已经有大量例子显示,这类变化会让细胞呈现癌样表型——癌症当然是非常可怕的疾病之一。
9:13
那么,如何从机制层面真正理解表观遗传,而不只是细胞生物学现象?
9:27
这张图可能让你觉得我疯了(笑)。这是我以前实验室成员 Sean Taverna(后来在约翰·霍普金斯医学院任教)画的漫画。DNA 分子像刚洗完澡出来,面临选择:拿一条写着 “histone(组蛋白)” 的毛巾,还是拿一条写着 “herstone” 的毛巾。看起来大自然选择了 histone。【注:这里是英文双关,her + stone 模仿 histone 发音的玩笑】
我想表达的是:我说的“histone(组蛋白)”究竟是什么?
10:02(标题:染色质是基因组的生理存在形式)
如果你看细胞核的示意图,高等生物的 DNA 并不是“裸露的 DNA”,而是被蛋白质包装起来的。
10:16
它看起来像“串珠”一样:基本重复单位称为核小体(nucleosome)。核小体的核心是一组组蛋白,作为支架,DNA 大约绕着它缠绕近两圈。
10:36
这些“珠子串”还会进一步卷曲形成更高阶的结构(具体细节至今仍在研究中)。
10:43
如果你把它想成弹簧玩具(Slinky):当弹簧被拉开、染色质更“开放/去凝聚”时,基因更容易表达;相反,当弹簧被压紧、染色质高度致密时,基因更难表达。
11:10
即使是转录因子,在致密环境下也更难找到其 DNA 靶序列并发挥作用。
11:23(标题:电镜下的细胞核:常染色质 Eu 与异染色质 Het)
我也得给你看一张真实的细胞核:这是人细胞核的电镜图像,通过染色能看到致密与去凝聚的染色质。
11:32
你会看到核内有一些斑块状、非常致密的区域——称为异染色质(heterochromatin);其间散布着更开放、去凝聚的区域——称为常染色质(euchromatin)。
11:49
这就是一个真实的人细胞核。
11:57(标题:在染色质背景下将基因在“开/关”之间切换)
在某些条件下,你甚至可以把染色质从细胞核里“抽离/拉出来”(原转写里“flute it”应为“pull/tease it out”之意),在核外看到这些纤维。
12:11
下图可以看到经典的“串珠样”结构,像珍珠项链。相对开放的常染色质状态,似乎可以通过某种“分子开关”,被切换成上图那种更凝聚、更致密的状态(异染色质)。
12:32
因此对我们这种生化实验室来说,关键问题是:这个分子开关是什么?它的靶点是谁?是什么在控制常染色质与异染色质的切换?
12:51(标题:第二个突破性发现——组蛋白会发生化学修饰)
这就引到洛克菲勒的另一个开创性发现。我要致敬 Vince Allfrey。
13:03
这张图展示核小体的原子结构:不同颜色表示不同组蛋白,DNA 绕在外面。你还能看到从核小体伸出来的“尾部”(histone tails)。
13:24
Allfrey 在 1964 年提出:这些尾部会被加上化学修饰/化学标记,它们可以充当“开关”或“信号”,指示细胞让基因处于开启或关闭状态。
13:47
这就是他的设想:这些化学修饰可能就是我们刚才说的“开关”。
14:01
而任何化学修饰都需要酶系统去“添加”这个标记——我们把它们叫作 writers(书写者);也需要相反的酶系统去“去除”这些化学基团——我们叫作 erasers(橡皮擦)。两者对抗,形成开/关调控。
14:22
这是 1964 年提出的假说,距今已超过半个世纪。
14:29(标题:谨慎选择“模式系统/模式生物”)
那怎么找到这些酶?你可能不会想到这种模式生物。
14:36
上图是纤毛虫(ciliate)的一种示意图;下图是四膜虫(Tetrahymena)的真实染色照片。纤毛虫有个很独特的特性:它们一生中有两个细胞核。
15:04
大的那个叫大核(macronucleus),主要负责基因表达,它的染色质更像大量常染色质的集合。
15:16
小的那个叫小核(micronucleus),图中红色标出,非常致密、完全转录沉默。小核几乎不做转录,它主要保留用于性生殖周期相关的遗传传递。
15:50
我做博士后时对这个系统很着迷,虽然研究一个这么“奇怪”的生物看起来很另类。
15:58
我给你们看一张我当年的凝胶图(非常“史前”)。我尝试把大核和小核做生化分离,我加入的 Martin Gorovsky 实验室建立了这些方法。
16:17
凝胶上左侧是大核的组蛋白,右侧是小核的组蛋白。你不用理解凝胶细节,只要注意:大核组蛋白显示出更明显的“梯状/阶梯状”条带(我用绿点标出)。而小核里这种梯状形式少得多(虚线框处基本看不到)。
17:13
这些“梯子”来自电荷差异;每一级“阶梯”代表多加了一个化学基团——这正是 Allfrey 当年设想的化学修饰,其中一个重要的就是乙酰基(acetyl group)。
17:31
这给了我思路:我可以纯化大核,去分离负责把乙酰基加到组蛋白上的“writer”。
17:48
我建立了自己的实验室,吸引了几位勇敢的学生,其中之一是 Jim Brownell。
17:54(标题:活性凝胶检测)
这是 Jim 博士论文里的内容。他的想法非常巧妙:做一个“凝胶内活性测定”。
18:07
他把组蛋白(我们一直在讲的蛋白)“掺进”凝胶里(图中用蓝色表示;当然组蛋白本身不是真的蓝色)。作为对照,他还做了另一块同样的凝胶,但掺入的是非组蛋白蛋白质。
18:42
然后把四膜虫大核提取物上样电泳。在理想情况下,如果凝胶经过一系列处理(细节略),再加入反应所需的放射性辅因子——放射性乙酰辅酶 A(acetyl-CoA;乙酰基来自代谢)——孵育、洗涤后,就能在自显影里看到酶所在的位置:一个红色条带(箭头所指)。对照凝胶中则不应出现。
19:35
这是 Jim 第一次尝试就得到的结果:左边(含组蛋白的凝胶)出现了很强的条带;右边对照凝胶(他用的是 BSA,牛血清白蛋白,不是组蛋白)没有强条带。这说明“组蛋白 writer”不会对 BSA 起作用。
20:07
Jim 的厉害之处在于:他从 200 升四膜虫培养物中纯化出足够的 p55 条带蛋白,完成测序并克隆了基因。
20:20(标题:可逆乙酰化“开/关开关”——1996)
这成为我们实验室最知名的工作之一。Jim 那条神秘条带,最终被证明是组蛋白乙酰转移酶(histone acetyltransferase, HAT)。
20:34
有趣的是:在酵母里,这个蛋白原本就被认为是正向转录调控因子,但当时没人知道它是一个酶、并且具有 HAT 活性。这个发现极大推动了整个领域:大家开始认真对待“染色质这一层”的问题。
21:01
Jim 的论文发表于 1996 年 3 月。几乎同时(一个月后),哈佛大学 Stew Schreiber 团队(研究生 Jack Taunton)用非常巧妙的化学策略发现了能把乙酰基“去掉”的酶:组蛋白去乙酰化酶(histone deacetylase, HDAC)。
21:29
更令人惊讶的是:这个蛋白在酵母中早已被认为是转录共抑制因子(co-repressor)。于是“魔法发生了”:一个被认为促进转录的蛋白其实在给组蛋白写入乙酰化;一个被认为抑制转录的蛋白其实在擦除乙酰化。
21:57
因为这项工作是我们最为人熟知的内容之一,我想把真正的 Jim Brownell 介绍给你们:左边是“日常版”,他在我实验室里总戴着帽子进冷室——因为他追的是一个叫 HAT 的东西,而冷室也很冷(笑)。右边是他 1997 年获奖的样子:他赢得了“世界最佳博士论文国际奖”,被邀请去瑞典斯德哥尔摩与诺奖得主交流。我想问你:你觉得 Jim 哪边看起来更自在?我觉得他明显更适合当实验室里的“实验鼠”,而不是领奖台上的那种(笑)。
22:46
顺便一提:Jim 的 Cell 论文发表在 1996 年 3 月 22 日——那天刚好是我的生日,所以我觉得很酷。
22:59(标题:不同于 DNA 突变,表观遗传错误是可逆的,因此提供了大量有前景的药物靶点)
来快速总结一下。左边是经典遗传学:DNA 分子是遗传的核心,序列相对固定,不易被改变;基因调控因子通过识别 DNA 序列实现激活或抑制。
23:32
而我讲的是右边:更灵活、更流动的表观遗传层。各种化学基团可以被添加、也可以被去除,使基因组能快速响应外界变化。
23:53
关键差异在于:若 DNA 上发生遗传性突变(来自父母、可遗传给后代),我们往往缺乏有效办法去“修复序列”。如果它致病,我们就很被动。
24:11
但如果是表观遗传“景观”被错误设置,理论上可以通过干预把它“纠正回来”,因为 DNA 模板本身并未突变。问题在 DNA 之上。
24:32
因此,当 writers/erasers 被识别后,许多实验室开始将它们作为药物研发靶点。
24:41
其中一个非常成功的例子来自我现在所在的洛克菲勒大学“街对面”的纪念斯隆-凯特琳癌症中心(Memorial Sloan-Kettering):Paul Marks 团队开创性地把表观遗传药物用于临床试验。
24:48(标题:表观遗传癌症治疗:在治疗后数周逆转人群中的错误)
这是一个病人:左边是治疗前的胸腔影像,两张不同扫描。箭头指向胸腔内很大的肿瘤团块——这是喉癌转移至肺。
25:19
右边是治疗 8 周后同一病人的影像:肿瘤团块几乎消失或明显空泡化,对病人来说显然改善很多。
25:40
这些令人鼓舞的结果推动 Marks 实验室把工作发展到最终形成首批 FDA 批准、用于特定癌种治疗的表观遗传药物之一。
25:53(标题:癌症表观遗传学:在人体中逆转错误)
这里还有一位女性皮肤癌患者:上图是治疗前;下图是接受这种“表观遗传抑制剂/表观遗传药物”(epi-inhibitors)治疗后。视觉上很容易看出病灶显著改善。
26:10
这也在全球范围内引发对这些酶作为强大药物靶点的兴趣与深入研究。
26:24(标题:肿瘤学中的表观遗传靶点:组蛋白修饰靶点)
总结到这里:无论你最喜欢哪种修饰,都可以把它理解为一门“语言”。有 writers、有 erasers;现在我们还知道有些修饰可以被 **readers(读取模块)**识别——它们读到标记后,会结合到组蛋白尾部,从而引发染色质开放或关闭。
26:53
而且就当下而言,连这些 readers 也已经被成功开发出药物。
27:00
所以,“读、写、擦”三类环节都成为有吸引力的药物靶点,其中一些已经进入临床应用。
27:13(标题:遗传学如何揭示表观遗传在工作:果蝇位置效应斑驳 PEV)
我前面主要讲了激活相关的系统;现在讲沉默系统。Allfrey 在 1964 年还提出过另一类修饰:比如甲基化(methyl)。
27:38
乙酰化常被视为“开启”的标记;那甲基化呢?领域最初是怎么对甲基化产生兴趣的?一个经典的表观遗传现象提供了答案:不改变 DNA,却改变表型。
27:58
这是果蝇的眼睛。右侧两只果蝇眼睛颜色不同:一个很红,一个很白。发生了什么?
28:08
果蝇有个基因叫 white gene(白基因),位于 X 染色体末端。果蝇研究者常按突变表型给基因命名:white 基因正常活跃时反而会让眼睛变红(这确实有点反直觉)。
28:33
当染色体被 X 射线处理后,white 基因在不改变序列的前提下被“搬到”异染色质附近,它就会被沉默。
28:51
这种现象叫 位置效应斑驳(position-effect variegation, PEV):基因换了“邻居”,序列没变,但眼睛从红变白;有时呈现斑驳不均,因此叫 variegation。
29:14
由于这是果蝇系统,研究者可以筛选突变体:哪些突变会让 PEV 不容易发生。我要讲其中一个对表观遗传领域极其重要的突变:Su(var)3-9。
29:31
Su(var)3-9 的含义是“抑制斑驳(suppressor of variegation)”的突变。3-9 表示它定位在第 3 号染色体、突变体编号 9。关键不是编号,而是它的分子功能是什么。
30:09
我职业生涯里最喜欢的故事之一:右下角这位科学家 Thomas Jenuwein 获得证据表明,小鼠的 Su(var)3-9 同源蛋白可能是一种“甲基转移酶 writer”。他想与我们合作验证。
30:39
我们用类似于当年研究乙酰化 writer 的思路,与 Thomas 合作。这里展示的是我办公室的黑板(直到今天还在)。结论是:哺乳动物 Su(var)3-9 的确是甲基转移酶(methyltransferase)。
30:57
它的靶标是组蛋白 H3 的第 9 位赖氨酸(lysine 9)。也就是 H3K9。简直不可思议——这个基因从名字到功能居然“早就写对了”,尽管当年给它命名的遗传学家并不知道它到底做什么。
31:25(标题:逻辑相似,但“开/关”由两套系统实现)
现在人们通常这样理解:
31:31
(1)开启基因(ON)的一种典型方式:
用 HAT(组蛋白乙酰转移酶)把乙酰基写到组蛋白上;写上之后,会有蛋白质(读者,图中 X)识别乙酰化并结合,促进基因激活;当要关闭时,用 HDAC 把乙酰基擦掉。
32:17
(2)沉默基因(OFF)的一种典型方式:
先确保该赖氨酸位点没有乙酰化(可先用 HDAC 去乙酰化);然后由 HMT(组蛋白甲基转移酶,如 Su(var)3-9)写入甲基;接着读者蛋白(图中 Y)读到甲基标记后,促使染色质压缩并沉默。
33:04
当年做这张图时我们还不知道有没有“去甲基化酶”;后来 Yang Shi 等人发现甲基标记也能被酶促去除,所以这张图其实也已经有点过时。
33:22
但总体而言:乙酰化常与“开启”相关,甲基化常与“关闭”相关(当然具体取决于位点与修饰类型)。
33:36
下面给几个生物学例子。
33:43
这张漫画里,妈妈爸爸在忙,姐姐在逗弟弟:她说自己的表观基因组(epigenome)更复杂。某种意义上她说得对:女性有两条 X 染色体(两条粉色 X),男性是 XY。
34:07
自然界如何在 X 染色体剂量上实现平衡?答案是:在早期发育中把女性两条 X 里的其中一条关闭,这叫 X 染色体失活(X inactivation)。
34:25(标题:表观遗传沉默的“相同基因组”例子:三花猫)
你可能觉得我在编,但实验能看到:把女性哺乳动物细胞染色,如果用“甲基化相关的组蛋白抗体”染色,会看到局部红色亮斑——那就是被完全关闭、异染色质化的失活 X。
34:50
在男性细胞里做同样染色,一般看不到这种现象。
34:55
顺便说:你如果见过三花猫(右图),毛色斑块就是女性 X 失活的结果:与毛色基因相关的那条 X 在不同细胞群里随机失活,早期发育就决定了不同区域的毛色,所以三花猫几乎都是雌性。
35:30
我提到同卵双胞胎:他们遗传上完全相同,但现实中往往并不完全一样。研究发现:同卵双胞胎在表观基因组层面并不完全一致,因此成为“遗传相同、表观遗传不同”的很好的模型。
36:05
再往前一步:我小时候以为,如果我做些对自己不好的事(比如不良生活方式),至少只是在伤害自己;而“人如其食(we are what we eat)”也常被提起。
36:25
但表观遗传学革命后,有人把这句话改写成:“我们也在某种程度上是我们父母吃了什么的结果……甚至是更上一代的结果。”
36:32(标题:遗传完全相同的鼠,但母鼠饮食不同)
这里是一个直观例子:这些小鼠遗传完全相同,唯一变量是母鼠在怀孕期间的饮食不同(更具体地说,是会影响甲基化水平的饮食差异)。结果是:后代虽然基因相同,但表型差异巨大——体型、毛色都明显不同。这想想其实有点“吓人”。
37:22
再极端一些:Michael Meaney 团队研究更偏社会行为的问题:是否存在类似“好妈妈 vs 不那么好的妈妈”的动物模型?
37:40
他发现某些鼠系的母鼠会频繁舔幼崽、拥抱/照料更多(“好妈妈”),另一些则相反(“不怎么照料”)。
38:01
他进一步研究:把遗传相同的幼崽暴露给“好妈妈”或“差妈妈”,会发生什么?结果显示:脑区中的表观遗传标记会改变,进而影响基因表达,最终影响应激反应与可遗传的表型。
38:31
这也让人想到社会层面的现实意义:当养育环境差异巨大时,后果可能不仅是成长经历的悲剧,还可能通过表观遗传层影响下一代,甚至下一代的下一代。
38:55
这些要点你在总结图里应该已经看到了。
39:01
我想给你留下这样的印象:左侧是经典遗传学,这当然没有错——比如吸烟、过度日晒确实可能造成 DNA 突变并致病,我们当然希望从父母那里继承到“好的 DNA”。
39:26
但现在越来越清楚:遗传学并不能解释全部;基因组还存在更动态、更快速响应需求的机制层——表观遗传。
39:50
为了完整起见:不只是组蛋白会被化学修饰,DNA 本身也会(比如 DNA 甲基化),这些都是生化学家非常关注的热点。
39:58
writers、erasers、readers 这些概念,以及这些修饰与人类疾病(尤其癌症)的关联,使其成为药物开发的重点。原因之一是:基因组序列相对难“修”,表观基因组更可能被“纠正”。
40:27(标题:一本新的表观遗传学教材《EPIGENETICS》)
如果你对这个领域感兴趣……我不是在硬推书,但这是个复杂领域。以前 Cold Spring Harbor 找了少数几个人写一本教材,这是第二版,刚在去年出版。
40:49
我也想借此介绍编辑团队:最左边你可能认得 Thomas Jenuwein(我们做 Su(var)3-9 甲基化合作的那位)。此外还有 Monica Lachner、Danny Reinberg、我本人,以及 Marie-Laure Caparros。
41:10
这也说明:除了实验室里的同伴,能在全球范围内建立合作、一起做学术项目,也是很大的荣幸,而且很有趣(原转写 “run” 应为 “fun”)。
41:30
我的第一讲到此结束。最后如果不感谢很多人就太说不过去了:我展示过少数实验室成员的照片(如 Jim Brownell),但我整个职业生涯有无数杰出的学生、博士后与技术人员;许多项目也离不开重要合作者;iBiology 团队非常棒——我不习惯被拍摄,但他们对我非常友好;也感谢 Lasker Foundation 促成这次录制;感谢我的资助来源、洛克菲勒大学;更感谢各位投入时间聆听。非常感谢。
如果你希望我进一步“增强可读性”,我也可以在不改动原意的前提下:
把这一讲整理成中文讲义版(按逻辑分节:背景→问题→概念→机制→案例→临床意义→总结);
或者做一个术语表(HAT/HDAC/HMT、euchromatin/heterochromatin、PEV、H3K9、X inactivation 等)方便你复习。
临床研究
2024-05-05
前言:干细胞衍生免疫细胞,为“现货型”CAR-T、CAR-NK提供稳定、大量、相对均一的细胞来源。年末,时间充裕,稍微梳理一下全球干细胞企业及其进展,发现不乏Bayer,BMS,富士胶片,诺和诺德等巨头身影。1.Allele Biotechnology and Pharmaceuticals公司简介:成立于1999年,专注于开发临床和治疗用途的前沿技术,包括RNA干扰、荧光蛋白、诱导多能干细胞(iPSCs)、基因组编辑和单域抗体。2015年,圣地亚哥建立了cGMP厂房,以进一步推进基于iPSC的细胞治疗的临床开发。Allele的创始人和CEO Dr. Jiwu Wang (王继武博士)公司网址:https://www.allelebiotech.com2.Aspen Neuroscience公司简介:2018年建立,致力于干细胞治疗神经系统疾病。创始人为The Scripps Research Institute再生医学中心,干细胞中心创始主任Jeanne Loring’教授。技术基础由Jeanne Loring教授开发,自体诱导多能干细胞(iPSCs)分化多巴胺能神经元。ANPD001目前拟递交IND,适应症:散发性帕金森病。公司网址:https://www.aspenneuroscience.com/3.Avery Therapeutics公司简介:Avery是一家致力于开发组织工程疗法,治疗疾病和人体肌肉损伤的公司。Avery主要产品MyCardia™用于治疗心力衰竭。MyCardia是一种稳定和坚固的异体工程组织移植物,可以大规模生产,冷冻保存,并在4℃运输。在心力衰竭的临床前研究中,与未经治疗的队列相比,MyCardia改善了心肌功能,并改善了日常活动和运动耐量等生活质量指标。公司网址:https://averythera.com/4.BlueRock /Bayer公司简介:2016年Bayer与Versant Ventures 合资成立BlueRock Therapeutics,2019年8月8日,拜耳公司(Bayer AG)和BlueRock Therapeutics联合宣布了一项协议。根据该协议,拜耳将完全收购BlueRock Therapeutics。BlueRock Therapeutics专注于利用专有的诱导性多能干细胞(iPSC)平台开发神经病学、心脏病学和免疫学领域的工程细胞疗法。管线进展MSK-DA01:临床1期(NCT04802733),治疗晚期帕金森氏病,2022年1月第一个病人用药2021年5月,BlueRock、富士细胞和Opsis治疗公司建立了研发联盟,开发异基因ipsc衍生的眼病细胞疗法。公司网址:https://bluerocktx.com/5.Bone Therapeutics公司简介:位于比利时戈斯利,致力于将未分化的干细胞转化为“成骨细胞”,这些细胞可以通过微创手术使用,避免侵入性手术。临床重点开发ALLOB®,这是一种来自健康供体干细胞的异体“现货型”细胞治疗产品,目前正在进行II期研究,用于治疗延迟愈合骨折。公司网址:https://bonetherapeutics.com/6.Brooklyn Immuno Therapeutics(纳斯达克:BTX)公司简介:Brooklyn Immuno Therapeutics(BTX)是一家临床阶段的生物制药公司,使用最新技术,如合成mRNA、细胞重编程、基因编辑、细胞因子开发等,推动正在进行的项目,包括iPSC衍生的MSC(iMSC)、基因编辑的iMSC和体内基因编辑。细胞因子药物IRX-2,正在进行2B期临床研究,适应症头颈部鳞癌。公司网址:https://brooklynitx.com7.Cellectis(纳斯达克:CLLS,和泛欧交易所增长市场:ALCLS)公司简介:使用其注册的TALEN® 技术,产生UCART (Universal Chimeric Antigen Receptor T-cells)。Cytovia Therapeutics 和 Cellectis基于TALEN®,通过iPSCs诱导产生NK细胞,进行细胞治疗,这部分合作由再凌生物CytoLynx Therapeutics(Cytovia Therapeutics中国合资公司)完成。公司网址:https://www.cellectis.com/8.Cellino Biotech(AI驱动的干细胞疗法公司)公司简介:Cellino使用机器学习、人工智能和激光技术来使细胞治疗制造变得自动化。细胞是在一个封闭的盒式格式中创建的,这允许数千个患者样本在一个单一的设施中并行处理,填补干细胞行业的一个关键空白,即无法大规模制造个性化的细胞。Bayer投资这家初创公司8000万美金。公司网址:https://www.cellinobio.com/9.Century Therapeutics公司简介:Century是一个创新的生物技术公司,位于西雅图,集成前沿基因编辑、蛋白质工程生成异基因iPSC衍生NK和T细胞。目前管线均处于临床前期。2022年1月10日,施贵宝(BMY.US)与Century Therapeutics(IPSC.US)宣布就联合开发iPSC衍生的同种异体细胞疗法达成一项研究合作和许可协议。根据协议,Century将获得1.5亿美元现金(1亿美元预付款和5000万美元股权投资),并有可能获得超过30亿美元的开发、监管和商业里程碑付款。公司网址:https://www.centurytx.com/10.Citius Pharmaceuticals(纳斯达克:CTXR)公司简介:CTXR是一家生物制药公司,专注于一流的重症护理产品的开发和商业化,包括辅助癌症护理、肿瘤、干细胞治疗、抗感染药物。Citius通过真皮成纤维细胞创建克隆iPSC主细胞库,再诱导产生i-MSCs,为未来所有的细胞提供了一个同质的、经过验证的细胞来源。Citius的i-MSCs具有优越的效力和高细胞活力,分泌免疫调节蛋白,可减少或预防与呼吸窘迫综合征(ARDS)相关的肺部症状,预计2022年提交IND申请。管线进展如下图:公司网址:https://citiuspharma.com/11.Cynata Therapeutics(ASX:CYP)公司简介:澳大利亚干细胞和再生药物公司,2017年,全球领先的影像和医疗器械巨头富士胶片(FUJIFILM)公司支付了397万澳元收购了Cynata Therapeutics的9%的股权,成为Cynata Therapeutics的大股东之一。根据入股协议,双方将合作开发和商业化CYP-001用于移植物抗宿主病(GvHD),富士胶片(FUJIFILM)有权利优先获得干细胞药物CYP-001全球独家的商业化销售权益。2019年9月日本住友制药会社(Sumitomo Dainippon Pharma Co., Ltd)提出的每股2澳元的收购要约,总价值2.02亿澳元。但10月份放弃收购。Cymerus™是从诱导多能干细胞(iPSCs)生成间充质血管母细胞(MCAs),再用此中间细胞中生成细胞产品。这项技术最初是由威斯康星大学麦迪逊分校(UWM) Igor Slukvin教授(Cynata创始人之一)开发。目前,Cynata正专注于利用Cymerus技术生产基于间充质干细胞(MSC)的产品。目前有三个临床试验:NCT04537351,The MEseNchymal coviD-19 Trial: MSCs in Adults With Respiratory Failure Due to COVID-19 or Another Underlying Cause (MEND),Phase1/2NCT02923375,A Study of CYP-001 for the Treatment of Steroid-Resistant Acute Graft Versus Host Disease,Phase1NCT05165628,Safety, Tolerability and Efficacy of CYP-006TK in Adults With Diabetic Foot Ulcers,Phase1公司网址:https://www.cynata.com12.FUJIFILM Cellular Dynamics (FDCI)公司简介:Cellular Dynamics公司于2004年由iPSC研究领域的先驱James Thomson创立,总部位于威斯康星州的麦迪逊市。2015年3月FUJIFILM以3.07亿美元收购Cellular Dynamics International(CDI),进入基于诱导多能干细胞(iPSC)的细胞发现,开发支持和企业服务领域。通过此次收购,获得了世界领先的iPSC开发生产技术。目前有帕金森、心血管、眼科三个主要管线,均处于临床前阶段。公司网址:https://www.fujifilmcdi.com/about-us/13.Cytovia Therapeutics公司简介:Cytovia在马萨诸塞州纳蒂克有研发实验室,在波多黎各有GMP细胞制造厂房,与Cellectis(前文已述)、CytoImmune、INSERM、国家癌症研究所和加州大学旧金山分校(UCSF)等有广泛合作。Cytovia专注于开发三种类型的iPSCs衍生的NK细胞未编辑的iNK细胞,具有功能改善和持久性的TALEN®基因编辑的iNK细胞,嵌合抗原受体的TALEN®基因编辑的iNK细胞第二种基石技术是一种四价多特异性抗体平台,通过使用专有的Flex-NK™技术靶向NKp46,吸引自然杀伤细胞执行杀伤功能。针对GPC3,CD38和EGFR的管线预计在2022年申请IND。公司网址:https://www.cytoviatx.com/14.Editas Medicine (纳斯达克: EDIT)公司简介:Editas医学总部位于马萨诸塞州的剑桥,是一家领先的基因组编辑公司,专注于将CRISPR/Cas9和CRISPR/Cas12a基因组编辑系统进行商业化开发。基因编辑的CD34+人造血干细胞和祖细胞 (EDIT-301),开展了严重镰状细胞病和输血依赖型地中海贫血(TDT)临床研究。iPSCs-iNk在临床前,iNK和αβ T细胞开始提交IND。公司网址:https://www.editasmedicine.com/15.Exacis Biotherapeutics公司简介:Exacis生物疗法公司通过采用高效的制造工艺来降低治疗成本,同时提高细胞治疗的安全性,使癌症细胞治疗大众化。Exacis通过mRNA编辑iPSC,生产肿瘤学的工程NK和T细胞。三个技术平台:ExaNK™, ExaCAR-NK™ and ExaCAR-T™公司网址:https://www.exacis.com/16. ElevateBio公司简介:ElevateBio 2019年成立于剑桥,A轮融资1.5亿美元,B轮1.7亿美元,C轮5.25亿美元,总计完成超8亿美元融资,基于临床级iPSCs技术,打造下一代免疫细胞治疗和再生医学平台(BaseCamp平台)。HighPassBio是其专注免疫细胞治疗的子公司,重点方向是TCR-T,目前有一项临床开展(NCT03326921,HA-1 T TCR T Cell Immunotherapy for the Treatment of Patients WithRelapsed or Refractory Acute Leukemia After Donor Stem Cell Transplant)。另外一家子公司Allovir,致力于现货型病毒特异性T细胞开发。其中进展最快的是 ALVR105,已进入关键3期临床ALVR106,ALVR109进入临床1期,ALVR107在临床前 公司网址:https://www.elevate.bio/ https://www.allovir.com/17. Fate Therapeutics (纳斯达克:FATE)公司简介:Fate是一家临床阶段的生物制药公司,致力于建立克隆主iPSC细胞系,作为一种可再生细胞源,生产现货型iT细胞,iNK细胞,iCD34细胞,用于免疫治疗。 目前Fate有6个管线进入临床1期,和Janssen(首付款5000万美金,总价值超30亿美金)以及OnoPharma的合作在临床前阶段。 公司网址:https://fatetherapeutics.com/ 18. Heartseed Inc 公司简介:位于日本东京,创始人为庆应义塾大学心脏病学系教授Keiichi FUKUDA, MD/PhD/FACC,致力于通过iPSCs治疗心脏疾病。 2021年6月1日,Heartseed和诺和诺德(NovoNordisk)就Heartseed的心力衰竭治疗药物HS-001的开发、制造和商业化达成了一项全球独家合作和授权协议。该药物是一种细胞疗法,使用诱导多能干细胞(iPSC)来源的纯化心肌细胞。根据协议条款,除日本外,诺和诺德获得HS-001在全球范围内的独家开发、生产和商业化权利。Heartseed将保留HS-001在日本的独家开发权利,诺和诺德有权与Heartseed在日本共同商业化该产品。Heartseed有资格获得总额最高达5.98亿美元的付款,包括5500万美元的预付款和近期里程碑付款。Heartseed还有资格获得日本以外地区年度净销售额的高个位数至低两位数的分级特许权使用费。 公司网址:https://heartseed.jp/ 19. Healios K.K.(2015年东京证券交易所:4593)公司简介:2011年成立于福冈,2013年迁往日本东京,拥有两个干细胞治疗技术平台:体细胞干细胞再生医学和iPSCs衍生NK细胞免疫治疗、再生医学等,治疗实体肿瘤、缺血性中风和急性呼吸窘迫综合征。目前缺血性中风和急性呼吸窘迫综合征在日本获得孤儿药认定,准备申请上市 肿瘤和再生医学在临床前阶段 公司网址:https://www.healios.co.jp/20. Hebecell公司简介:2016年创立,从诱导多能干细胞(iPS细胞)无限生产异基因自然杀伤(NK)细胞。创始人为Shi-Jiang (John) Lu,武汉大学本科毕业,北京协和医学院获得硕士学位,多伦多大学/安大略省癌症研究所获得了肿瘤学和癌症生物学博士学位。一直在利用人类胚胎干细胞(hESC)、诱导多能干细胞(iPSC)及其衍生物进行转化研究和开发新的治疗策略。 加科思药业(1167.HK)于2021年8月31日宣布战略投资Hebecell。交易完成后,加科思药业董事长、CEO王印祥将出任Hebecell公司董事长。 公司网址:https://hebecellcorp.com/21. Hopstem Biotechnology公司简介:霍德生物,位于浙江杭州,由范靖博士创立,以iPSCs技术为核心,进行再生医学开发。天使轮获得1200万融资,后经过5轮,获得数亿元融资。公司的核心技术之一为自主研发的RONA 2.0神经分化技术。霍德生物二期GMP生产基地位于“中国最佳医疗健康产业园区Top10”的杭州医药港,满足美国FDA及欧盟/中国cGMP要求,三套完全独立B+A和C+隔离器的无菌生产条件细胞培养间,同时满足三种不同细胞产品独立生产和检验的需求。 产品管线均在临床前期,其中hNPC-01准备申报IND。 公司网址:http://www.hopstem.com/ 22. Implant Therapeutics公司简介:2020年创立的小型细胞治疗公司,总部马里兰州。主要技术为低免疫原性的 iPSC-MSC,以此为基础生产免疫治疗细胞。 公司网址:https://www.implant-rx.com/ 23. I Peace Inc公司简介:公司总部位于加州Palo Alto,创始人田边刚士2013年在京都大学医学研究院山中伸弥实验室(iPS细胞研究所) 获得博士,在诱导性多能干细胞的早期开发阶段开始参与研究,是全世界第一篇人体诱导性多能干细胞论文的作者,并撰写多篇关于诱导性多能干细胞和体细胞直接重编程为神经元的论文。 I PeaceInc致力于以自动化设备制作细胞(自动化细胞制作工厂),以提高制作效率并降低价格。2020年为AveryTherapeutics的MyCardia产品提供生产。 公司网址:https://ipeace.com/ 24. Mesoblast(纳斯达克MESO,ASX:MSB)公司简介:Mesoblast Ltd是一家创办于2004年的澳大利亚生物技术公司,开发以干细胞为基础的再生治疗产品,拥有间充质细胞技术平台开发临床上方便使用的异体干细胞治疗产品。 3期候选产品组合包括remestemcel-L用于治疗类固醇难治性急性移植物抗宿主病(SR-aGVHD),以及用于covid-19感染引起的中度至重度急性呼吸窘迫综合征(ARDS);REVASCOR®用于晚期慢性心力衰竭;MPC-06-ID用于治疗退行性疾病引起的慢性腰痛。 和诺华、天士力等有相关项目合作。进行的临床研究 公司网址:https://mesoblast.com/ 25. Neurophth Biotechnology Ltd(纽福斯)公司简介:纽福斯成立于2016年,是李斌教授团队于华中科技大学同济医学院附属同济医院自2008年起十几年来对眼科基因治疗技术探索和积累的成果。作为一名眼科医生和视网膜疾病专家,李教授带领团队成功完成了从实验室到临床应用的转化研究,通过一系列国内和国际多中心临床试验证实了腺相关病毒(AAV) 载体技术在眼科基因治疗应用价值。公司地址:https://neurophth.com/ 26. Novo Nordisk stem cell R&D unit公司简介:是诺和诺德独立的研究机构,致力于治疗慢性疾病(帕金森氏病,心力衰竭,慢性肾病,1型糖尿病等)。诺和诺德和BioLamina、Heartseed(前文已述,合作心力衰竭干细胞治疗)、Procyon Technologies(可植入式干细胞治疗1型糖尿病)进行合作。27. Procyon公司简介:Procyon公司2016年建立在亚利桑那,由亚利桑那大学图森医学院创新商业化的初创公司。Procyon细胞封装设备是一个小、平、薄、高度耐用、灵活的可植入式腔,类似于茶包(tea-bag),这是一种皮下植入的免疫隔离设备,用来填充可分泌治疗产品的同种异体细胞,并保护它们免受免疫排斥。该设备利用生物相容性合成膜制成,可减轻异物反应,促进其表面血管结构的形成,使营养物质迅速扩散到设备内部的细胞,并迅速吸收被包裹细胞分泌的胰岛素(或其他治疗分子),同时提供一个屏障,保护这些细胞免受人体免疫系统的攻击,而无需免疫抑制剂药物。Procyon技术是以实际临床应用为驱动力而设计的,它还包括将氧气输送整合进植入式设备中,这样可以在增强细胞生存活力和功能的同时实现更紧密的细胞包装。 公司网址:https://procyon-technologies.com/ 28. REPROCELL公司简介:2003年创立,日本领先的干细胞试剂及科学研究公司,是第一家提供iPSC来源的人类心肌细胞、肝细胞和神经元细胞用于研究应用的公司,并于2013年在日本JASDAQ/Growth股票市场上市。目前开发治疗神经退行性疾病的新型再生医学疗法,目前正在日本进行治疗脊髓小脑共济失调的临床试验。 公司网址:https://www.reprocell.com/ 29. RheinCell Therapeutics GmbH(CDMO)公司简介:RheinCell位于德国Langenfeld(Rheinland)细胞治疗CDMO企业,开发和制造GMP级人诱导多能干细胞,用于下一代细胞治疗。专注于高免疫相容性和低排斥潜力HLA-homozygous iPSC,重点关注现货型同种异体治疗的解决方案。2021年6月,Catalent宣布收购 RheinCell Therapeutics GmbH。 公司网址:https://www.rheincell.de/30. Catalent (细胞基因治疗CDMO龙头,纽交所:CTLT)公司简介:公司2007年被黑石集团收购后成立;2014年开始盈利并重新上市,总部位于美国新泽西州的萨默塞特,在中国上海投资了两家临床供应中心,分别位于外高桥保税区和毗邻张江药谷的浦东唐镇。作为全球生物制品CDMO的领导者,在欧洲和北美拥有最先进的设施,与从全球顶级跨国制药公司到小型和虚拟公司等各种规模的客户都有合作的经验。拥有先进技术,包括GPEx®、SMARTag®和腺相关病毒(AAV)载体,开发生产iPSCs ,CAR-T、TCR-T、NK等各种细胞类型产品。公司网址:https://www.catalent.com/31. xCell Science/RxCell Inc公司简介:RxCell是一家2012年创立,位于加州,专注于诱导多能干细胞(iPSC)治疗应用的生物技术公司,开发了几个治疗级的iPSC系种子库,并正在制造一个大型的主细胞库。结合靶基因敲除和敲入,开发低免疫原性诱导多能细胞系,生成一个通用细胞系,可用于制造多种治疗细胞。现在主要与大小制药公司合作,推动这些治疗项目向前发展。工作主要集中在使用iPSC来源的间充质干细胞作为抗原和抗体传递载体。除了治疗领域外,RxCell还通过其XCell品牌销售细胞、培养基和其他相关试剂,用于学术和药物发现研究。主要重点是将神经和视网膜谱系细胞用于药物发现和研究使用。 公司网址:https://www.rxcellinc.com/32. Sana Biotechnology(纳斯达克:SANA)公司简介:Sana Biotechnology成立于2018年,由原Juno的多为高管创立,是一家临床前生物科技公司,目前没有管线进入临床试验阶段。管线包括体内细胞工程和体外细胞工程两种。体内细胞工程(Fusogen Technology,In Vivo Cell Engineering),通过基因编辑方法,在体内修改细胞,治疗单纯基因治疗无法治愈的疾病。SG295 (CD8/CD19),SG239 (CD8/BCMA),SG242 (CD4/CD19),SG221 (CD4/BCMA),SG328,SG418体外细胞工程(Ex Vivo Cell Engineering)分为三类1)Hypoimmune Technology: Donor-derivedSC291 (CD19),SC255 (BCMA)2)Hypoimmune Technology: Stem cell-derivedSC4513)Stem cell-derived (to migrate toHypoimmune Technology)SC379、SC187公司网址:https://sana.com/ 33. SCG Cell Therapy Pte Ltd/星德汉生物公司简介:总部位于新加坡细胞治疗公司,在德国慕尼黑拥有研发基地,上海建立了星德汉(上海)生物医药有限公司。建立了CAR-T、TCR-T、双抗、疫苗等技术平台。TCR-T进入临床1期。 公司网址:http://www.scgcell.com/34. SCM Lifescience公司简介:2008年成立于韩国仁川的细胞治疗企业,用于干细胞、免疫细胞、组织工程三个管线。2010年开展GVHD1期临床,2016年启动GVHD 2期临床,2018年开始现货型细胞疗法研发,2020年CMN-001在美申请2期临床,治疗肾脏肿瘤。公司网址:http://scmlifescience.co.kr/35. Semma Therapeutics/Vertex公司简介:2014年成立于剑桥,开发全球领先的干细胞衍生的人类胰岛(SC-islets)作为1型糖尿病的潜在治愈性疗法。2019年Vertex将以9.5亿美元现金收购Semma所有已发行股票。Semma将成为Vertex的一个独立运营子公司。 公司管线网址:https://www.vrtx.com/research-development/pipeline/ 36. Shoreline Biosciences公司简介:2020年创立,总部位于加州SanDiego, 是一家生物技术公司,正在开发iPSC 衍生的同种异体现货型NK和巨噬细胞细胞免疫疗法。 Shoreline公司与百济神州达成合作,以联动Shoreline公司的iPSC NK细胞技术与百济神州的临床前研发和临床开发能力,开发并商业化一系列基于NK细胞疗法的产品组合。为此,Shoreline公司将从百济神州处获得4500万美元的现金预付款。吉利德科学(Gilead Sciences)子公司Kite和细胞免疫疗法研发公司Shoreline Biosciences (Shoreline)宣布建立战略合作伙伴关系,以开发针对各种癌症靶标的新型细胞疗法。公司网址:https://shorelinebio.com/ 37. Stemson Therapeutics(干细胞治疗脱发)公司简介:Stemson是一家临床前阶段的细胞治疗公司,成立于2018年,其使命是通过利用诱导多能干细胞(iPSC)的再生能力,来生长新生毛囊,治疗脱发。2020年获得750万美金融资,2021年A轮获得1500万美金融资。公司网址:https://stemsontx.com/ 38. TreeFrog Therapeutics 公司简介:2018年11月在法国成立,该公司开发了名为C-Stem的仿生细胞培养技术-一种高通量细胞封装技术,允许在工业生物反应器中大规模生产和分化干细胞,可以在短时间内大规模生产干细胞,2021年4 月,TreeFrogTherapeutics宣布实现在10L生物反应器中单批生产150亿个多能干细胞(在6.59天内实现了276倍放大),结果被证实具有高度可重复性。2021年9月宣布完成7500万美元的B轮融资,本轮融资由BpifranceLarge Venture领投,Leonard Green & Partners、百时美施贵宝(BMS)和XAnge跟投。本轮融资将用于为临床级制造仿生C-Stem技术,发展和推进在研细胞疗法管线至临床试验阶段,以及资助该公司在美国马萨诸塞州波士顿和日本神户开设技术中心。 公司网址:https://treefrog.fr/39. Vita Therapeutics公司简介:设立在印度的细胞治疗公司,以iPSCs技术为基础,生产各类细胞用于再生医学及免疫治疗,总部设在Telangana,是Cambrian Biopharma药物公司的子公司,A轮获得3200万美金融资。VTA-110,基于iPSCs治疗杜兴型肌营养不良症(Duchennemuscular dystrophy,DMD),2020年获得美国FDA孤儿药认定。并获得马里兰干细胞研究基金(MSCRF) 300K美金商业捐款资助。 公司网址:https://www.vitatx.com/40. Opsis公司简介:Opsis治疗公司成立于2016年,是富士胶片FCDI和David Gamm博士的合资企业,总部设在威斯康星州的麦迪逊市。Opsis治疗公司是一家专注于推进针对眼病的细胞替代治疗管道的公司。利用诱导多能细胞(iPSC)生成、视网膜细胞分化和功能生物材料方面的创新,正在开发治疗干性老年性黄斑变性(AMD)、遗传性黄斑变性(IMDs)和遗传性视网膜疾病(IRDs)患者的细胞疗法。 公司网址:https://opsistx.com/参考来源内容来源于各公司网站,请参考文中提供的公司网址。识别微信二维码,添加生物制品圈小编,符合条件者即可加入生物制品微信群!请注明:姓名+研究方向!版权声明本公众号所有转载文章系出于传递更多信息之目的,且明确注明来源和作者,不希望被转载的媒体或个人可与我们联系(cbplib@163.com),我们将立即进行删除处理。所有文章仅代表作者观点,不代表本站立场。
细胞疗法并购临床2期免疫疗法信使RNA
100 项与 Avery Biomedical Devices, Inc. 相关的药物交易
登录后查看更多信息
100 项与 Avery Biomedical Devices, Inc. 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2026年06月14日管线快照
无数据报导
登录后保持更新
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
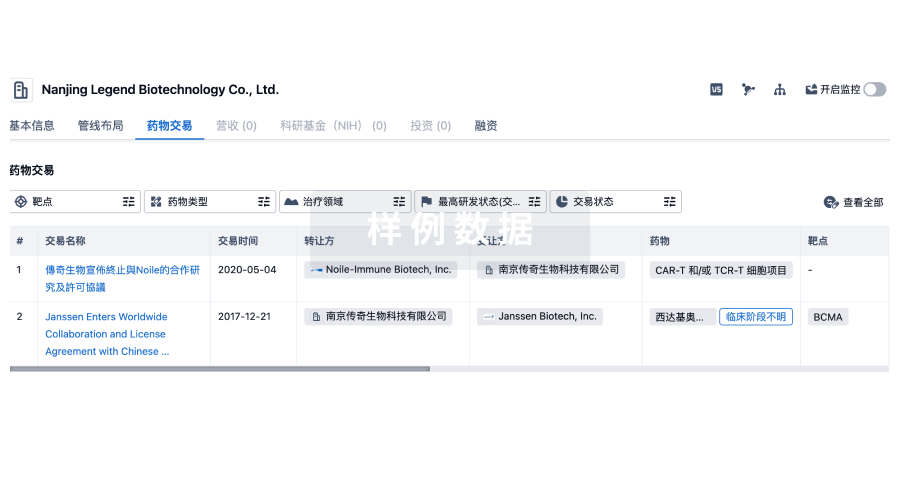
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
生物医药百科问答
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用