预约演示
更新于:2025-05-07
GlaxoSmithKline (GSK) SRL
更新于:2025-05-07
概览
关联
100 项与 GlaxoSmithKline (GSK) SRL 相关的临床结果
登录后查看更多信息
0 项与 GlaxoSmithKline (GSK) SRL 相关的专利(医药)
登录后查看更多信息
4
项与 GlaxoSmithKline (GSK) SRL 相关的新闻(医药)2024-06-30
摘要:病毒样颗粒(VLPs)是具有多种应用的纳米结构,包括治疗、免疫和诊断。随着生物医学工程技术的最新进展,商业化的基于VLP的疫苗正在被广泛用于抗击传染病,同时还有更多疫苗处于临床研究的不同阶段。由于它们在效果、安全性和多样性方面的期望特性,基于VLP的方法在未来几年可能会变得更加常见。然而,在基于VLP的方法能够广泛应用于治疗之前,必须解决一些生产和制造方面的挑战。这篇综述提供了对近期基于VLP疫苗开发情况的见解,重点介绍了它们的特性、表达系统以及作为理想候选者对抗新兴病毒病原体的潜在适用性。最后,详细阐述了基于VLP的疫苗作为可行且高效的免疫剂,诱导对包括SARS-CoV-2在内的病毒性传染病病原体以及基于蛋白质纳米颗粒的疫苗产生免疫的潜力。因此,VLP疫苗可能作为传统疫苗策略的有效替代品,用于抗击新兴的传染病。
1.引言
目前市场上的大多数疫苗主要基于灭活(杀死)或减毒活疫苗的方法。尽管这些传统疫苗已经有效地用于对抗各种传染病,但它们存在一些限制,包括较低的诱导更强免疫反应的潜力和较差的效力。最近传染病的爆发表明了开发强大疫苗以克服这些限制的需求。主要挑战在于开发新的技术方法,增强免疫力,同时不损害安全性、效力和耐受性。最近在DNA、mRNA和重组病毒载体疫苗方面的进展为针对难以靶向的病原体和控制传染病爆发的有效疫苗开发方法。
病毒样颗粒(VLP)技术为开发有效疫苗以抗击严重关注的传染病提供了一个替代平台,并且它与mRNA和病毒载体疫苗并行发展。与其他亚单位疫苗相比,VLPs在免疫原性方面也更强,因为它们在表面以更真实的确认形式呈现重复的抗原表位,免疫系统可以容易地检测到。另一方面,亚单位疫苗由于目标抗原的错误折叠或对免疫系统的呈现不足而具有较差的免疫原性。此外,它们需要佐剂和多次疫苗接种才能引发足够的免疫反应。Fraenkel-Conrat和Williams(1955年)首次通过从纯化的RNA和蛋白质组分重新组装烟草花叶病毒(TMV)颗粒来描述病毒样颗粒这一术语。这些纳米结构诱导强效免疫反应的潜力随后得到了进一步研究。VLP平台可以克服通常与传统疫苗相关的各种问题;具体来说,与减毒活疫苗相关的传染性、向毒株的逆转、突变风险、灭活疫苗的免疫原性降低、不稳定毒性、产量低和配方时间长(图1)。
图1: VLP疫苗与传统疫苗相关风险的比较。
这些生物启发的纳米结构具有来自不同病原体的重复和高度密集的抗原,有助于触发强烈的免疫反应。此外,这些高度免疫原性分子具有病毒蛋白的自组装特性。它们是生物相容的,并在其合成过程中具有结构灵活性的潜力。它们可以通过化学或遗传方式进行修改,具有更高的稳定性、均匀性和功能性,被认为是各种生物医学应用中的有效工具(图2)。它们根据是否存在脂质膜被归类为包膜或非包膜VLP。
图2: 不同类型VLPs的生产及其应用、特性和挑战。(A) 不同的人类致病病毒和寄生虫,(B) 识别形成病原体结构特征并能导致VLPs形成的基因,(C) 将识别的基因整合到质粒等表达载体中,(D) 允许载体在各种表达系统中表达,(E) 形成不同类型的VLPs,如包膜、非包膜和嵌合VLPs。非包膜VLPs可以有两种类型:单蛋白或多蛋白。在多蛋白VLPs中,可能存在单层、多层,并且有些也是马赛克式的。嵌合VLPs可以内部、外部修改,或者可以通过化学偶联进行修改。
然而,与VLPs相关的一些关键挑战包括较低的稳定性、下游处理困难、生产成本高和对环境条件的敏感性。已经在各种表达系统(ESs)中合成了许多不同的VLP,例如细菌、酵母、哺乳动物细胞、昆虫细胞和植物。基于VLP的疫苗可能用于治疗包括HIV、流感、乙型肝炎、戊型肝炎、疟疾、埃博拉病毒、SARS-CoV-2、寨卡病毒、登革热和口蹄疫在内的各种传染病。
几种基于VLP的疫苗已经获得许可,并在市场上商业化,包括Engerix-B R(GlaxoSmithKline)和Recombivax HB R(Merck & Co)针对HBV,Gardasil R(Merck & Co)和Cervarix R(GlaxoSmithKline)针对HPV,Hecolin R(厦门万泰生物技术有限公司)针对HEV,以及MosquirixTM(GlaxoSmithKline Inc.)针对疟疾。这篇综述着重于VLP疫苗开发的基础知识和技术方面的高级技术。此外,还讨论了用于VLP生产的不同ESs以及针对各种传染病,尤其是SARS-CoV-2的潜在疫苗的开发。最后,详细阐述了作为疫苗开发中承载抗原的蛋白质纳米颗粒的支架。
2. 病毒样颗粒的特性
病毒样颗粒(VLPs)通常因其独特特性(图2)而被视为有效的疫苗候选物。它们是强效的免疫刺激分子,展现出高度密集的病毒表面蛋白,以适当的构象和高度重复的方式呈现,引发强烈的T细胞和B细胞获得性免疫反应。大多数VLPs来源于病毒的衣壳或包膜蛋白,尽管核心蛋白也可以使用。VLPs天然生物相容且不具传染性,因为它们缺乏病毒遗传物质,因此无法复制。它们还被认为是比传统减毒活疫苗更安全的(不会恢复到野生型)。此外,它们是高度多功能的分子,大小从20到200纳米不等。这种大小范围是最优的,可以自由地将它们排入淋巴结,并被抗原呈递细胞(APCs),特别是树突细胞(DCs)更容易地摄取,然后通过主要组织相容性复合体(MHC)II类分子进行抗原处理和呈递。它们高度有序,可以自组装成不同的几何对称性,通常以二十面体、螺旋对称性、杆状结构或球形的形式出现,这取决于病毒的来源。
VLPs已在广泛的表达系统(ESs)中合成,包括原核(细菌)和真核(昆虫细胞、哺乳动物细胞系、植物细胞或酵母)。可以通过肽偶联、遗传融合和化学交联等不同方法,在它们的外部或内部表面展示感兴趣的异源表位,从而增加VLPs的功能。VLP技术提供了一个重要优势,因为它是一种更快的疫苗合成方法。在特定菌株测序后12-14周内就可以制备出针对该菌株的新VLP疫苗,而传统疫苗通常需要24-32周的制造过程。这些疫苗不含卵白蛋白,这对容易过敏的个体来说是巨大的缓解,并且比传统疫苗提供更强的疾病保护。
3. 基于结构的病毒样颗粒类型
根据它们的结构复杂性,VLPs可以分为两组:包膜和非包膜VLPs。两组都展示外来抗原(图2)。
3.1. 非包膜病毒样颗粒
这些VLPs通常由目标病原体或病毒蛋白结构的单个或多个自组装组分构成。这些新形成的VLPs中没有宿主细胞膜(脂质包膜)。主要病毒核衣壳蛋白的表达主要负责这些VLPs的形成。非包膜VLPs仍在作为开发针对几种病原体疾病的亚单位疫苗的首选候选物进行研究,因为它们更容易生产和纯化。此外,这些VLPs体积更小,使它们能够轻松穿过组织屏障并排入淋巴结。病毒粒子的主要结构组成部分是由单个病毒编码蛋白形成的,来源于病原体,如杯状病毒、乳头瘤病毒和小病毒,而多蛋白非包膜VLPs则更为复杂,包含多个相互作用的衣壳蛋白。它们展示了几个显著的结构特征,如多个不同衣壳蛋白的复杂同心层。它们比仅由一个主要衣壳蛋白(CP)构成的VLPs更难制造,来源于传染性腺胃炎病毒、脊髓灰质炎病毒和逆转录病毒。
3.2. 包膜病毒样颗粒
与非包膜VLPs相比,包膜VLPs(eVLPs)的组成要复杂得多。这些纳米结构由来自宿主细胞的细胞膜称为包膜,外表面有病毒蛋白存在。它们的脂质双层中嵌入了一个或多个糖蛋白刺突,作为产生中和抗体的靶抗原。这些eVLPs显示出更高的灵活性,因为它们针对相同或异源病毒的抗原表位。例如,已经开发了包含SIV的Gag蛋白和HIV的Env蛋白的eVLPs。尽管这可能会影响下游应用,因为存在宿主蛋白。eVLPs已被用于开发针对病毒疾病,如汉坦病毒、丙型肝炎病毒(HCV)、流感A和逆转录病毒的疫苗。这些大型eVLPs的尺寸大于100纳米,因此它们可能会在注射部位聚集,并且不会到达淋巴结,这限制了它们的应用。相比之下,携带mRNA疫苗的脂质纳米颗粒尺寸范围在100纳米或更小,并在将疫苗高效传递到目标部位方面显示出高效性。
3.3. 嵌合病毒样颗粒
嵌合病毒样颗粒(cVLPs)被认为是开发提供更广泛、更强大、更全面保护以应对新兴传染病的有效工具。这些复杂的多蛋白宏观结构包含不同病毒的表位。cVLPs可以通过构建编码相关病毒蛋白和外来肽或蛋白的重组DNA分子来创建。这些基因工程cVLPs在其表面展示大量重复序列,可以通过化学偶联或遗传融合从其他病毒加载外源抗原。不同的嵌合疫苗已经进行了临床试验,包括针对乙型肝炎的基于VLP的疫苗(M2–HBcAg)、抗疟疾疫苗(MalariVax)、抗流感A、抗HIV和减少吸烟者血液中尼古丁水平的尼古丁-Qb VLP疫苗。这些嵌合颗粒有利,因为它们显著增加了对外国抗原的免疫反应和抗体滴度。cVLPs给药后,它们诱导强烈的细胞溶解性T淋巴细胞免疫反应。这些疫苗还针对非传染性疾病,如高血压、阿尔茨海默症、尼古丁成瘾、过敏和糖尿病。cVLPs的上游和下游处理产量通常较低,其体内稳定性也相当不确定。
4. 病毒样颗粒作为免疫原
已经证明,与亚单位疫苗相比,VLPs具有潜在的高免疫原性和抗原性。这些颗粒的效力有可能显著诱导细胞和体液免疫(图3)。在对VLPs的反应中,各种成熟标志物如CD40、CD80和CD86在树突细胞表面表达,这些标志物负责激活树突细胞。在第一步中,树突细胞通过与树突细胞表面存在的特定模式结合,称为模式识别受体(PRRs),即Toll样受体(TLR2),来激活VLPs。随后,VLPs在树突细胞的细胞质中发生内化,并通过MHC I类和II类分子分别呈递给细胞毒性T细胞和辅助T细胞。VLPs不仅可以刺激B细胞介导抗体反应,还可以刺激CD4+和CD8+细胞增殖。一些研究表明,外源性抗原也可以通过一种称为交叉渗透的过程到达MHC I类途径。
图3: VLPs诱导先天和适应性免疫反应(A)体液免疫;(B)细胞介导的免疫),(1)基于VLP的抗原通过树突细胞等抗原呈递细胞(APCs)增强吸收和呈递,这些细胞向T细胞通报潜在风险,(2)VLP有效运输到淋巴结,这是适应性免疫反应的关键场所,(3)改善B细胞、T细胞和APCs之间的细胞通信,以及(4)基于VLP的抗原有效交联并激活B细胞受体,这些受体在抗原暴露后发展成记忆细胞和长期及短期存活的浆细胞。
此外,B细胞的激活可以通过Toll样受体(TLR)信号或同源相互作用诱导Th细胞的扩增和分化,这些相互作用控制细胞因子的产生。为了提高VLPs的效力,还可以将不同的分子如Toll样受体配体、生物活性介质或其他细胞受体附加到VLPs上。
5. 基于病毒样颗粒的商业批准疫苗
第一种重组VLP是从病毒衣壳蛋白合成的,来自乙型肝炎病毒(HBV)的两个基因(HBsAg和HBcAg)和烟草花叶病毒(TMV)。第一种通过重组DNA方法生产的商业VLP疫苗在1986年获得了美国食品药品监督管理局(FDA)的批准。这些是在酵母中生产的HBsAg疫苗,被命名为Recombivax HB R。后来,在2006年,第二种基于VLP的疫苗Gardasil R针对人类乳头瘤病毒(HPV)的疫苗获得了FDA的许可。此后,几种针对HPV和HBV的VLP疫苗已经获得批准,其中一些在临床和临床前试验中显示出效力。研究表明,至少有110种VLP已经从35个不同家族的病毒中生产出来。针对不同病毒的几种VLP疫苗,包括诺如病毒、HIV、埃博拉病毒、SARS-CoV-2病毒、呼吸道合胞病毒(RSV)、流感病毒仍在不同的临床试验阶段。然而,只有少数VLP疫苗进入市场,显示出它们的商业可行性,其中大多数针对非包膜病毒有效。
6. 特定于基于病毒样颗粒疫苗平台的挑战
目前,VLPs与传统疫苗一样有效,还有更安全的额外好处。然而,在成功开发基于VLP的疫苗方面需要解决几个障碍。主要挑战是确定与下游处理有关的问题,以生产临床可行的VLPs,以便及时管理并具有经济可行性。
6.1. 包膜病毒样颗粒的稳定性
VLPs通常被认为比亚单位疫苗更稳定。然而,当环境条件发生变化时,特别是在下游处理过程中,VLPs可能变得非常不稳定,因为它们缺乏病毒的遗传物质。尽管市场上已经有多种VLP疫苗可用,但一些候选疫苗的稳定性存在问题。通常,eVLPs通常比非包膜VLPs更容易受到外部环境条件的影响。温度变化、剪切应力、溶解氧、流体动力学、搅拌速率和化学处理等条件的变化都可能影响颗粒的完整性和稳定性。此外,这种结构破坏显著降低了eVLPs的免疫原性。它还干扰细胞生长和代谢蛋白的产生,这影响了VLP的生产。这一直是将它们作为疫苗制造中活病毒替代品使用的关键障碍之一。然而,已经进行了几种修改以增加它们的热稳定性。一种典型的程序是插入稳定突变。Marsian等人进行的一项研究表明,当在3型脊髓灰质炎病毒VLP的衣壳蛋白中诱导稳定突变时,它比野生型VLP更稳定。它修改了衣壳前体和病毒蛋白酶,而不影响其抗原表位和结构确认。Schumacher等人的另一项研究通过添加C末端连接体-六组氨酸肽增加了嵌合体(HBcAg-VLP)的稳定性。
6.2. 高生产成本
一些基于VLP的疫苗在结构上更为复杂,因此生产成本更高。与eVLPs相关的众多杂质带来了巨大的挑战。在下游处理(DSP)过程中,会像包膜杆状病毒颗粒的情况一样,共同纯化各种杂质,如宿主细胞碎片(HCD)、宿主细胞蛋白(HCP)和宿主细胞DNA(HCD)。如果这些污染物在DSP期间没有正确去除,它们可能会在疫苗中引起不良副作用。大规模生产和纯化VLPs需要不同的工艺,如密度梯度或甚至色谱法,以制造最终配方产品。这些复杂工艺成本高昂且耗时。这也导致了在工业规模生产上的困难,并需要进行多项质量控制措施,因为不同的下游加工步骤可能会降低VLPs的质量。需要更健壮和更好的分析方法来确保产品的质量和数量,以便促进它们在临床和制药领域的应用。在VLPs的上游和下游加工过程中,已经实施了不同的策略,如克隆筛选、高通量筛选、生物反应器工程、材料/基质筛选、过滤、流穿或尺寸排除色谱和抛光,以实现可扩展和成本效益高的工业制造。
6.3. 组装困难
将表位序列遗传融合到VLPs中有时可能具有挑战性,因为VLPs可能会失去自我组装属性或导致颗粒错误折叠。将抗原遗传融合到病毒的衣壳蛋白通常会阻碍抗原组装或VLP,使技术变得繁琐。因此,需要耗时的规划和优化,以单独测试每个单一抗原。
7. 病毒样颗粒生产的表达宿主系统
已经使用了不同的表达系统(ESs)来制造VLP疫苗,包括植物、哺乳动物、昆虫、酵母和细菌(图4)。
图4:不同表达系统在病毒样颗粒开发中的优缺点。
7.1 细菌
大多数细菌系统都集中在大肠杆菌(Escherichia coli)的工业菌株和表达载体上,这些已经得到了深入研究。细菌细胞培养作为VLP开发的平台进行了研究,具有成本和可扩展性方面的优势。使用细菌系统生产VLP的其他有利特点包括(a)易于操作,(b)高水平表达,(c)快速生长速率,(d)遗传稳定性,以及(e)表达的简单性。
Huo等人使用pCold表达载体在大肠杆菌菌株(BL21)中表达和纯化诺如病毒(NoV)VLPs,并展示了在大肠杆菌中组装的VLPs与在Sf9细胞中组装的VLPs具有相似的结合模式。类似地,在最近的一项研究中,表达全长CP的2型猪圆环病毒和猪小病毒的VP2蛋白在大肠杆菌中表达,它们自我组装成VLPs。研究表明,大肠杆菌中CP和VP2的表达可以用于大规模开发VLP疫苗。Yazdani等人研究了利用葡萄扇叶病毒(GFLV)的VLPs作为呈现HPV L2表位的潜在载体的可能性。抗原因子决定簇序列被遗传地结合到GFLV衣壳蛋白的“αB-αB”域C中,然后在大肠杆菌和毕赤酵母中过度表达。在大肠杆菌中观察到最高的表达产量。对于戊型肝炎病毒(HEV),ORF2蛋白区域368-606 aa从大肠杆菌不溶性分数中纯化体外组装成VLPs。这种HEV VLP在临床试验中对症状性HEV有100%的效果,并已在中国作为疫苗批准商业使用。在另一项研究中,从重组IHHNV的CP重建的传染性皮下和血胞坏死病毒(IHHNV)VLPs在大肠杆菌中显示出出色的物理稳定性。
由于几个因素,细菌系统并不总是VLP开发的最佳策略,包括(a)免疫原性差,(b)无法发展具有类似哺乳动物的翻译后修饰(PTMs)的重组蛋白,(c)蛋白质溶解度问题,(d)无法形成正确的二硫键,以及(e)在重组蛋白的制备中存在细菌内毒素/或脂多糖。
Brito和Singh强调了各种类型疫苗的可接受内毒素水平。有各种方法可以在纯化步骤中去除内毒素,如固定化琼脂糖、表面活性剂、活性炭、超滤和阴离子交换色谱。然而,使用这些技术通常会导致产量显著降低、成本增加或目标蛋白的生物活性缺乏。因此,开发了ClearColiTM,这是一种基因工程大肠杆菌菌株,具有基因修饰的LPS,不会在人类中引起内毒素反应。
7.2 酵母
真核表达系统是原核系统的有力替代品,特别是当涉及到解决疫苗生产中细菌内毒素问题时。通过在酵母细胞中重组表达蛋白,可以更具成本效益地生产VLPs。酵母中的基因操作相对简单,转化细胞可以生长到极高的密度,直到诱导重组蛋白的表达。这允许商业规模的发酵罐大规模生产VLPs。一些哺乳动物病毒的结构基因在酵母中表达可以形成VLPs。
美国食品药品监督管理局(FDA)已经批准了一些基于酵母衍生的VLP疫苗,包括针对人类乳头瘤病毒(HPV)的Gardasil®和Gardasil9®,以及针对恶性疟原虫(P. falciparum)的MosquirixTM。一些研究已经使用酵母作为表达系统(ES)来研究VLP的生产。Wetzel等人建立了一个健壮且成本效益高的酵母模型,用于生产嵌合VLPs。选择了鸭乙型肝炎病毒膜整合小表面蛋白(dS)作为VLP的支架,以及安全且工业上应用的Hansenula polymorpha酵母作为异源表达宿主。从感染动物的四种病毒中获得了8种不同的高分子量抗原,并将它们与蛋白dS遗传链接,然后鉴定并纯化了重组分离物。在所有情况下,融合蛋白都高度表达,并且与蛋白dS共产后,可以生成包含两种蛋白的嵌合VLP。基于酵母的生产系统允许生产成本低廉的产品,且不仅限于小规模基础研究。在另一项研究中,Alireza等人通过将HPV-16 L2的交叉中和表位插入到HPV-16 L1基因中,首次在P. Pastoris系统中生产了嵌合蛋白L1/L2 VLPs。随后,将HPV-16的嵌合L1/L2引入质粒(pPICZA)并在P. pastoris中表达。L1-HPV-16 Ab和L2-HPV-16 Ab检测的ELISA结果表明,与商业测试套件的灵敏度相当的阳性反应。同样,在P. pastoris ES中,Gupta等人生产了一种针对HEV的重组VLP,包括蛋白ORF2的112-608 aa区域。结果表明,对于112-608 aa VLP的开发,P. pastoris ES似乎是比杆状病毒(Bv)ES更优越且更安全的选择。在病毒感染期间,像脊髓灰质炎病毒这样的肠道病毒会产生与成熟病毒粒子在抗原上无法区分的空衣壳。通过异源系统(如酵母)的帮助,这种衣壳的重组合成作为VLP疫苗候选物具有巨大的潜力。Sherry等人通过共表达病毒蛋白酶3CD和结构前体蛋白P1,在P. pastoris中展示了VLP的生产。构建酵母ES比细菌ES更具挑战性,特别是Pichia和Hansenula菌株。此外,VLP产量低于大肠杆菌。酵母ES的另一个缺点是它在蛋白质翻译后修饰(PTMs),特别是糖基化方面与哺乳动物ES的相似性不足。它们的糖型主要是高甘露糖型,这对大多数制药糖蛋白(GPs)来说是不受欢迎的。
7.3. 昆虫细胞
基于杆状病毒的昆虫细胞系蛋白表达已成为开发复杂基于蛋白质的生物制剂的有效工具,用途多样,从多蛋白复合体到用于治疗用途的蛋白质开发,如VLPs。Bv在能够容纳大量异源DNA并将其忠实地传递到目标宿主细胞的能力方面,与其他广泛使用的病毒载体不同寻常。Gopal和Schneemann详细描述了生产重组杆状病毒(rBVs)、筛选昆虫细胞系中VLP表达以及纯化VLPs的程序。
Bv-昆虫细胞系统是一个两步程序,首先将昆虫细胞培养到所需的细胞浓度,然后感染rBVs以表达蛋白质。使用Bv表达载体系统(EVSs)的重组蛋白和VLP生产是快速、多功能的,并提供大量的产量。该系统可以产生异常高的蛋白质产量,结合复杂的真核蛋白质PTMs,这对于某些VLPs的正确自组装和释放可能至关重要。
MultiBac是一个创新的Bv EVS,它由一个已经被设计用于特定目的的病毒基因组组成。最近,一组科学家描述了基于MultiBac的VLP工厂的开发,该工厂依赖于流感病毒的CP(M1)及其在生成具有流感病毒血凝素(HA)功能修饰的流感衍生VLPs中的利用,这些修饰预计将调节VLP衍生的免疫反应。
昆虫细胞-Bv EVSs已被证明与流感病毒疫苗生产的常规鸡蛋和基于细胞的方法一样有效,具有高产量和短生产时间的额外特点。此外,rBVs构建变得更快、更容易、更灵活,允许在相同的表达载体中融合不同类型和/或亚型的流感病毒基因。Sequeira等人成功创建了一个强大的High Five基于昆虫的细胞平台,该平台结合了稳定的表达和Bv介导的表达,以产生多价流感VLPs。通过用两种不同的H3亚型HA蛋白(称为HA2和HA1)感染High-Five细胞系,并用表达M1和三种额外的H3亚型HA蛋白(称为HA5、HA4和HA3)的Bv,这种方法的模块化能力得到了证明,以产生五价VLPs(H3)。
扎伊尔埃博拉病毒血清型(ZEBOV)是所有血清型中最有毒的,并且具有最高的死亡率。通过共表达3种病毒结构蛋白,核衣壳蛋白(NP)、基质结构蛋白(VP40)和糖蛋白(GP),已经在昆虫和哺乳动物细胞系中实现了ZEBOV-VLPs的开发。Pastor等人报告了一种在昆虫细胞系中生成ZEBOV-VLPs的技术,基本上包括使用高感染度(MOI)的bac-GP和bac-VP40,并限制NP表达,要么通过防止感染,要么通过降低bac-NP MOI,是开发VLP的最适当方法。
昆虫细胞ES的主要限制是(a)被包膜杆状病毒污染的蛋白质,(b)难以扩大规模,以及(c)比哺乳动物细胞更简单的N-糖基化。
7.4. 哺乳动物细胞
二十多年来,各种哺乳动物细胞系,如小鼠骨髓瘤(Sp2/0、NS0)、中国仓鼠卵巢(CHO)、小鼠C127、婴儿仓鼠肾(BHK21)、HT-1080和HEK293,由于它们能够正确折叠、组装和翻译后修饰蛋白质,已被建立为商业化蛋白质治疗剂的可能来源,用于医疗目的。基于哺乳动物细胞培养的系统提供了许多好处,包括在开发过程中的一致性和灵活性。它还有助于糖蛋白以与病毒宿主相似的脂质膜组成被恢复。据报道,将病毒基因稳定转染到哺乳动物细胞系中,包括293或Vero细胞,可以产生VLP。
多项研究报告了从哺乳动物细胞培养中高效生产VLPs。Hsin等人利用Huh7细胞作为表达系统,开发了一种MERS VLP系统,用于了解病毒感染和形态形成。在另一项研究中,通过瞬时转染在Vero、293 T或CHO-K1细胞系中表达了编码神经氨酸酶(NA)、血凝素(HA)和基质M1蛋白的流感VLPs。在BALB/c小鼠的临床前研究显示,当肌肉注射给药时,流感VLPs在低剂量下具有显著的免疫原性,诱导针对NA和HA的功能抗体。Wu等人设计了一种可扩展的方法,用于有效生产在哺乳动物细胞系中表达的各种亚型的流感VLPs。该研究表明,这些哺乳动物流感VLPs在粒径、结构、宿主因子组成和病毒抗原糖基化方面与原始病毒非常相似。同样,在另一项工作中,开发并利用了一种可诱导的人类胚胎肾HEK-293细胞系,表达NA和HA,以在用编码HIV-1 Gag的质粒进行瞬时转染后生成VLPs。为在哺乳动物细胞悬浮培养中通过瞬时基因表达合成HIV-1 Gag VLPs制定的协议表明,细胞培养上清中存在的大部分Gag-GFP完全组装成与HIV-1未成熟颗粒一致的预测形态和大小的VLPs。
基于哺乳动物细胞的表达系统需要大规模生产设施,如发酵生物反应器,这些设施建造成本过高。因此,生产成本高昂是细胞基础的哺乳动物ES的一个具有挑战性的部分。其他限制包括长时间的表达时间、产量低以及易受哺乳动物病原体感染的脆弱性。
7.5. 植物
“分子农业”是用来描述利用植物或植物细胞生产重组蛋白或其他生物药物的术语,这些药物用于作为化妆品、生物制药、治疗剂、疫苗和其他试剂。植物为VLP疫苗的生产提供了一个诱人的替代系统,因为它们有潜力以最小的成本生成大量的重组蛋白,它们具有正确的PTM和蛋白质折叠组装的真核生产机械,并且它们引入意外人类病原体的风险低。植物不需要安装昂贵的发酵设施来生产生物质,也不需要建立重复的设施来扩大生产规模。与微生物发酵不同,植物有能力进行N-糖基化作为糖蛋白PTM。在植物细胞中表达的工程VLP形成人类或动物病毒衣壳蛋白包括人类诺如病毒CP、HPV L1蛋白、乙型肝炎核心和表面抗原、HIV Gag多蛋白和流感病毒的HA蛋白。
多项研究已经广泛讨论了在各种植物ESs中构建VLPs的概念、在植物宿主中有效生长VLPs以及在植物中自组装多种病毒结构蛋白。Santi等人利用TMV衍生的瞬时ES,在烟草植物叶片中生产了正确组装的重组诺如病毒VLPs。Young等人生成了可追踪的基于血凝素的VLPs,使他们能够在植物中检查颗粒组装和VLPs在哺乳动物免疫系统内的相互作用。van Zyl等人通过农杆菌介导的瞬时表达,研究了在烟草植物中生产蓝舌病病毒VLPs,这是一个低成本系统。同样,Veerapen等人在烟草植物中瞬时表达了口蹄疫病毒的VLPs。Diamos和Mason使用改良的双生病毒载体在基于植物的系统中生产了诺如病毒VLPs。最近,Roales-Mendoza等人提出了一种针对SARS-CoV-2的基于植物的VLP疫苗开发的视角,SARS-CoV-2是导致COVID-19大流行的罪魁祸首。
用于VLP平台生产的植物并不完全可接受,因为与哺乳动物ESs相比,VLP的生产水平相对较低,以及植物特有的N-糖基化糖蛋白。然而,最近创造的新型植物ESs,以及植物糖工程的进步,都已经解决了这些挑战。植物糖工程的最新进展允许人类样的糖修饰,并优化所需的糖结构,以提高重组药物糖蛋白的功能和安全性。
8. 针对新发传染病的病毒样颗粒疫苗的开发
近年来,已经生产并使用了多种VLP疫苗来对抗不同的病毒和寄生虫感染(表1)。下面讨论了针对寨卡病毒、基孔肯雅病毒、流感病毒和人类乳头瘤病毒的VLP疫苗的开发。
表1: 针对不同病毒和寄生虫的VLP疫苗
8.1. 寨卡病毒
寨卡病毒(ZIKV)是一种小包膜蚊媒神经性正链RNA病毒,属于黄病毒科。ZIKV感染与急性产前异常有关,例如感染母亲所生的新生儿小头症,以及成人中的格林-巴利综合征(GBS)。目前,没有治愈方法或商业疫苗可用于有效治疗,因此开发针对ZIKV的疫苗非常重要。
ZIKV具有一个RNA基因组,只有一个开放阅读框,并形成一个单一的多蛋白。这种蛋白链被细胞和病毒蛋白酶切割成7种非结构蛋白(NS5、NS4B、NS4A、NS3、NS2B、NS2A和NS1)和3种结构蛋白(C、prM和E)。E蛋白参与病毒粒子的结合及其融合。E蛋白具有三个域结构,像包膜域-I也称为ED-I,然后是ED-II和ED-III。体液反应主要针对属于ED-II的融合环。针对融合环表位的抗体增强了通过Fcγ受体摄取DENV。因此,这种蛋白可能是疫苗开发的一个可能靶点。
已经开发了许多针对ZIKV的VLPs。Garg等人比较了小鼠中不同的ZIKV VLPs疫苗,并表明CprME-VLPs(衣壳前膜-包膜)比prME-VLPs(前膜-包膜)效果更好。由于需要共表达蛋白酶NS2B-3,无法使用CprME-生成细胞系。为了克服这个障碍,生成了一个双顺反子载体,利用IRES序列产生NS2B-3和CprME-VLPs。Alvim等人展示了HEK293细胞持续表达ZIKV-VLPs。简而言之,稳定产生寨卡VLPs的细胞系是开发疫苗的理想选择。
8.2. 基孔肯雅病毒
这是一种由携带蚊虫传播的CHIKV(基孔肯雅病毒)引起的严重和周期性传染病。症状包括高烧、皮疹等。目前没有可用的疫苗,但有许多VLP疫苗候选物处于不同的开发阶段。通过选择性表达CHIKV包膜和衣壳蛋白产生的自组装VLPs可以提供对这种病毒的多种菌株的保护。
Arévalo等人报告说,当使用无佐剂CHIKV VLPs时,成年小鼠对CHIKV感染实现了100%的保护。同样,Metz等人比较了在昆虫中由重组杆状病毒产生的三种不同疫苗,它们分别产生sE1、sE2 CHIKV和CHIKV(VLPs)。与VLP免疫的小鼠相比,E1和E2免疫的小鼠有一半存活,显示出不完全的保护。Akahata和Nabel发现,可变CHIKV结构蛋白表达产生的VLPs,模仿具有复制能力的α病毒。这种疫苗引起针对这种病毒的多种菌株的大量中和抗体的诱导,并提供完全保护。
与CHIKV-OPY-1菌株的VLPs相比,CHIKV 37997菌株的VLPs更容易产生,尽管它们具有高度的氨基酸序列相似性。了解CHIKV VLP生产的机制将有助于其他疫苗的开发,也扩大了它们对其他病原体的应用范围。此外,VLP生产受到E2的第234个氨基酸在酸敏感区域的影响。酸敏感区域的突变和pH变化也提高了VLPs的产量。
8.3. 流感病毒
流感病毒(Iv)感染是导致人类慢性呼吸道症状的主要原因,导致严重的公共卫生结果,包括流行性和季节性感染,甚至导致不可预测的大流行爆发。Iv是一种包膜的、分段的、负义RNA病毒,属于Orthomyxoviridae科。迄今为止,已经鉴定出四种类型的Iv,根据它们的核心蛋白存在情况分类:A、B、C和D。三种Iv类型,A、B和C对人类细胞具有致病性并引起严重感染,其中A型和B型是最普遍流行的类型。流感病毒的主要表面糖蛋白是神经氨酸酶(NA)和血凝素(HA)。
HA是参与感染的关键抗原,它附着在细胞膜表面的唾液酸残基上,促进Iv进入宿主细胞。与HA相比,NA在病毒表面上出现的频率较低,通常观察到的NA:HA比例为1:4。它的酶功能在切割唾液酸中至关重要,从而促进病毒从感染的宿主细胞表面释放。NA激活还使流感病毒能够通过涉及唾液酸切割的机制成功穿透粘液。
Kim等人研究了来自2009年H1N1流感病毒大流行的含NA(N1 VLP)的VLP的交叉保护效率和免疫原性,并将其与灭活分裂流感疫苗进行了比较。用N1 VLPs免疫的小鼠能够诱导病毒特异性抗体(Ab)反应以及交叉反应的NA抑制活性,而灭活分裂疫苗则诱导了株特异性的血凝抑制试验。用N1 VLPs免疫的小鼠导致对抗原多样化的Iv产生交叉保护免疫力,通过体重变化、肺病毒滴度、先天白细胞浸润、细胞因子和Ab分泌细胞、生发中心B细胞的测量。此外,在未处理的小鼠中,N1 VLPs的免疫血清提供了交叉保护。N1 VLPs诱导的免疫力在缺乏Fc受体γ链的小鼠中既没有受损也没有减少。这些发现表明,代表NA的VLPs,以及当前的流感疫苗,可能进一步改进并开发成为交叉保护疫苗的重要候选物。
最近,Kirsteina等人研究并检查了一系列广泛保护原型Iv疫苗的效力和免疫原性,重点关注流感三联矩阵蛋白2离子通道(3M2e)和并入噬菌体AP205 VLPs的三叉抗原。仅含3M2e抗原的VLPs在小鼠中刺激了对标准同源以及异源病毒挑战的保护。将流感病毒的两种保守抗原结合到一个单独的VLP中,可在小鼠中实现对高剂量同源流感H1N1感染的完全保护。
8.4. 人类乳头瘤病毒
人类乳头瘤病毒(HPVs)是Papillomaviridae家族的成员,是微小的、圆形的、双链DNA病毒,导致宫颈癌,这是继乳腺癌之后世界上第二致命的女性疾病。目前,有两种基于VLP的疫苗可供商业使用,用于抑制由HPV引起的疣和治疗宫颈癌。这些包括GlaxoSmithKline(GSK)制药公司的Cervarix和Merck制药公司的Gardasil。两种疫苗均由HPV 16和18的免疫原性主要衣壳蛋白(L1)VLPs组成,Gardasil还含有6和11 VLPs。
使用L1蛋白作为VLP疫苗的抗原的缺点是它在不同的HPV血清型中并不保守。相反,次要衣壳蛋白(L2)在所有HPV血清型中高度保守,长期以来一直被认为是开发具有广泛保护的HPV疫苗的主要潜在靶点抗原。与衣壳蛋白(L1)相比,衣壳蛋白(L2)不能用于VLP生产,因此免疫原性较低。L2蛋白免疫原性低的原因是由于其相对于L1的轻微表达,以及L2主要埋藏在衣壳表面下,在那里它在体外与细胞表面接触,或在体内与基底膜接触并诱导构象转变。
已经应用了几种方法来提高和增强L2肽的低免疫原性,例如由多种HPV类型的L2表位组成的串联融合肽,结合免疫激活的TLR激动剂,或在颗粒免疫刺激平台(如TMV、MS2或PP7噬菌体、腺相关病毒、干酪乳杆菌或细菌硫氧还蛋白的VLP)上重复表面阵列的L2表位。
另一种提高L2低免疫原性的有前景的策略是通过由HPV L1-L2嵌合蛋白组装的VLPs在表面展示L2表位。通过将高度保守的B细胞表位RG1的HPV16 L2遗传插入到HPV16 L1蛋白的表面环(DE)中实现,导致其在VLP表面上多价(360倍)免疫原性展示,而HPV16 L1框架的构象中和抗原决定因素大多保留。用这种嵌合RG1-VLP免疫引起了高型特异性HPV16滴度和广泛的异源粘膜和远亲皮HPVs的交叉中和。
Pouyanfard等人描述了基于单一肽的热稳定硫氧还蛋白疫苗开发,能够携带多达11种不同类型HPV的L2多肽。L2多肽的抗原在免疫应答的稳健性和保护方面表现出色。为了进一步提高和增强免疫原性,将硫氧还蛋白的多肽抗原L2与七聚体化域融合。在疫苗的最终设计中,实现了对所有14种致癌型HPV,以及较低风险的HPV类型(6和11),以及一系列皮肤HPV的保护反应。
9. 病毒样颗粒对抗严重急性呼吸综合征冠状病毒2
去年,全球有超过200万人的死亡和2亿多人受到影响,200多个国家受到严重影响,严重急性呼吸综合征冠状病毒2(SARS-CoV-2)已成为全球性的麻烦。由SARS-CoV-2引起的COVID-2全球大流行影响了数十亿人,并成为2009年H1N1大流行之后的第一个重大全球灾难。目前,一些商业上可用的抗病毒药物,包括Remdesivir®(吉利德科学公司)、Paxlovid®(辉瑞公司)、Molnupiravir®(默克公司)已获得FDA和WHO的批准,用于治疗感染SARS-CoV-2的患者。由于针对病毒的目标药物疗法在开发、成本和分发方面的限制,研究人员正将重点和努力放在开发疫苗上,以长期对抗COVID-19。针对SARS-CoV-2的VLPs可以帮助抗击广泛流行的大流行。VLPs可以作为治疗剂和针对这种病毒疾病的可行疫苗,此外还可以作为可行的诊断工具。SARS-CoV-2有一个缓慢的作用机制,因为病毒进入宿主后,一个人需要5到15天才会出现症状,因为病毒通过内体途径进入宿主细胞,并不擅长逃避免疫系统,而针对其他冠状病毒(CoVs)株的疫苗,这些株引起SARS和中东呼吸综合征(MERS),已被证明在动物模型中对SARS-CoV-2有效和活跃。像其他CoVs一样,SARS-CoV-2由四种在不同病毒血清型中结构保守的蛋白质组成:刺突(S)、核蛋白(N)、包膜(E)和膜(M)蛋白。上述蛋白质及其相关交互模式对VLPs的生产和组装至关重要。M、N和E对SARS-CoV-2的VLPs生产至关重要(图5)。S蛋白就是这样一种在不同冠状病毒株中常见的蛋白质,可能作为潜在的疫苗开发目标。通过在昆虫宿主细胞ES中展示S蛋白,已经生产了针对MERS-CoV的VLPs。
图5:提议的SARS-CoV2病毒样颗粒疫苗的生产系统和作用机制。编码SARS-CoV2的结构蛋白(S、N、M和E)的质粒可以转染到适当的哺乳动物细胞系中。然后收集、纯化组装的VLPs,并将其施用于人类。VLPs的施用刺激了先天和适应性免疫反应。如果原始的SARS-CoV2将来进入人体,记忆B细胞激活并释放针对它的抗体。同样,激活的CD8+ T细胞识别并杀死病毒感染的细胞。
在开发基于SARS-CoV-2 VLP的疫苗之前,必须首先考虑以前针对先前和密切相关的菌株的VLPs的工作。以前已经证明VLPs在通过rBV ES生产时,通过在小鼠中引起特定的体液和细胞免疫防御来有效对抗SARS-CoV。Ho等人报道了SARS-CoV的VLPs的合成。这些VLPs是通过rBV的共感染在昆虫ES中组装的。通过使用蛋白质冠层形成制备的VLPs在CoV感染的禽类模型中显示出有效的疫苗接种,并导致显著的免疫反应。同样,含有SARS-CoV的4种结构病毒蛋白(S、N、E和M)的质粒被转移到Vero E6细胞中以生产VLPs。这些VLPs可以作为研究SARS-CoV与宿主细胞病理机制的有效工具。最近,一种表达SARS-CoV-2 VLPs的COVID-19 mRNA疫苗在小鼠模型中显示了诱导强大的抗病毒免疫反应。
目前,NVX-CoV2372是唯一商业上可用的针对SARS-CoV-2的基于VLP的疫苗。这种疫苗是通过在rBV中展示SARS-CoV-2 S蛋白生产的,然后通过感染蛾细胞表达系统进行大规模生产。这种疫苗在不同国家的不同试验中显示出不同程度的有效性。目前,还有五个团队分别致力于针对COVID-19的基于VLP的疫苗,临床试验正在进行中,正在努力开发可行的基于VLP的疫苗。已经开发出生物计量VLPs,可以通过在RT-PCR过程中作为阳性对照来帮助准确诊断SARS-CoV-2。Ghorbani等人使用免疫信息学分析评估了各种SARS-CoV-2刺突衍生表位的免疫原性质,这些表位已被报道可诱导特定的免疫反应。最后,建议一组筛选出的表位,这些表位可以在不同植物物种中使用分子农业方法合成针对SARS-CoV-2的基于VLP的疫苗。
不同的哺乳动物ESs被用于合成SARS-CoV-2 VLPs。结果表明,来自Vero E6细胞系的SARS-CoV-2 VLPs比来自HEK-293T细胞的更耐用、更集成。最近,有报道称一种基于植物ESs的针对SARS-CoV-2的VLP疫苗。针对SARS-CoV-2的VLP疫苗(CoVLP)的临床I期试验结果表明,该疫苗在个体中显著发展了IL-4和IFN-γ免疫反应。CoVLP疫苗是通过在植物ESs(N. benthamiana)中短暂表达SARS-CoV-2的S蛋白生产的。三聚体S GPs展示在自组装VLPs的表面,模仿SARS-CoV-2的大小和形状。同样,针对SARS-CoV-2的组合小刺突VLP疫苗在小鼠中也显示出高水平的免疫,在单剂量后。疫苗引发了中和Abs的发展,并保护K18-hACE2小鼠免受COVID-19的侵害,类似于患有COVID-19的患者。
以衣壳VLP为中心的SARS-CoV-2疫苗(ABNCoV2)在小鼠模型中显示出有效的SARS-CoV-2中和。ABNCoV2疫苗是通过在昆虫(Drosophila)细胞中展示SARS-CoV-2 S蛋白的受体结合域生产的。研究人员还在致力于开发一种可以通过鼻腔输送到目标组织的基于DNA的疫苗,这将在个体中产生SARS-CoV-2 VLPs,从而在个体中产生强烈的免疫反应。
10. 蛋白质纳米颗粒在疫苗开发中的应用
蛋白质纳米颗粒(NPs)有助于开发针对免疫逃逸病原体的疫苗,如HIV、流感、疟疾,并通过单独作用或作为靶向药物传递的载体,帮助对抗新兴的毒株。利用NPs辅助的蛋白质分子工程进行抗原表达,用于疫苗开发以产生免疫反应,是治疗和药物开发中日益受欢迎和迅速发展的领域。基于蛋白质的病毒样颗粒(VLPs)作为天然NPs,因设计简便、自组装和高稳定性而在疫苗开发中得到广泛应用。有三种不同的方法用于将抗原附加到NPs上进行展示,(1)标签偶联,(2)化学偶联,以及(3)遗传融合。这些技术使平台能够装饰各种抗原,导致尺寸增加和展示效果增强。
大多数抗原通常不会自行组装成NPs,如在流感亚单位疫苗中使用的那些。在这种情况下,可以通过将这些抗原附加到多聚蛋白平台上来实现自组装。有许多天然存在的多聚蛋白,如蝶呤合酶(LS)、铁蛋白、二氢硫辛酸乙酰转移酶(E2p)、非结构蛋白10(nsp10)、包裹素和热休克蛋白(HSPs),这些已被开发用于设计靶向治疗的平台。使用NPs展示病毒糖蛋白在开发针对特定抗原的抗体方面是有效的。展示抗原的蛋白质NPs在产生针对特定抗原的免疫反应方面非常有效,可以单独使用或与疫苗结合使用。
在可用于展示抗原的广泛平台中,铁蛋白已成为一个主要的蛋白质复合物,可以与NPs结合用于疫苗开发。由于其高pH和热稳定性,它允许与表面分子容易结合。近年来,铁蛋白在临床前研究中作为针对多种病毒传染病的可行疫苗平台进行了研究,包括HIV、H1N1、HCV、HBV、HFMD、Epstein–Barr、轮状病毒和由冠状病毒引起的呼吸道疾病。
Kelly等人比较了体内展示流感HA的铁蛋白NPs与可溶性蛋白(HA)的免疫原性。与可溶性蛋白(HA)相比,HA-铁蛋白NPs显示出更高的免疫原性和对病毒挑战的更大保护作用。
最近,一种基于结构的自组装蛋白质NP免疫原的设计,针对SARS-CoV-2产生了保护性和强大的抗体反应,在小鼠的体内研究中展示了先天免疫的发展。同样,基于铁蛋白NPs的SARS-CoV-2疫苗在小鼠中诱导了有效的抗体反应,据报道这种反应至少在免疫后持续了7个月,有助于充分发展免疫。
尽管蛋白质NPs可以作为疫苗开发的可行候选物,但由于其有机性质,NPs的体内应用通常受到几个挑战的限制,包括细胞毒性、炎症反应和不足以递送到目标部位。“蛋白质冠层”形成这个术语被用来总结体内条件下蛋白质与NPs之间不利相互作用。附加到NPs表面的蛋白质影响其生物行为并改变其功能,有时导致功能增加或丧失。蛋白质冠层形成是一个复杂的过程,涉及两个相互作用实体之间的复杂动态和动力学。有些病例报告了在与NP表面相互作用后发生了结构变化,明显改变了NPs的原始功能。因此,为了克服这些挑战,了解蛋白质构象变化和展开过程对于加速NPs的生物医学应用至关重要。
11. 结论
在过去二十年中,一些病原体对商业化抗生素的耐药性急剧增加,主要是由于无监管的非处方销售、药物的滥用和过度使用以及遗传适应。药物开发是一个严格的任务,需要充足的时间、资金和重复试验,才能使一个可行的产品商业化,以帮助对抗众多新兴病原体。由于耗时的限制,特别是患者获得适当药物的机会受到阻碍,而病原体的耐药性却在持续增加。因此,预防措施对于解决传染病的紧急情况具有重要意义。在努力抗击这类疾病的过程中,VLPs在医疗行业中崭露头角,应用范围从疫苗开发、药物传递系统到分子诊断等多个领域。基于VLP的方法为疫苗开发提供了一种替代现有传统方法的选择。目前,一些针对危险病原体的基于VLP的疫苗已经商业化,如ZIKV、HCV、HBV和HPV。与此同时,正在努力生产和设计针对包括SARS-CoV-2在内的各种新兴毒株的有效VLP疫苗。然而,在基于VLP的治疗能够与传统药物疗法在成本和效果方面竞争之前,仍需要解决一些主要障碍。尽管如此,基于VLP的医疗策略未来可能会与药物制剂一起广泛使用,以帮助对抗众多传染病。
识别微信二维码,添加生物制品圈小编,符合条件者即可加入
生物制品微信群!
请注明:姓名+研究方向!
版
权
声
明
本公众号所有转载文章系出于传递更多信息之目的,且明确注明来源和作者,不希望被转载的媒体或个人可与我们联系(cbplib@163.com),我们将立即进行删除处理。所有文章仅代表作者观点,不代表本站立场。
疫苗信使RNA
2023-08-27
摘要:大多数疫苗都是通过肌肉接种。皮肤含有大量的抗原提呈细胞,被认为是疫苗接种更具免疫原性的部位。皮内接种已被批准用于提高流感疫苗的效力和节省流感疫苗剂量。为了应对最近暴发的猴痘疫情,皮内接种也被批准通过延长有限的猴痘疫苗剂量,以免疫更多有风险的人。疫苗佐剂的加入有望进一步提高皮内接种效力,节省更多疫苗剂量。然而,与肌肉接种相比,皮内接种与更显著的局部反应有关。因此,适合于皮内接种的佐剂应具有良好的局部安全性,而不会引起明显的局部反应。本文介绍了目前已批准用于人类疫苗中的佐剂及其皮内给药的相对反应原性、具有良好局部安全性可用于皮内接种的新型化学和物理佐剂,同时对开发物理佐剂的合理性、物理佐剂的类型以及物理佐剂用于皮内接种的独特优势也进行了介绍。1.皮肤结构皮肤覆盖全身,具有重要的生理功能,如防止热量和水分流失,防止环境病原体感染。皮肤可分为3层:角质层(SC)、表皮层和真皮层。SC层位于皮肤的最外层,由角质细胞组成,是终末分化的角质细胞,并且具有高度的亲脂性。SC层承担皮肤的主要屏障作用。表皮位于SC层之下,主要由角质细胞(90%)组成,中间散布着朗格汉斯细胞(LC,3-5%),作为免疫哨兵)。表皮层还含有黑色素细胞,可根据其密度和黑色素含量赋予皮肤特殊的颜色。表皮层下面是真皮层,它比SC层和表皮层包含更多的解剖结构。真皮层中含有汗腺和毛囊,以及血液和淋巴管网和神经纤维。真皮层还包含多种类型的细胞,包括成纤维细胞和先天性免疫细胞,如树突状细胞(DC)、巨噬细胞、肥大细胞和嗜酸性粒细胞。有趣的是,皮肤还含有大量的T淋巴细胞,其中大部分是表皮和真皮层中皮肤驻留记忆T细胞。皮肤先天性免疫细胞是抵御病原体感染的第一道防线,也有助于诱导病原体或疫苗特异的适应性免疫应答。图1显示了带有常驻天然免疫细胞的简化皮肤模型。图1.皮肤结构和固有免疫细胞的图解:皮肤包含3个主要层:角质层、表皮和真皮。表皮含有朗格汉斯细胞,真皮含有树突状细胞、巨噬细胞、肥大细胞和嗜酸性粒细胞为主的天然免疫细胞。2.皮内疫苗接种及其好处由于肌肉注射的便利性,仅需要简单培训即能操作,因此大多数疫苗都是通过肌肉接种进行免疫。过去20-30年的大量研究发现,将疫苗接种途径从肌肉途径改为皮内途径可以诱导更有效的免疫反应。免疫原性更强的皮内接种在很大程度上可以归因于皮肤而不是肌肉中存在丰富的抗原提呈细胞,如表皮LC和真皮DC。抗原提呈细胞(主要是真皮DC)负责真皮内疫苗抗原在主要组织相容性复合体(MHC)I或II分子上的摄取和递呈,然后迁移到引流淋巴结,引发抗原特异性T和B细胞应答。皮内接种疫苗还可以激活局部先天性免疫细胞来分泌细胞因子和趋化因子,并招募循环中的先天免疫细胞,从而促进疫苗引起的局部反应。皮内接种有望能提高疫苗功效并节省疫苗剂量。最近的一项综述研究发现,在老年人中,皮内递送流感疫苗通常比肌内或皮下注射更具免疫原性。另外,在年轻人中,皮内接种低剂量流感疫苗(6或9µg)与肌肉注射全剂量疫苗(15µg)引起的免疫反应相当。除了流感疫苗外,皮内接种也能有效增强其他疫苗诱导的免疫反应,例如狂犬病疫苗和乙型肝炎疫苗。由于近期猴痘疫情暴发以及猴痘疫苗短缺,美国食品和药物管理局(FDA)授权皮内接种猴痘疫苗,以扩大疫苗剂量,让更多高危人群获得免疫接种。3.皮内接种的局限性皮内接种虽然提高了疫苗的免疫原性,但提高的程度相对较弱。皮内接种只能节省40-60%的流感疫苗剂量。在老年人中,皮内接种仅能将H1N1株的血清转换率从40%提高到52.3%,将H3N2株的血清转换率从49.1%提高到61.3%,将B型株的血清转换率从56.4%提高到61.3%。另一项研究也发现了类似的结果,即通过皮内接种流感疫苗,老年人的血清转换率仅略有增加。疫苗佐剂的加入有望进一步提高疫苗效力并节省更多的疫苗剂量。然而,添加佐剂的皮内疫苗面临着诱发严重局部反应的风险。研究一致发现,与肌肉接种疫苗相比,不含佐剂的皮内接种疫苗会诱发更频繁、更严重的局部不良反应。皮肤中存在的各种类型的先天性免疫细胞(如LCs、DCs、巨噬细胞)可以解释这种情况。这些细胞有助于增加抗原摄取和更有效的适应性免疫,同时通过合成和释放细胞因子和趋化因子以及招募外周中性粒细胞、单核细胞和嗜酸性粒细胞而引起局部不良反应。因此,用于皮内接种的佐剂需要比用于肌肉注射的佐剂具有更好的局部安全性。本综述侧重于介绍用于预防传染病疫苗的皮内接种佐剂,尽管这种途径对其他疾病(例如过敏和恶性肿瘤)的疫苗接种也很有吸引力。本综述还重点介绍佐剂如何增强皮内蛋白和亚单位疫苗诱导的免疫反应。此外,具有佐剂效应的疫苗/佐剂制剂(如聚合物纳米颗粒、脂质体)(至少有一部分是由二者联合引起的),不包括在本综述中。4.已获批准的佐剂及其皮内反应原性疫苗佐剂在开发新型疫苗和改良疫苗方面发挥着关键作用,因此在21世纪受到越来越多的关注。美国国家过敏与传染病研究所(NIAID)于2008年启动了"佐剂开发计划",以支持新型佐剂的筛选、鉴定和临床前/临床开发。随着对这一领域的兴趣和投资不断增加,在过去二十年中,有五种佐剂被批准用于人类,而在过去八十年中,只有两种佐剂被批准用于人类(表1)。有趣的是,所有这些佐剂都被批准用于肌肉接种。如果没有皮内反应原性数据,则根据其肌肉注射后的局部不良反应来预测其皮内给药后诱发局部不良反应的可能。局部不良反应可能是皮肤红斑、肿胀、变色或组织学变化,如组织坏死或细胞死亡,或局部炎症反应,或细胞因子和趋化因子释放及免疫细胞浸润等。表1.目前批准的佐剂及其皮内反应原性l明矾佐剂明矾佐剂是一种铝盐类佐剂。临床上明矾佐剂主要以氢氧化铝为基础,由于纤维状纳米颗粒(图2A)或磷酸铝形成板状纳米颗粒的聚集而形成广泛的微米结构(0.5-10微米)。明矾佐剂是全球使用最广泛的佐剂,它已被用于多种儿童疫苗,如百白破(破伤风、白喉和百日咳)、乙型肝炎、乙型流感嗜血杆菌疫苗,明矾佐剂主要增强Th2细胞的抗体反应,但诱导细胞介导免疫反应的能力较弱。将明矾佐剂注射到肌肉组织中会引起明显的组织应激和细胞死亡。腹膜内注射明矾佐剂后也能观察到类似的现象。肌内注射明矾佐剂会诱导细胞因子/趋化因子的大量释放并快速募集中性粒细胞、单核细胞和嗜酸性粒细胞。研究发现,在小鼠皮肤中皮内注射Imject(一种实验性明矾佐剂)可诱导高密度炎症细胞浸润,并持续至少四天。由于明矾佐剂会诱发严重的组织应激和炎症,因此在皮内注射后很有可能诱发严重的局部反应。lAS04佐剂AS04佐剂是通过将MPL(3-O-去酰基-4'-单磷酰脂质A)吸附在氢氧化铝佐剂上制备的组合佐剂(图2B)。MPL是一种Toll样受体(TLR)4激动剂。MPL/TLR4结合激活骨髓分化初级反应88(MyD88)和下游信号通路,导致核因子-κB(NFκB)和激活蛋白(AP)-1激活以及促炎细胞因子基因表达。AS04被批准用于增强人乳头瘤病毒疫苗的效力。研究发现,与明矾佐剂疫苗相比,AS04佐剂疫苗可在局部注射部位诱导更高水平的细胞因子和趋化因子,并在引流淋巴结中募集更多数量的单核细胞和DC。研究发现,与不含佐剂的疫苗相比,含有AS04佐剂的疫苗会引起更频繁的局部反应,例如疼痛、发红和肿胀。局部不良反应可在几天内消失。研究发现AS04佐剂可诱导更平衡的辅助性T细胞1(Th1)/Th2免疫反应。我们之前的研究发现。皮内注射MPL/明矾佐剂会引起皮肤真皮组织红斑和肿胀,并伴有大量炎症细胞浸润。这些数据表明AS04佐剂在皮内递送后诱发严重局部反应的风险很高。lAS01佐剂AS01是一种含有MPL和QS21的脂质体制剂,QS21是从肥皂树(Quillajasaponaria)中纯化的皂苷成分(图2C)。AS01佐剂已被批准用于提高疟疾RTS,S疫苗的功效。研究发现AS01佐剂可诱导Th1为主的免疫反应,而对Th2反应的诱导较弱。研究发现肌肉注射AS01佐剂被快速清除,3h时AS01水平显著降低,24h后几乎检测不到AS01水平。AS01诱导细胞因子的短暂表达,该表达在24h内达到峰值,然后迅速消退。AS01还诱导中性粒细胞和单核细胞迅速聚集到注射佐剂的肌肉中。考虑到AS01含有MPL,预计局部细胞因子和趋化因子的刺激至少部分是由TLR4和下游MyD88信号通路的刺激引起的。研究发现MPL和QS21能协同激活驻留淋巴结的自然杀伤细胞(NK)和CD8+T细胞并促进早期干扰素(IFN)γ产生,该细胞通过白细胞介素(IL)-18分泌通过荚膜下的巨噬细胞控制白细胞介素(IL)-18分泌,并得到IL-12支持。MPL和QS21的协同作用以及对局部炎症的强烈诱导表明,AS01佐剂在皮内给药后诱发显著局部反应的风险很高。这种作用由巨噬细胞通过白细胞介素(IL)-18的分泌来控制,并得到IL-12的支持。MPL和QS21的协同作用以及对局部炎症的强烈诱导表明,AS01佐剂在皮内给药后诱发显著局部反应的风险很高。lMF59和AS03佐剂MF59和AS03是基于角鲨烯水包油型纳米乳液佐剂。角鲨烯是一种天然三萜油,作为动物和人类胆固醇合成过程中的一种生化中间体。角鲨烯的可生物降解性使其成为佐剂配方中的安全成分。MF59和AS03分别由Novartis和GlaxoSmithKline(GSK)开发,化学成分略有不同(图2D)。MF59的主要油相成分是角鲨烯,而AS03的油相中含有角鲨烯和α-生育酚(维生素E)。α-生育酚还具有可生物降解性和免疫调节功能。关于表面活性剂,MF59含有两种表面活性剂(Tween80和Span85),而AS03仅含有聚山梨醇酯80(Tween80)。MF59最初被批准用于提高欧洲老年人接种季节性流感疫苗功效。MF59最近在美国被批准用于增强老年人季节性流感疫苗(FluadQuadrivalent)的效力以及大流行前H5N1疫苗(Audenz)的效力。AS03最初被批准用于增强欧洲2009年甲型H1N1流感大流行期间的2009年甲型H1N1大流行流感疫苗(Pandemrix)效力,后来被批准用于增强美国大流行前H5N1疫苗的效力。研究发现MF59和AS03佐剂可诱导偏向Th2的免疫反应,而对Th1反应的诱导较弱。肌内注射MF59佐剂会诱导组织应激和细胞死亡。此外,MF59的清除不依赖于抗原,在肌肉中的清除半衰期为42h。研究发现,尽管时间招募模式相同,但发现肌内注射MF59比肌内注射明矾更能有力地募集中性粒细胞、单核细胞和嗜酸性粒细胞。与肌内注射后的明矾佐剂相比,肌肉注射MF59佐剂可诱导多种细胞因子和趋化因子释放模式。皮内注射MF59样AddaVax佐剂可强烈诱导中性粒细胞、单核细胞和嗜酸性粒细胞,注射后4d,中性粒细胞和单核细胞水平显著升高。皮内注射AddaVax佐剂还可诱导各种细胞因子和趋化因子的延长表达至少4d。这些数据表明MF59佐剂在皮内递送后诱发严重局部反应的风险很高。研究发现,肌内注射AS03佐剂可持续诱导至少7d的细胞因子表达(例如C-C基序趋化因子配体2(CCL2)、IL-1β)。AS03比明矾佐剂诱导更强的细胞因子表达。AS03佐剂中的α-生育酚可通过激活巨噬细胞和单核细胞显著增加细胞因子的表达,AS03佐剂可诱导脂质代谢快速且短暂的下调,导致细胞内脂质水平升高,诱导内质网(ER)应激并激活未折叠蛋白反应途径。此外,消耗髓系细胞中ER应激激酶传感器肌醇需要的跨膜激酶内切核糖核酸酶-1α(IRE1α)会减少AS03佐剂诱导的细胞因子表达和高亲和力抗体的诱导。局部炎性细胞因子和趋化因子以及ER应激的强烈诱导表明AS03佐剂也可能在皮内递送后诱导显著的局部反应。lMatrix-M佐剂Matrix-M佐剂由两种独特的40nm颗粒配方制成,来自皂苷提取物的两种组分(组分A和C)(图2E),并已被纳入最近批准的基于蛋白质的Novavax新冠疫苗(NVX-CoV2373),用于新型冠状病毒肺炎大流行期间的紧急使用。每种纳米颗粒制剂均由皂苷A或C部分以及胆固醇和磷脂制成。Matrix-M佐剂被发现可诱导偏向Th1的免疫反应。研究发现,含有Matrix-M佐剂的新冠疫苗与无效对照剂相比更容易引起局部反应(第一剂:57.6% vs 17.9%;第二剂:79.6% vs 16.4%)。大多数局部不良反应为轻度或中度,并在3天内消失。Matrix-M佐剂可刺激猪单核细胞和淋巴细胞表达IL-1β和C-X-C基序趋化因子配体8(CXCL8),但不表达肿瘤坏死因子α(TNFα)或IL-6。猪体内肌肉注射Matrix-M佐剂可诱发以中性粒细胞、单核细胞、淋巴细胞浸润为特征的急性炎症。局部不良反应还包括大多数猪的出血和部分猪的坏死。Matrix-M佐剂组比生理盐水对照组出现发红的频率更高。与对照相比,Matrix-M佐剂显着增加了384个基因表达并降低了162个基因表达。al,2012).在上调最多的基因中,23个中有10个与干扰素调节基因(IRGs)有关。尽管迄今为止还没有研究探索Matrix-M在促进皮内疫苗接种方面的有效性和安全性,但一项研究发现在猪模型中皮内给药脂质体中的QS-2(另一种基于皂素的佐剂)会剂量依赖性地诱发局部不良反应,如红斑、肿块和结痂。人类受试者肌肉注射免疫后出现较频繁的局部不良反应,至少一些猪出现强烈的局部炎症反应、出血和坏死,以及与之密切相关的QS-21佐剂皮内递送后诱发局部不良反应的剂量依赖性,都表明Matrix-M佐剂皮内递送后诱发严重局部反应的风险很高。lCpG 1018CpG 1018是一个未甲基化的含CpG基序的寡核苷酸。CpG 1018可激活TLR9和MyD88信号通路,从而激活各种转录因子,如NFκB、AP-1和干扰素调节因子(IRF),并控制细胞因子的表达,如TNFα、IL-1β、I型干扰素和IL-12。CpG1018刺激偏向Th1的抗体反应和细胞介导的免疫反应。CpG 1018已被纳入最近批准的乙肝疫苗。添加CpG 1018佐剂的乙肝疫苗被发现比添加明胶佐剂的乙肝疫苗诱导更有效的免疫反应。与物种特异性CpG寡核苷酸不同,CpG 1018在啮齿动物、非人类灵长类动物和人类中具有广泛的响应,从而在临床前和临床研究中不需要改变CpG序列。研究发现皮内注射小鼠特异性CpG只会诱发轻微的局部反应和低水平的免疫细胞招募。CpG 1018预计具有较低的皮内反应原性,有望安全地促进皮内疫苗接种。 图2.目前批准的颗粒状佐剂:A.氢氧化铝佐剂。B.AS04佐剂是由MPL吸附在氢氧化铝助剂上制备的。C.用脂质体包裹MPL和QS21制备AS01佐剂。D.角鲨烯纳米乳剂MF59和AS03佐剂。E.由两种皂素提取物(组分A和C)制成的两种纳米颗粒制剂组成的Matrix-M佐剂总之,大多数已批准的佐剂在皮内给药后诱发严重局部反应的风险很高。有趣的是,预测皮内反应原性较高的化学佐剂本质上都是自然界中微粒(表1),它们会强烈激活局部先天性免疫系统并诱发明显的局部反应。CpG1018可能是个例外,因为它能诱导轻微炎症,而不会引起明显的局部反应(图3A)。图3.皮内接种安全佐剂的化学结构A.CpG 1018。DNA骨架进行硫代修饰以提高佐剂的稳定性。B.具有六个14碳侧链的合成GLA。C.咪喹莫特。D.Poly(I:C)。E.cGAMP。F.PCPP聚合物。5.用于安全皮内接种的其他化学佐剂由于担心已获批准的佐剂具有较高的局部反应原性,人们还探索了其他化学佐剂的安全性和有效性,以促进皮内疫苗接种。其中几种佐剂已进入临床试验阶段。由于疫苗佐剂在皮内给药后有可能诱发明显的局部反应,因此本综述不包括未报告局部安全性数据的佐剂。lGLA-AF佐剂几种已获批准的佐剂,如AS01和AS04,都含有MPL作为提升疫苗接种效果的主要成分。虽然MPL还没有被批准作为一种独立的佐剂,但我们在小鼠模型中发现MPL皮内注射引起了轻微的炎症,没有明显的局部不良反应。Carter等人评价了一种合成的TLR4激动剂(GLA-AF)(图3B)以促进雪貂和人类皮内接种H5N1疫苗,并发现GLA-AF对于皮内接种H5N1疫苗在雪貂中诱导单针保护和在人类中诱导血清保护至关重要。此外,在人类临床研究中,加入GAL-AF的H5N1疫苗没有在豚鼠体内引起体温上升或体重下降,并没有引起一过性红斑和其他明显的不良反应。这项研究表明GLA-AF和其他可能的TLR4激动剂用于皮内接种的有效性和安全性。l外用咪喹莫特佐剂FDA批准外用咪喹莫特乳膏(5%阿尔达拉)用于治疗各种疾病,如光化性角化病、浅表性基底细胞癌和外生殖器疣。咪喹莫特是一种TLR7激动剂(图3C)并激活TLR7-MyD88信号通路,导致NFκB和IRF7的激活以及促炎细胞因子和I型干扰素基因的表达。有趣的是,外用咪喹莫特乳膏的载体也可能有助于观察到的疗效,因为在没有咪喹莫特的情况下,诱导激活炎症小体、角质细胞死亡和IL-1释放。在一项双盲、随机对照的临床试验中,在皮内接种流感疫苗前局部应用咪喹莫特乳膏可以获得更高的疫苗病毒株血清转换率和更好的非疫苗病毒株血清转换率。最常见的局部不良反应是1级红肿,尽管与其他组没有统计学差异,但局部应用咪喹莫特组红肿发生率更高。在炎症性肠病(IBD)患者中,外用咪喹莫特乳膏后皮下注射乙型肝炎疫苗的血清保护率明显高于外用水乳膏后肌肉注射疫苗的血清保护率。虽然局部咪喹莫特组引起更频繁的瘙痒和肿胀,但各组之间的局部不良反应无明显差异。这些结果表明,外用咪喹莫特乳膏可显著提高皮内疫苗的疗效,且总体上具有良好的局部安全性。lPoly(I:C)佐剂聚肌胞苷酸(Poly(I:C))是双链RNA(dsRNA,图3D)的合成类似物,可根据位置激活两种不同的信号通路。内质多聚体(I:C)激活TLR3并诱导IL-12和I型IFN产生,而胞质多聚体(I:C)可激活视黄酸诱导基因I(RIG-I)和黑色素瘤分化相关蛋白5(MDA-5)并增强I型干扰素的产生。Poly(I:C)还用于皮内或透皮疫苗接种,以诱导针对各种类型传染病的有效免疫反应。Weldon等人发现,当在小鼠模型中将Poly(I:C)佐剂亚单位流感疫苗涂覆在微针表面时,可引发与单独的亚单位疫苗类似的免疫反应和保护佐剂效果不明显,可能与该研究中使用的Poly(I:C)剂量相对较低有关(1µg)。在另一项研究中,Bardel等人发现25µg Poly(I:C)可以显著增强皮内HIV-1gp140或HSV-2gD糖蛋白诱导的全身和粘膜抗体反应以及在HSV-2攻击研究中对小鼠模型的保护作用,而不会明显增强小鼠模型中对皮肤反应原性的保护作用。这项研究表明,皮内接种添加Poly(I:C)佐剂的HIV-1gp140或HSV-2gD糖蛋白疫苗具有良好的安全性和效力,可诱导有效的粘膜免疫。Du等人发现,将白喉毒素(DT)和Poly(I:C)封装到脂质体中,然后以空心微针为基础进行皮内给药,与单独以空心微针为基础进行皮内单独接种DT相比,可显著增强DT特异性IgG2a抗体反应。有趣的是,该研究中使用了相对较低的poly(I:C)剂量(0.31µg),但并未观察到总IgG或IgG1抗体反应的显著增强。最近,在一项1期临床研究中测试了用聚-L-赖氨酸稳定的poly(I:C)与光化学内化(PCI)相结合增强人乳头瘤病毒多肽疫苗接种的效果,光化学内化是一种通过光诱导过程促进内吞抗原的胞体释放,从而促进诱导细胞介导的免疫反应的技术。尽管没有研究基于poly(I:C)佐剂在增强人乳头瘤病毒多肽疫苗接种方面的效力,但研究发现,当光敏剂剂量低于一定阈值时,整体对研究对象是安全的。上述研究表明,poly(I:C)佐剂用于加强皮内接种具有良好的安全性。lcGAMP佐剂2′3′-环GMP-AMP(cGAMP,图3E)是细胞内干扰素基因刺激物(STING)通路的刺激物,可导致NFκB和IRF3激活以及促炎细胞因子和I型IFN基因表达。cGAMP及其类似物,如cdAMP和cdGMP,已被探究为诱导Th1免疫应答对抗传染性疾病和癌症的有效佐剂。研究发现,在小鼠和猪模型中,皮内注射cGAMP与H5N1流感疫苗或2009年大流行H1N1疫苗可显著增强疫苗诱导的免疫反应,而不会引起明显的局部不良反应。此外,在小鼠模型中,cGAMP被发现是一种比CpG或MPL更有效的佐剂,可增强皮内对H5N1免疫。最近,Hernandez-Franco等人探索了cdAMP和一种以植物为基础的阳离子α-D-葡聚糖纳米粒(Nano-11)增强小鼠和猪皮内OVA免疫的安全性和有效性,发现cdAMP佐剂的皮内OVA免疫在注射后24小时诱发一过性变色,14天后消失,而Nano-11佐剂的皮内OVA免疫则不会诱发明显的不良反应。这些研究支持STING激动剂作为安全的佐剂来促进皮内疫苗接种。lPCPP佐剂微针是将疫苗递送至皮肤的一种极具吸引力的技术。疫苗可以涂布在金属或不可溶解聚合物微针的表面,或封装在可溶解聚合物微针结构中。考虑到微针通常会诱导与肌肉注射给药相当的免疫反应,人们探索了安全的佐剂来促进基于微针的皮内疫苗接种。Andrianov等人发现,PCPP(图3F)是一种合成的、水溶性、可生物降解的聚磷嗪聚合物,在药物输送和组织工程中有着广泛的应用,是一种安全的佐剂,可促进猪的基于微针的皮内乙型肝炎疫苗接种。此外,PCPP可作为一种关键的微针疫苗制造材料,以减少对表面活性剂的依赖。在这项研究中,只观察到轻微的皮肤红痕,没有发现严重的不良反应。除PCPP外,研究还发现一种新型聚磷腈聚合物PCEP可安全促进仔猪皮内接种H1N1流感疫苗,尽管对H3N2病毒缺乏异种亚型保护。总之,皮内反应原性低的化学佐剂通常是水溶性的或制成水性制剂,以便于从局部给药部位快速清除。这些佐剂大多基于病原体相关分子模式(PAMPs)或其合成类似物,可激活模式识别受体(PRRs),如TLR3、TLR4、TLR7、TLR9和STING,从而在不过度激活先天性免疫系统的情况下发挥佐剂作用。虽然PCPP不是PAMP,但研究发现PCPP可通过TLR3、TLR4和TLR9直接激活免疫细胞。这些研究表明,某些PRR激动剂作为皮内接种佐剂是安全的。6.安全用于皮内疫苗接种的物理佐剂除了注射化学佐剂,人们还探索了在皮内注射疫苗前在皮肤表面短暂施加物理能量,以促进皮内疫苗接种。l开发物理佐剂的合理性由于缺乏对佐剂作用原理的了解,过去的佐剂开发主要依赖于经验或试错方法,尤其是那些非PAMP型佐剂,如明矾、MF59和QS21。这些佐剂没有特定的细胞受体来介导其佐剂效应,因此不会直接激活先天性免疫细胞,如直流电和巨噬细胞。相反,这些佐剂更有可能刺激组织应激和细胞死亡,从而释放内源性危险信号或损伤相关分子模式(DAMPs)来介导其佐剂效应。20世纪90年代,Matzinger和她的同事提出了"危险理论",证明免疫系统是由受压或受损细胞释放的危险信号激活的。"危险理论"解释了在没有外来病原体入侵的情况下诱导适应性免疫,如移植排斥、某些化疗诱导的全身性抗肿瘤免疫激活。在过去的二三十年中,人们发现了各种类型的DAMPs,其中包括尿酸和ATP等小分子化学物质以及dsDNA等大分子物质,它们能够根据"危险理论"调节免疫系统的功能。这些物质在生理条件下被封闭起来,无法被免疫系统识别,但在组织应激或细胞死亡时会释放出来,激活免疫系统。研究发现,明矾佐剂可刺激尿酸释放,从而激活NOD-、LRR-和含吡啶结构域蛋白3(NLRP3)炎性小体和Caspase-1激活,部分介导其促炎症反应。明矾佐剂还能刺激宿主DNA的释放,从而诱导IgG1抗体的产生和IgE类的转换,并延长DC和CD4+T细胞的相互作用。MF59佐剂可刺激ATP释放,从而增强疫苗接种的免疫反应。这些进展表明,刺激组织应激或细胞死亡的物理模式可能会诱导内源性危险信号的释放,从而促进疫苗接种。l物理佐剂类型研究最多的两种疫苗佐剂物理能量是激光和射频。图4总结了各种类型的激光佐剂和射频佐剂。图4.不同类型的物理佐剂。简要说明不同类型的激光佐剂(全表面、非消融部分、消融部分)和射频佐剂l激光佐剂激光发出窄波长的光,可被特定的组织发色团吸收。可见光主要被黑色素和血红蛋白吸收,而近红外光主要被组织水吸收。激光在美学和医学方面有着独特的应用。由于其超细的光束,高能激光已被用于无刀片激光辅助原位角膜磨镶术(LASIK),术后并发症最少。激光还被用于低强度激光治疗,以刺激伤口愈合和缓解疼痛。激光还被用于皮肤表面处理、胎记和皱纹去除。已经探索了几种激光来促进皮内疫苗接种。Onikienko等人探索了一种发射两种波长(510nm和578nm)的铜蒸气激光,通过诱导持续的局部炎症来增强疫苗诱导的免疫反应,类似于化学佐剂。我们探索了非侵入性Q开关Nd:YAG532nm激光治疗来增强皮内疫苗诱导的免疫反应,并发现非侵入性激光治疗可以显著增强模型抗原OVA和流感疫苗诱导的小鼠模型的免疫反应,而不会导致可见或组织学皮肤损伤。激光治疗几乎没有诱导细胞因子的表达或免疫细胞的招募。进一步的研究发现,非炎症性激光治疗显著促进了MHCII+细胞(如巨噬细胞和DC)在皮肤内的迁移。MHCII+巨噬细胞和DC的局部运动增强可能有助于增强抗原摄取和增强获得性免疫。我们进一步发现,激光佐剂可以通过促进DC进入淋巴管和迁移到引流淋巴结来增强基于DC的抗肿瘤免疫治疗。激光佐剂被发现还能增强皮内尼古丁疫苗诱导的抗尼古丁抗体的产生。研究人员还开展了1064纳米近红外激光(NIR)增强疫苗皮内接种效果的研究。结果发现,无组织损伤的近红外治疗可以瞬时诱导一组明确的趋化因子,如CCL2和CCL20的表达,并增加表皮和真皮中DC的浓度。近红外激光治疗被发现显著提高皮内卵白蛋白和流感疫苗诱导的小鼠模型的免疫反应。进一步研究发现,近红外激光增加并激活皮肤中的Lang+和CD11b-DC亚群,并促进DC迁移到引流淋巴结,以增强疫苗诱导的免疫反应。与这项研究一致的是,皮肤中的CD103+DC亚群被发现在近红外佐剂效应中发挥关键作用。初步临床研究探索了近红外治疗在人类中的最大耐受量,并发现近红外激光促进CD1a+LC和CD11c+DC在真皮中的迁移。与非侵入性全表面激光治疗不同的是,非消融性部分激光(NAFL)也被用于增强皮内接种效果。NAFL发射高能微激光,并在皮肤表面形成微温区。在低温区域的死亡细胞招募浆细胞样树突状细胞(PDC),它可以被局部咪喹莫特激活以增强皮内流感疫苗接种效果。NAFL治疗后皮内接种流感疫苗,然后局部注射咪喹莫特,在NAFL或局部咪喹莫特存在的情况下,比皮内接种流感疫苗产生更好的免疫反应,而在AddaVax佐剂存在的情况下,肌肉接种流感疫苗产生类似的免疫反应。有趣的是,NAFL/咪喹莫特佐剂的组合流感疫苗在小鼠和猪模型中显示出良好的局部和系统安全性。另一项研究发现,在小鼠模型中,与在NAFL或单独使用咪喹莫特的情况下皮内接种疱疹病毒多肽相比,局部应用咪喹莫特和NAFL治疗,然后皮内接种疱疹病毒多肽疫苗,可以诱导最有效的疫苗特异性CD4+和CD8+T细胞反应,并对生殖器疱疹病毒感染提供最显著的保护。上述研究表明,NAFL的使用顺序、局部应用咪喹莫特和皮内接种并不是该组高免疫原性的关键,只是在皮内接种疫苗之前使用NAFL以避免疫苗抗原的潜在热灭活。进一步研究发现,NAFL刺激死亡细胞释放dsDNA,并激活STING和IRF途径来介导其佐剂作用。NAFL还被发现在小鼠和猪模型中安全地加强基于微针的流感疫苗接种。关于激光疫苗佐剂更详细分类可参考其他综述。除了仅将激光用作佐剂外,最近开发了基于激光的粉末递送系统(LPD),用于无需外部佐剂的无针经皮疫苗递送和辅助植入。LPD是基于消融部分激光(AFL)治疗产生皮肤微通道,然后局部应用粉末疫苗涂层阵列贴片,通过AFL产生的皮肤微通道将疫苗输送到皮肤。研究发现,能量为10mJ、覆盖率为10%的超脉冲CO2 AFL在促进OVA皮内免疫方面具有强大的佐剂作用。此外,基于AFL的粉末注射流感疫苗比皮内注射相同剂量的流感疫苗产生更强的抗体反应。在致命的病毒攻击后,流感疫苗的LPD也比皮内注射提供更显著的保护。AFL治疗诱导了显著的细胞因子和趋化因子的释放,并将中性粒细胞、单核细胞和嗜酸性粒细胞招募到微通道周围的组织中。由于消融了一小部分皮肤和靠近皮肤微通道的局部炎症反应,AFL治疗的皮肤没有明显的不良反应,并在几天内完全恢复。AFL的佐剂作用被认为是由于旁观者的光热效应。由于激光的脉冲宽度较长,在激光消融过程中产生的热量会扩散到周围组织,并导致周围一层薄薄的微通道组织的热损伤。dsDNA被发现从低温区域释放并诱导细胞因子表达。研究发现AFL还能激活NLRP3炎症体和Caspase1,导致IL-1β的活性释放。有趣的是,dsDNA、NLRP3炎症体和IL-1β对于激光佐剂效应是非必需的,而MyD88对激光佐剂有重要作用。与光热应激不同的是,在微针疫苗接种过程中施加的机械压力也被发现可以增强疫苗诱导的免疫反应。Ng等人发现,增加微投影阵列应用能量来诱导机械压力和细胞死亡增强疫苗(包被在微针表面)诱导的免疫反应,而不加入化学佐剂。这种方法被发现可以诱导与肿瘤坏死因子α和核因子-κB信号通路相关的基因表达。这些开创性的研究支持开发自佐剂疫苗递送技术,以便在不注射针头或化学佐剂的情况下进行更具免疫原性的疫苗接种。l射频佐剂射频(RFs)是交变电磁波。中高频(0.3-10MHz)的射频可产生组织加热,在美学和医学上有着广泛的应用。由于其热效应,RFs已被用于肿瘤治疗中的肿瘤组织消融或在皮肤表面修复中诱导新的胶原合成。为了寻求替代的物理能量来促进皮内疫苗接种,我们探索了在皮内疫苗注射之前对一小块皮肤进行双极射频治疗的潜在佐剂效果。射频治疗受到控制,以诱导显著的加热效应,但不会对皮肤造成损害。我们发现,非侵入性RF治疗诱导了一过性、低水平的局部炎症,而化学佐剂,如明矾、AddaVax和MPL,则诱导了持久且更强烈的局部炎症。RF治疗被发现显著增加真皮CD103+和CD11b-DC亚群的抗原摄取,并促进真皮DC向引流淋巴结的迁移。此外,射频治疗显著增加了引流淋巴结中传统DC和PDC的成熟。RF治疗显著增加了OVA诱导的抗体反应,佐剂效果与AddaVax相当,也增强了OVA特异性细胞免疫反应,佐剂效果与CpG佐剂相当。此外,研究发现,与0.06μg疫苗剂量的AddaVax佐剂疫苗相比,0.3μg疫苗剂量的RFA同样增强了2009年H1N1流感大流行疫苗诱导的抗体应答和保护作用。低剂量下的RFA效果较好,提示其具有优良的剂量节约效应。为了验证这一点,我们研究发现RFA可以帮助毫微克H3N2疫苗在小鼠模型中诱导有效的免疫反应和保护。考虑到先前的激光佐剂效果主要是与相对较弱的明矾佐剂进行比较,RFA是第一个与广泛使用的化学佐剂(类似MF59的AddaVax)相比的物理佐剂,可以增强流感疫苗接种。由于RFA能够帮助蛋白质或亚单位疫苗诱导细胞免疫反应,我们进一步探索了RFA在基于重组核蛋白(NP)和基质蛋白1(M1)的通用流感T细胞疫苗开发中的作用。通用流感T细胞疫苗主要针对保守的病毒细胞内抗原,如NP和M1。佐剂提供了一种方便的策略来帮助NP和M1诱导CD8+T细胞反应。我们发现RFA可以增强真皮内NP和M1诱导的CD8+T细胞反应。在小鼠模型中,在RFA存在的情况下,皮内NP/M1免疫对致死性病毒攻击具有显著的保护作用。在加强NP/M1免疫方面,RFA显示出与AddaVax相似的佐剂作用。最近发现,RFA还可以扩大流感疫苗诱导的针对异源病毒的免疫反应。这种交叉保护免疫可能是由于它诱导了交叉保护的细胞毒性T淋巴细胞。RFA还有效促进了雄性和雌性小鼠的H5N1流感疫苗皮内免疫。含有RFA的H5N1流感疫苗皮内接种(RFA/ID)诱导的血清血凝抑制(HI)效价和免疫球蛋白抗体效价显著高于单独皮内疫苗(ID)。在致死性病毒攻击后,RFA/ID组而不是ID组显示出显著的肺病毒滴度降低。有趣的是,与ID组相比,RFA/ID组攻击后肺组织中分泌肿瘤坏死因子α和干扰素γ的CD4+和CD8+T细胞显著增加。RFA/ID组对体重减轻也有显著的保护作用。RFA/ID组小鼠不分性别全部存活,ID组部分小鼠存活(雌鼠占25%,雄鼠占75%)。有趣的是,在这项研究中,与AddaVax佐剂相比,RFA对促进H5N1流感疫苗接种的作用较弱。比较组织蛋白质组学最近被用来鉴定RFA治疗后独特表达的蛋白质。我们发现,与化学佐剂处理(MPL、AddaVax、Alum、MPL/Alum)相比,RFA诱导了14种唯一表达的蛋白质。在唯一表达的蛋白质中,热休克蛋白70(HSP70)在RFA效应中发挥了关键作用。验证研究发现,与野生型小鼠相比,HSP70基因敲除小鼠显著降低了促进OVA和pdm09疫苗接种的RFA效应。有趣的是,这项研究发现RFA引起的局部组织蛋白质组变化最小,而AddaVax佐剂引起的局部组织蛋白质组变化最大。RFA对局部组织蛋白质组的最小变化与其诱导的一过性和低水平的局部炎症是一致的。l物理佐剂的优点物理佐剂将物理能量短暂地应用于皮肤表面,然后皮内注射疫苗,以增强疫苗诱导的免疫反应(图5)。物理佐剂主要由医疗器械输送,可在室温下保存,无需冷链储存。这与化学佐剂不同,化学佐剂通常与疫苗一起保存,并需要冷链储存。物理佐剂设备可以重复使用,每次应用的成本可能类似于化学佐剂。物理佐剂不需要与疫苗混合,因此不需要改变或修改疫苗制造或给药过程。物理佐剂不太可能引起显著的全身性或长期副作用,这可能是化学佐剂的一个问题。这是因为物理佐剂的作用主要局限于治疗部位,而化学佐剂可以迁移到引流淋巴结,甚至到达体循环,从而广泛激活免疫系统。物理佐剂可诱导良好控制的组织应激(例如,热),并允许皮肤快速恢复,而不会产生持久的影响。物理佐剂经常引起短暂的、低水平的局部炎症,而化学佐剂往往引起持续的、更强烈的局部炎症。物理佐剂主要是改变局部环境以增加DC的运动性,诱导内源性危险信号的释放以增强抗原摄取和DC成熟,或促进DC迁移到引流淋巴结以增强疫苗诱导的免疫反应(图5)。经过适当的开发,物理佐剂的效果可以与化学佐剂一样强大。图5.化学佐剂和物理佐剂在皮内接种中的不同作用机制。皮内给药后,化学佐剂通过与PRRs结合直接激活DC,诱导DC成熟,诱导DC释放细胞因子/趋化因子,招募天然免疫细胞,或促进DC向引流淋巴结迁移,以增强疫苗诱导的免疫应答。物理佐剂传递激光或射频能量,诱导组织应激释放危险信号,刺激DC成熟,诱导DC释放细胞因子/趋化因子,并招募先天免疫细胞。物理佐剂还可以增加DC在真皮组织中的运动性,并促进DC迁移到引流淋巴结,以增强疫苗诱导的免疫反应。7.专家评论皮肤已被公认为疫苗接种的高免疫原性部位。与肌肉接种相比,皮内接种疫苗通常能诱导更强的免疫反应,在提高疫苗效力或节省疫苗剂量方面有很大作用。事实上,已有几种疫苗获得了皮内接种的许可,其中包括流感疫苗、狂犬病疫苗和最近的猴痘疫苗。然而,皮内接种面临着诱发严重局部反应的风险。目前获批的大多数佐剂都不适合皮内接种,因为它们具有诱发严重局部反应的高风险。我们需要新型佐剂来安全地增强皮内接种效果,同时避免皮肤出现明显的反应。最近的研究发现了几种有可能安全增强疫苗皮内接种效果的化学佐剂,包括CpG1018、GLA-AF、局部咪喹莫特、poly(I:C)、cGAMP和PCPP。CpG1018已被授权用于促进肌肉接种乙型肝炎疫苗,而GLA-AF和局部咪喹莫特已在临床试验中用于增强皮内接种流感疫苗。由于这些佐剂在人体中的安全性已得到证实,因此可随时用于皮内疫苗接种。poly(I:C)已在临床试验中被用于增强肿瘤内、肌内或皮下注射后的抗肿瘤免疫力。poly(I:C)其总体良好的耐受性支持对其进行评估,以增强皮内疫苗接种。皮内接种佐剂疫苗的好处是可以进一步提高疫苗效力,节省更多疫苗剂量,并有可能诱导交叉保护性免疫。流感疫苗可作为测试皮内佐剂接种的良好目标,以解决疫苗失配问题,因为疫苗失配会大大降低疫苗效力,或通过节省疫苗剂量来解决每年流感疫苗生产的负担。在批准皮内佐剂疫苗供大众使用时,需要仔细评估其效益/风险比。除化学佐剂外,人们还探索了不同类型的物理佐剂(激光、射频),以安全地促进皮内疫苗接种。物理佐剂是一种相对较新的佐剂类型,它利用物理能量诱导组织应激以促进皮内接种。虽然物理佐剂提高人类疫苗效力的潜力仍有待探索,但最近发现内源性危险信号在化学佐剂效应中发挥关键作用(如明矾、MF59),这支持了物理佐剂的发展,以诱导组织应激和释放内源性危险信号来促进疫苗接种。最近在临床前动物模型中研究了物理佐剂的概念及其增强皮内疫苗接种的安全性和有效性。这些研究支持物理佐剂在促进皮内疫苗接种方面具有良好的安全性和有效性。尽管取得了这些令人鼓舞的成果,但物理佐剂在人类使用方面仍面临着一些挑战。首先,物理佐剂是依靠设备的。目前,还没有指导物理佐剂开发和审批的指导方针。其次,物理佐剂的开发需要设备制造商和疫苗公司的共同努力,还需要引入佐剂开发和医疗设备审批方面的监管机构。多方的参与可能会给物理佐剂的开发带来新的挑战。第三,使用目前的物理佐剂进行皮内接种需要两个步骤:物理佐剂处理和皮内接种。与化学佐剂结合疫苗接种相比,两步接种过程需要花费更多时间,而且公众对这种新型疫苗接种的接受程度还有待进一步探讨。佐剂的开发在21世纪受到极大关注。在批准新佐剂供人类使用和阐明疫苗佐剂的内在机制方面取得了重大进展。所有这些努力都有望促进安全的化学和物理佐剂的开发,从而增强皮内接种疫苗的效果。参考来源:Chen X. Emerging adjuvants for intradermal vaccination. Int J Pharm. 2023 Feb 5;632:122559. doi: 10.1016/j.ijpharm.2022.122559. Epub 2022 Dec 28. PMID: 36586639; PMCID: PMC9794530.推荐学习:疫苗佐剂概述:发展现状和未来展望基于TLR激动剂佐剂的设计疫苗佐剂:选择标准、与免疫应答相关的作用机制和未来方向识别微信二维码,添加生物制品圈小编,符合条件者即可加入生物制品微信群!请注明:姓名+研究方向!版权声明本公众号所有转载文章系出于传递更多信息之目的,且明确注明来源和作者,不希望被转载的媒体或个人可与我们联系(cbplib@163.com),我们将立即进行删除处理。所有文章仅代表作者观点,不代表本站立场。
疫苗
2023-04-19
关注并星标CPHI制药在线 阿必鲁肽(Albiglutide)是一种GLP-1受体激动剂,用于治疗2型糖尿病。它最初由GlaxoSmithKline(GSK)公司开发,并于2014年获得美国FDA批准上市。它是一种长效药物,每周注射一次,可帮助患者更好地管理他们的血糖水平,曾经被批准在美国和欧洲等地销售。 该药物的研发历程可以追溯到2004年,当时GSK公司开始了一项针对2型糖尿病的药物研究项目,旨在开发一种长效的GLP-1受体激动剂。在接下来的几年中,该公司进行了大量的药物筛选和临床试验,最终选择了阿必鲁肽作为最有前途的候选药物。在临床试验中,阿必鲁肽被证明能够显著降低患者的血糖水平,并具有良好的安全性和耐受性。在2014年,阿必鲁肽获得了美国FDA的批准,并在全球范围内上市。 GSK突然放弃了阿必鲁肽的销售,原因销售不佳? 尽管阿必鲁肽在治疗2型糖尿病方面取得了一定的成功。但是,比较突然的是,2017年底,葛兰素史克表示将停止一切对阿比鲁肽的进一步研发、制造和销售活动,将其正式退出医药市场。并且,GSK给出的原因是:由于市场竞争激烈,以及其他更具创新性的药物的出现,它的销售额并不如预期。这很奇怪,因为事实上,阿必鲁肽此时的疗效已经显现,不仅具有对心血管有益,还比先前的同类GLP-1抑制剂药品--艾塞那肽的效果更具优势。目前,阿必鲁肽早已经在主流国家停止销售。GSK在阿必鲁肽投入市场三年后,在没有发现任何问题的前提下,突然停止使用,引起了人们的无数猜疑。 前车之鉴:GSK历经多起药品索赔,最高30亿美元 历史上,GSK医药公司曾经历过一系列巨额的药品不良反应索赔官司,以解决数千名患者的索赔。例如,GSK医药公司曾经赔付的最大一笔赔偿金是在2012年,该公司同意支付30亿美元的赔偿金,解决了美国政府对其多种药品的诉讼。其中包括其生产的多种药物的安全性问题,这是有史以来医药公司支付的最高罚款之一。以解决因该公司非法推广某些处方药、未能报告某些安全数据以及涉嫌虚假价格报告行为的民事责任而产生的刑事和民事责任。该决议是美国历史上最大的医疗保健欺诈和解,也是制药公司有史以来最大的付款。 这些事件引起了公众对制药公司的信任度的担忧,并促使监管机构加强了对制药行业的监管。最后,GSK也承诺采取措施改进其商业行为,并强调其致力于研发安全有效的药物,为患者带来更好的健康结果。 当年的阿必鲁肽,有什么严重风险出现吗? 关于不良反应 阿必鲁肽常见的不良反应包括恶心、呕吐、腹泻、便秘、胃肠道不适、头痛、注射部位疼痛、瘙痒和皮疹等。这些不良反应通常是轻度或中度的,并在使用药物一段时间后逐渐减轻或消失。阿必鲁肽也可能引起一些更严重的不良反应,例如低血糖、胰腺炎、过敏反应和肝功能异常等。 阿必鲁肽的药品说明书中列出的严重不良反应包括低血糖、胰腺炎、甲状腺C细胞瘤、过敏反应和肝功能异常等。其中,最明确的可能是,阿必鲁肽可能会增加患甲状腺C细胞瘤的风险,这是一种罕见的甲状腺癌。成因可能是:GLP-1受体也存在于甲状腺C细胞中,这些细胞是甲状腺中的一种神经内分泌细胞,可以分泌钙调素。如果甲状腺C细胞突变导致肿瘤的形成,可能会导致甲状腺C细胞瘤的发生。阿必鲁肽可能会在甲状腺C细胞中诱导增殖,进而导致甲状腺C细胞瘤的发生。总体来说,甲状腺C细胞瘤是一种罕见的肿瘤,只占所有甲状腺癌的约5%。 小结 迄今为止,GSK公司当年为何突然放弃前景无限的阿必鲁肽的销售和研发,背后真实的原因我们仍未可知。先前已有的数据表明阿必鲁肽的短期潜在风险通常被认为是相对较低的,长期风险未知。因此,需要注意的是,对于GLP-1激动剂类药物,当前仍旧需要在医生的指导下使用药物并密切关注任何长期的不良反应。 参考资料: GlaxoSmithKline to Plead Guilty and Pay $3 Billion to Resolve Fraud Allegations and Failure to Report Safety Data | OPA | Department of Justice 智药研习社近期课程报名来源:CPHI制药在线声明:本文仅代表作者观点,并不代表制药在线立场。本网站内容仅出于传递更多信息之目的。如需转载,请务必注明文章来源和作者。投稿邮箱:Kelly.Xiao@imsinoexpo.com▼更多制药资讯,请关注CPHI制药在线▼点击阅读原文,进入智药研习社~
引进/卖出上市批准
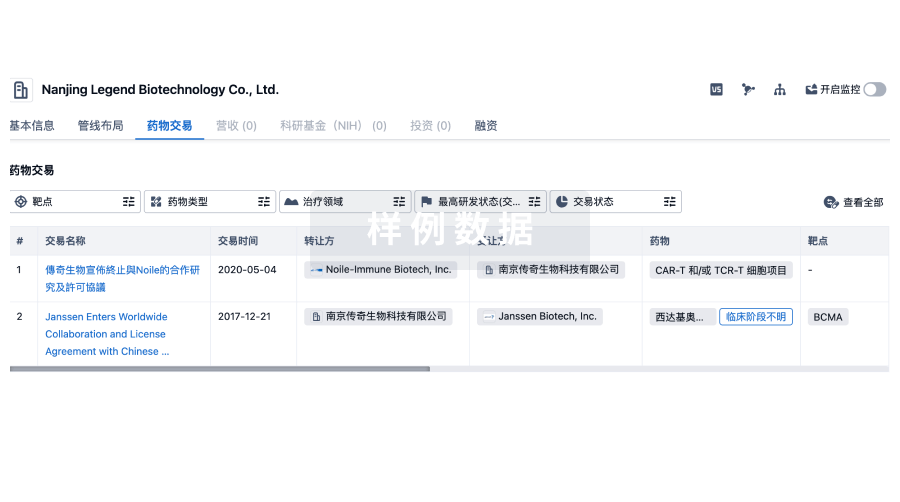
100 项与 GlaxoSmithKline (GSK) SRL 相关的药物交易
登录后查看更多信息
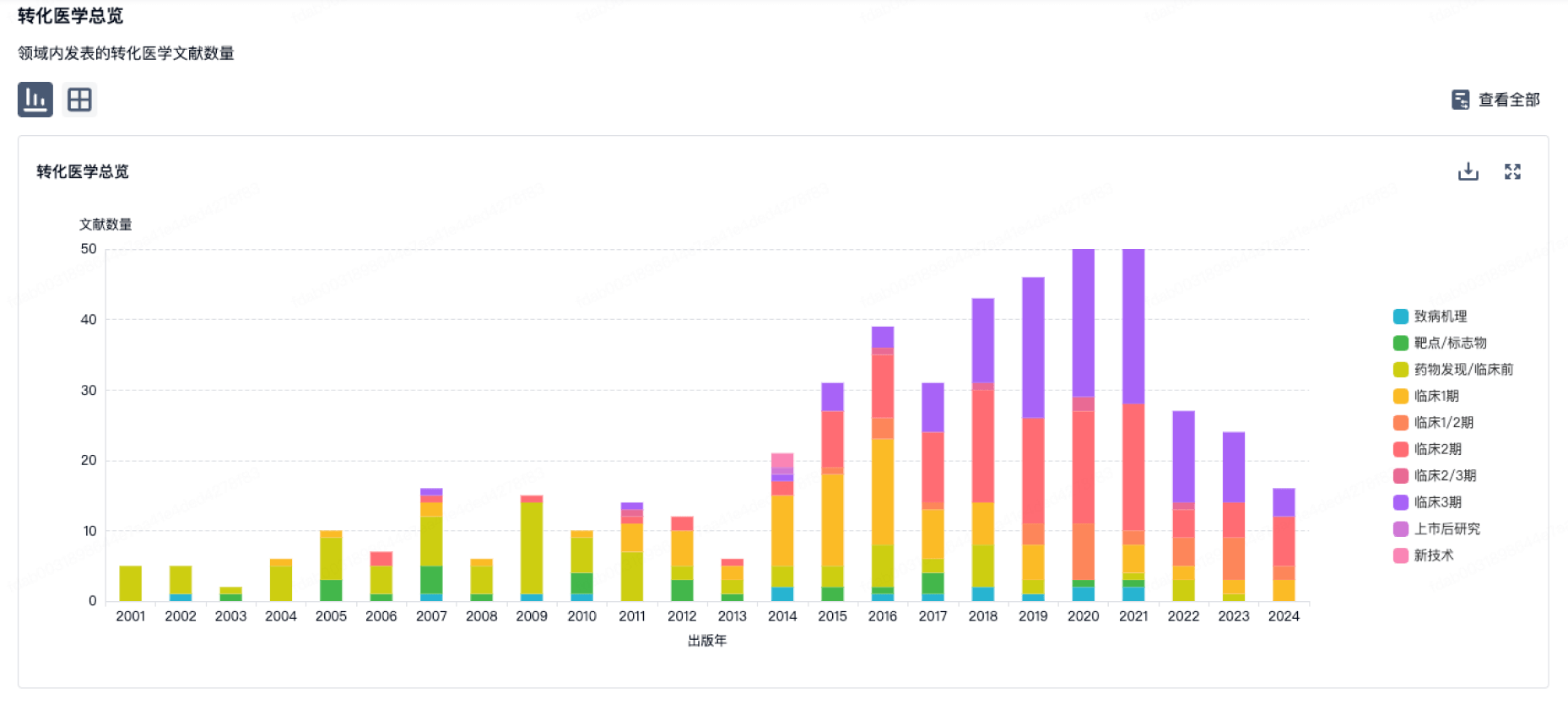
100 项与 GlaxoSmithKline (GSK) SRL 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2026年02月08日管线快照
无数据报导
登录后保持更新
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
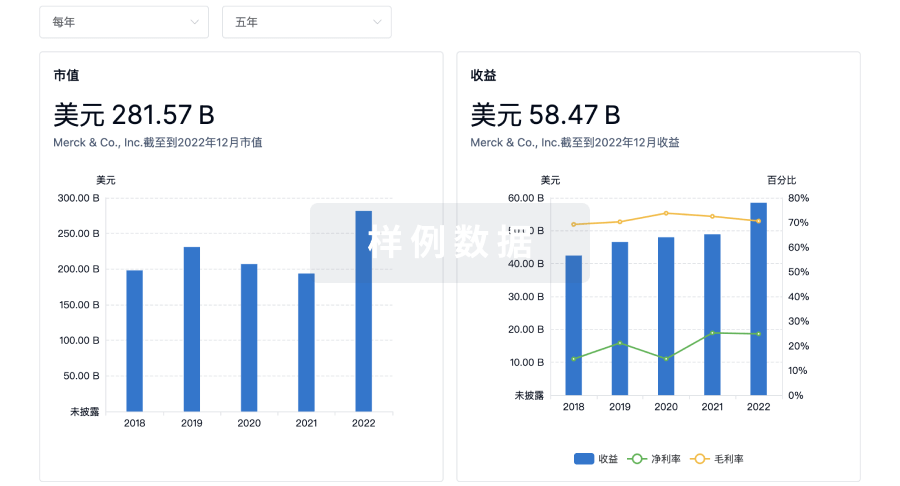
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
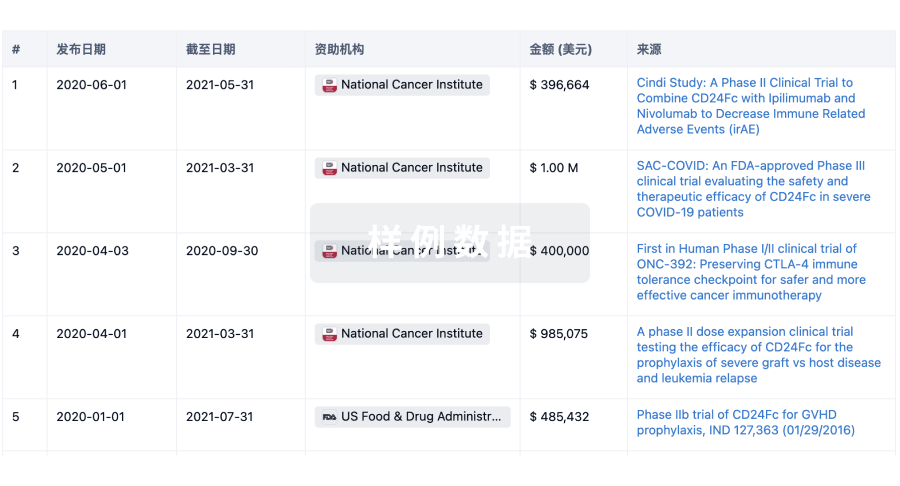
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
生物医药百科问答
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用