预约演示
更新于:2026-05-25
Nanjing Tuge Medical Technology Co., Ltd.
更新于:2026-05-25
概览
关联
100 项与 南京图格医疗科技有限公司 相关的临床结果
登录后查看更多信息
0 项与 南京图格医疗科技有限公司 相关的专利(医药)
登录后查看更多信息
13
项与 南京图格医疗科技有限公司 相关的新闻(医药)2026-05-01
伙伴企业动态速览
◼ 昭衍新药与英矽智能战略合作签约
◼ 健适收购内窥镜企业图格医疗
◼ 阿诺医药宣布约1.5亿美元融资
◼ 恒宇医疗完成超亿元融资
◼ 瑞尔刀落户长春肿瘤医院
◼ 星测未来与中交水规院达成战略合作
◼ 研微半导体新金属ALD产品再度交付客户
◼ 钧天一号04A星高质量卫星图像发布
◼ 勇芯科技完成近亿元A轮融资
◼ 意优科技机器人关节项目落户无锡
◼ 自变量机器人完成近20亿元B轮融资并发布全球首个世界统一模型
昭衍新药与英矽智能战略合作签约
近日,昭衍新药与英矽智能签署战略合作协议。双方将围绕创新药非临床研究、实验动物资源保障、AI驱动的生命科学模型(Life model)研发等领域建立长期、深度、全方位的战略合作伙伴关系。
英矽智能是全球领先的生成式AI驱动药物研发企业。公司依托自主研发的Pharma.AI平台,在靶点发现与评估、药物分子设计及优化、生命科学基础研究等核心领域拥有国际领先技术实力。公司已有12条自研管线获得临床试验许可,其中进展最为领先的管线Rentosertib已完成临床II期概念验证,持续引领AI赋能药物研发的产业化发展。
昭衍新药是国内领先的医药研发服务企业,具备覆盖药物发现、非临床评价、临床试验、药品质量检定等药物研发全产业链一体化服务能力。凭借深厚的行业积淀、规模化稳定的全球资源布局及完备的国际化合规资质,能够高标准、全方位支撑各类创新药研发项目,保障全流程高质量落地与稳定交付。
健适收购内窥镜企业图格医疗
近日,健适医疗对外宣布,公司已完成对南京图格医疗科技有限公司(简称图格医疗)的控股权收购,后者是国内医用内窥镜成像技术领域的硬科技企业。此次战略并购后,图格品牌将成为健适大外科业务版图中的重要组成部分,共同构建更完善的微创外科产品生态。
健适医疗成立于2019年,是一家新兴的医疗科技企业。健适致力于打造一个开放的医疗科技平台:通过全球创新网络、本土研发生产以及持续升级的商业化能力,健适深耕于广袤的中国市场,为国内的医患提供优质的医疗科技产品和服务。
阿诺医药宣布约1.5亿美元融资
近日,阿诺医药集团有限公司宣布已就一项上市公司私募股权融资(PIPE)签署证券购买协议,预计融资总额约为1.5亿美元(未扣除配售代理及其他相关费用)。
阿诺医药(纳斯达克股票代码:ANL)是一家全球性临床阶段生物制药公司,致力于开发创新的癌症治疗方案,公司正打造丰富强劲的候选药物管线,涵盖两大关键治疗领域:1)RAS通路精准靶向疗法:包括口服泛RAS (ON) 抑制剂AN9025,以及靶向CEACAM5 的抗体药物偶联物(ADC)AN4035,将强效泛 RAS (ON) 抑制剂高效递送至肿瘤部位。2) 下一代PD-1/L1通路调控免疫疗法:包括可同时调节T细胞与抗原呈递细胞的多功能融合蛋白AN8025及首创口服PD-L1抑制剂AN4005。
恒宇医疗完成超亿元融资
日前,专注于血管介入影像的企业——天津恒宇医疗科技有限公司(简称“恒宇医疗”)宣布完成超亿元融资。本轮融资由中金资本和锡创投领投。
恒宇医疗成立于2016年,专注于血管内超声(IVUS)和光学相干断层成像(OCT)设备的自主研发、生产和销售,掌握光学、超声及导管平台技术,并提供IVUS/OCT一体机、基于影像的无创FFR平台、OCT-NIRS多模态成像及血管内激光斑块消蚀等产品,致力于构建覆盖泛血管疾病精准诊疗的全链条解决方案。
瑞尔医疗产品“瑞尔刀”
落户长春肿瘤医院
日前,立体定向放射治疗机器人——全国第二台瑞尔刀正式在长春肿瘤医院开机启用,同步举行瑞尔刀培训基地揭牌仪式。
江苏瑞尔医疗科技有限公司自主研发的“瑞尔刀”(RayerKnife)是国内首台实现全国产化的X射线立体定向放射治疗设备。2024年9月,RayerKnife瑞尔刀获批NMPA产品注册证。
星测未来与中交水规院达成战略合作
近日,星测未来科技(北京)有限责任公司与中交水运规划设计院有限公司(下称“中交水规院”)在北京正式签署战略合作框架协议。双方将发挥各自在太空算力与交通水运规划设计领域的核心优势,共同打造 "陆海空天" 一体化交通解决方案,助力交通强国、海洋强国、数字中国建设。
星测未来成立于2020年,专注卫星智能化,赋能高效空天信息服务。公司致力于“AI+航天”的深度融合,为通信、遥感、态势感知、太空计算等领域提供领先的天基智能解决方案。
研微半导体新金属ALD产品
再度交付头部客户
日前,研微(江苏)半导体科技有限公司thermal ALD Spritz 系列金属ALD再度交付头部存储客户。公司集数十位国内外技术专家,深耕ALD等薄膜沉积技术,合力解决产品及客户技术难题。同产品机台的多次重复交付也展现公司的品质承诺及量产能力。
研微半导体成立于 2022 年,坐落于无锡经济开发区。公司致力于研发、生产和销售具有自主知识产权且具备国际竞争力的半导体设备,专注于原子层沉积(ALD)、 Si 外延沉积(SI EPI)、等离子体化学气相沉积(PECVD)、SiC 外延(SiC EPI)以及原子层刻蚀(ALE)等设备及工艺技术的研发。
钧天航宇钧天一号04A星
高质量多模式卫星图发布
日前,钧天一号04A星在酒泉卫星发射中心成功发射,卫星顺利进入预定轨道后,仅用3天即获取首幅清晰影像,圆满完成入轨初期各项测试任务。
北京钧天航宇技术有限公司成立于2021年12月31日,总部位于北京市,是一家以商业卫星研发制造为主营业务的国家高新技术企业,致力于以颠覆式创新方式研发高质量低成本的新一代商业卫星。
勇芯科技完成近亿元A轮融资
日前,无锡勇芯科技有限公司宣布完成近亿元A轮融资,资金将聚焦核心技术迭代、供应链生态整合、产线扩容及团队扩充,进一步巩固公司在AI硬件Chiplet方向的技术与产业化优势。
勇芯科技创立于2018年,坐落于江苏无锡,是国内专注AIoT场景、提供专业Chiplet芯片级整体解决方案的高新技术企业,2025年成功获评无锡市准独角兽培育企业,成长潜力备受地方产业与资本市场认可。
意优科技机器人关节项目落子无锡
近日,意优科技机器人关节全自动产业化基地项目正式签约落户无锡锡东新城。据悉,项目总投资10亿元,聚焦人形机器人核心关节研发与自动化生产,将打造全国最大的人形机器人关节自动化生产基地,为无锡高端装备制造与未来产业高质量发展再添强劲动力。
意优科技是国内领先的一体化伺服关节提供商。公司推出的proServo系列关节模组,性能优异、稳定性强,被广泛应用于人形机器人、协作机械臂、工业自动化等高端装备领域,企业估值达20亿元,产品已进入欧美、日韩等海外高端市场。
自变量机器人获近20亿元B轮融资
并发布全球首个世界统一模型
近日,自变量机器人完成近20亿元B轮融资,同期发布全球首个世界统一模型——具身智能基础模型 WALL-B。这是全球首个基于世界统一模型架构(World Unified Model,WUM) 的具身智能基础模型,标志着具身基础模型从VLA架构向原生多模态融合架构的重大跨越。
自变量机器人科技有限公司(X Square Robot)成立于 2023年12月,聚焦自研“通用具身智能大模型”,以真实世界数据为主要数据来源,构建具备精细操作能力的通用机器人,是国内最早采用完全端到端路径实现通用具身智能大模型的公司之一。
【会员动态】无锡市创业投资协会会员单位多位女性入选年度优秀女性投资人榜单
【行业数据】2025年无锡市PE/VC行业报告
【会员动态】协会多家会员单位荣登 “融中2025年度中国股权投资年度排名榜单”
【会员动态】协会多家会员单位入围母基金研究中心2025年度专项榜单
编辑/校对:江叶
审核:雨砚
无锡市创业投资协会
扫描二维码,加入我们
版权声明:除非特别说明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流之目的。转载的稿件版权归原作者或机构所有。如有侵权,请及时联系我们删除。
END
协会地址:无锡市滨湖区建筑西路583号2204/2205/2206
联系方式:wuxi_xiehui@163.com
并购临床2期
2026-05-01
·锡创投
伙伴企业动态速览
◼ 昭衍新药与英矽智能战略合作签约
◼ 健适收购内窥镜企业图格医疗
◼ 阿诺医药宣布约1.5亿美元融资
◼ 恒宇医疗完成超亿元融资
◼ 瑞尔刀落户长春肿瘤医院
◼ 星测未来与中交水规院达成战略合作
◼ 研微半导体新金属ALD产品再度交付客户
◼ 钧天一号04A星高质量卫星图像发布
◼ 勇芯科技完成近亿元A轮融资
◼ 意优科技机器人关节项目落户无锡
◼ 自变量机器人完成近20亿元B轮融资并发布全球首个世界统一模型
昭衍新药与英矽智能战略合作签约
近日,昭衍新药与英矽智能签署战略合作协议。双方将围绕创新药非临床研究、实验动物资源保障、AI驱动的生命科学模型(Life model)研发等领域建立长期、深度、全方位的战略合作伙伴关系。
英矽智能是全球领先的生成式AI驱动药物研发企业。公司依托自主研发的Pharma.AI平台,在靶点发现与评估、药物分子设计及优化、生命科学基础研究等核心领域拥有国际领先技术实力。公司已有12条自研管线获得临床试验许可,其中进展最为领先的管线Rentosertib已完成临床II期概念验证,持续引领AI赋能药物研发的产业化发展。
昭衍新药是国内领先的医药研发服务企业,具备覆盖药物发现、非临床评价、临床试验、药品质量检定等药物研发全产业链一体化服务能力。凭借深厚的行业积淀、规模化稳定的全球资源布局及完备的国际化合规资质,能够高标准、全方位支撑各类创新药研发项目,保障全流程高质量落地与稳定交付。
健适收购内窥镜企业图格医疗
近日,健适医疗对外宣布,公司已完成对南京图格医疗科技有限公司(简称图格医疗)的控股权收购,后者是国内医用内窥镜成像技术领域的硬科技企业。此次战略并购后,图格品牌将成为健适大外科业务版图中的重要组成部分,共同构建更完善的微创外科产品生态。
健适医疗成立于2019年,是一家新兴的医疗科技企业。健适致力于打造一个开放的医疗科技平台:通过全球创新网络、本土研发生产以及持续升级的商业化能力,健适深耕于广袤的中国市场,为国内的医患提供优质的医疗科技产品和服务。
阿诺医药宣布约1.5亿美元融资
近日,阿诺医药集团有限公司宣布已就一项上市公司私募股权融资(PIPE)签署证券购买协议,预计融资总额约为1.5亿美元(未扣除配售代理及其他相关费用)。
阿诺医药(纳斯达克股票代码:ANL)是一家全球性临床阶段生物制药公司,致力于开发创新的癌症治疗方案,公司正打造丰富强劲的候选药物管线,涵盖两大关键治疗领域:1)RAS通路精准靶向疗法:包括口服泛RAS (ON) 抑制剂AN9025,以及靶向CEACAM5 的抗体药物偶联物(ADC)AN4035,将强效泛 RAS (ON) 抑制剂高效递送至肿瘤部位。2) 下一代PD-1/L1通路调控免疫疗法:包括可同时调节T细胞与抗原呈递细胞的多功能融合蛋白AN8025及首创口服PD-L1抑制剂AN4005。
恒宇医疗完成超亿元融资
日前,专注于血管介入影像的企业——天津恒宇医疗科技有限公司(简称“恒宇医疗”)宣布完成超亿元融资。本轮融资由中金资本和锡创投领投。
恒宇医疗成立于2016年,专注于血管内超声(IVUS)和光学相干断层成像(OCT)设备的自主研发、生产和销售,掌握光学、超声及导管平台技术,并提供IVUS/OCT一体机、基于影像的无创FFR平台、OCT-NIRS多模态成像及血管内激光斑块消蚀等产品,致力于构建覆盖泛血管疾病精准诊疗的全链条解决方案。
瑞尔医疗产品“瑞尔刀”
落户长春肿瘤医院
日前,立体定向放射治疗机器人——全国第二台瑞尔刀正式在长春肿瘤医院开机启用,同步举行瑞尔刀培训基地揭牌仪式。
江苏瑞尔医疗科技有限公司自主研发的“瑞尔刀”(RayerKnife)是国内首台实现全国产化的X射线立体定向放射治疗设备。2024年9月,RayerKnife瑞尔刀获批NMPA产品注册证。
星测未来与中交水规院达成战略合作
近日,星测未来科技(北京)有限责任公司与中交水运规划设计院有限公司(下称“中交水规院”)在北京正式签署战略合作框架协议。双方将发挥各自在太空算力与交通水运规划设计领域的核心优势,共同打造 "陆海空天" 一体化交通解决方案,助力交通强国、海洋强国、数字中国建设。
星测未来成立于2020年,专注卫星智能化,赋能高效空天信息服务。公司致力于“AI+航天”的深度融合,为通信、遥感、态势感知、太空计算等领域提供领先的天基智能解决方案。
研微半导体新金属ALD产品
再度交付头部客户
日前,研微(江苏)半导体科技有限公司thermal ALD Spritz 系列金属ALD再度交付头部存储客户。公司集数十位国内外技术专家,深耕ALD等薄膜沉积技术,合力解决产品及客户技术难题。同产品机台的多次重复交付也展现公司的品质承诺及量产能力。
研微半导体成立于 2022 年,坐落于无锡经济开发区。公司致力于研发、生产和销售具有自主知识产权且具备国际竞争力的半导体设备,专注于原子层沉积(ALD)、 Si 外延沉积(SI EPI)、等离子体化学气相沉积(PECVD)、SiC 外延(SiC EPI)以及原子层刻蚀(ALE)等设备及工艺技术的研发。
钧天航宇钧天一号04A星
高质量多模式卫星图发布
日前,钧天一号04A星在酒泉卫星发射中心成功发射,卫星顺利进入预定轨道后,仅用3天即获取首幅清晰影像,圆满完成入轨初期各项测试任务。
北京钧天航宇技术有限公司成立于2021年12月31日,总部位于北京市,是一家以商业卫星研发制造为主营业务的国家高新技术企业,致力于以颠覆式创新方式研发高质量低成本的新一代商业卫星。
勇芯科技完成近亿元A轮融资
日前,无锡勇芯科技有限公司宣布完成近亿元A轮融资,资金将聚焦核心技术迭代、供应链生态整合、产线扩容及团队扩充,进一步巩固公司在AI硬件Chiplet方向的技术与产业化优势。
勇芯科技创立于2018年,坐落于江苏无锡,是国内专注AIoT场景、提供专业Chiplet芯片级整体解决方案的高新技术企业,2025年成功获评无锡市准独角兽培育企业,成长潜力备受地方产业与资本市场认可。
意优科技机器人关节项目落子无锡
近日,意优科技机器人关节全自动产业化基地项目正式签约落户无锡锡东新城。据悉,项目总投资10亿元,聚焦人形机器人核心关节研发与自动化生产,将打造全国最大的人形机器人关节自动化生产基地,为无锡高端装备制造与未来产业高质量发展再添强劲动力。
意优科技是国内领先的一体化伺服关节提供商。公司推出的proServo系列关节模组,性能优异、稳定性强,被广泛应用于人形机器人、协作机械臂、工业自动化等高端装备领域,企业估值达20亿元,产品已进入欧美、日韩等海外高端市场。
自变量机器人获近20亿元B轮融资
并发布全球首个世界统一模型
近日,自变量机器人完成近20亿元B轮融资,同期发布全球首个世界统一模型——具身智能基础模型 WALL-B。这是全球首个基于世界统一模型架构(World Unified Model,WUM) 的具身智能基础模型,标志着具身基础模型从VLA架构向原生多模态融合架构的重大跨越。
自变量机器人科技有限公司(X Square Robot)成立于 2023年12月,聚焦自研“通用具身智能大模型”,以真实世界数据为主要数据来源,构建具备精细操作能力的通用机器人,是国内最早采用完全端到端路径实现通用具身智能大模型的公司之一。
2026-04-13
数据来源:企业官方公告、FDA/NMPA/CDE、ClinicalTrials.gov、Nature/Science/Cell/NEJM/Lancet、彭博、路透 一、国内外药企与Biotech重大事件(本周) (一)跨国药企 1. 礼来(LLY.US)- 4月7日:与英矽智能达成27.5亿美元AI制药合作(首付1.15亿),聚焦自免与罕见病靶点发现- 来源:礼来公告、FTC披露
2. 罗氏(RHHBY.US)- 4月8日:CD20/CD3双抗Mosunetuzumab获FDA批准用于复发难治滤泡性淋巴瘤(2线+),ORR 80%,CR 60%- 来源:FDA、罗氏官网
3. 诺和诺德(NVO.US)- 4月9日:口服司美格鲁肽III期OASIS 1顶线:52周减重14.7%(vs安慰剂2.1%,P<0.0001),胃肠道AE≤4%- 来源:诺和诺德公告 (二)国内药企/Biotech 1. 恒瑞医药(600276.SH)- 4月6日:HER2 ADC SHR-A1811 III期达主要终点,HER2阳性晚期乳腺癌PFS显著优于拉帕替尼+卡培他滨(HR=0.38,P<0.001)- 来源:公司公告2. 百利天恒(688506.SH)- 4月10日:EGFR/HER3双抗ADC IcaBRen获FDA突破性疗法认定(三阴乳腺癌),全球首个该靶点双抗ADC获BTD- 来源:公司公告
3. 汇伦医药- 4月8日:递交港交所IPO申请,主营小分子靶向(JAK、FGFR),2025年营收6.8亿元- 来源:港交所披露
4. 映恩生物- 4月9日:帕康曲妥珠单抗(HER2 ADC) NMPA申报上市(乳腺癌),为公司首款商业化ADC- 来源:CDE官网 二、重点靶点与赛道临床进展(本周数据) (一)肿瘤 1. KRAS G12C(NSCLC)- 药物:JDQ-4334(劲方医药)- 方案:II期,既往经治KRAS G12C突变NSCLC- 结果:ORR 42.3%,DCR 87.2%,mPFS 7.8个月;3级AE 8.5%- 结论:国产KRAS抑制剂疗效追平国际,安全性更优- 来源:AACR 2026(4.8)、公司公告
2. Claudin 18.2(胃癌/胃食管结合部癌)- 药物:OS-102(石药集团)- 方案:II期,一线联合化疗- 结果:ORR 68.4%(vs化疗41.2%,P=0.003),mPFS 10.2m vs 5.7m(HR=0.41,P<0.001)- 结论:Claudin 18.2单抗联合化疗显著增效- 来源:公司公告(4.7)
3. BCMA/CD3双抗(多发性骨髓瘤)- 药物:TNB-383B(德琪医药)- 方案:I/II期,≥3线复发难治- 结果:ORR 71.4%,sCR 28.6%,mDOR 11.5个月- 结论:国产双抗深度缓解率提升- 来源:EHA 2026摘要(4.10) (二)自免 - IL-17A/F(银屑病关节炎)- 药物:Bimekizumab(优时比)- 方案:III期 BE HAPPY,500例- 结果:ACR20应答率71.2% vs 43.5%(P<0.001);关节结构损伤进展降62%(P=0.002)- 来源:NEJM(4.9) (三)代谢(GLP-1) - 诺和诺德口服司美格鲁肽:52周减重14.7%,HbA1c降1.9%;恶心、腹泻发生率<10%,3级≤4%- 礼来Orforglipron(口服GLP-1):中国III期顶线:52周减重15.1%(vs安慰剂2.4%,P<0.0001),申报NDA- 来源:礼来中国公告 (四)神经/抗衰老 - 阿尔茨海默病(Tau抗体)- 药物:Tilzapirumab(卫材/渤健)- 方案:III期 TAURIEL,早期AD- 结果:认知衰退速率降28%(P=0.004),Tau负荷降35%;ARIA-E 3.2%- 来源:Lancet(4.11)- 抗衰老(Senolytics)- 药物:UBX1325(Unity Biotech)- 方案:II期,老年虚弱综合征- 结果:6分钟步行距离提升22%(P<0.01),炎症因子IL-6降42%(P<0.001)- 来源:Nature Aging(4.8) 三、器械与前沿技术进展(本周) (一)脑机接口 - Neuralink- 4月7日:第四代侵入式BCI FDA批准临床,用于ALS患者,意念打字速度达65字符/分钟- 来源:Neuralink公告- 博睿康- 4月11日:无创脑电AI解码系统获NMPA批准,用于癫痫灶定位,准确率92.3%- 来源:NMPA、公司公告 (二)医疗AI - 香港科大(Nature Cancer, 4.10)- PRET模型:无需训练、泛癌种病理诊断,准确率94.7%,覆盖肺癌/胃癌/乳腺癌- 推想医疗- 4月8日:AI肺结节诊断系统纳入全国医保,报销70%-85%,覆盖837家三甲 (三)手术机器人/介入器械 - 微创机器人(2252.HK)- 4月9日:图迈骨科机器人获FDA批准用于全髋置换,国产第二张FDA骨科机器人证- 来源:公司公告- 心脉医疗- 4月10日:可降解主动脉覆膜支架 III期:12个月通畅率97.1%(vs金属支架91.5%,P=0.018)- 来源:公司公告 (四)合成生物学 - Ginkgo Bioworks- 4月8日:与Moderna合作开发CGT载体,交付周期缩短65%,成本降50%- 来源:Ginkgo公告- 凯赛生物- 4月11日:生物基聚酰胺520产线投产,全球首发,用于医疗植入物- 来源:公司公告 四、重要获批与III期数据读出(本周) (一)国内获批 1. 恒瑞 SHR-A1811:4月6日,III期成功,申报NDA(HER2乳腺癌)2. 映恩生物 帕康曲妥珠单抗:4月9日,NMPA申报上市(HER2乳腺癌)3. 博睿康 无创脑电AI系统:4月11日,NMPA获批(癫痫定位) (二)海外获批/III期 1. 罗氏 Mosunetuzumab:4月8日,FDA获批(滤泡淋巴瘤)2. 诺和诺德 口服司美格鲁肽:4月9日,III期成功(减重)3. 卫材/渤健 Tilzapirumab:4月11日,III期成功(AD)4. Orca Bio Orca-T:4月6日,FDA延长审评(AML) 五、顶刊研究突破(本周发表) 1. Nature(4.8)浙大—工程化iCDCs治疗心衰,非人灵长类纤维化降65%,心功能升42%2. Nature(4.10)碱基编辑CS-101治疗β地贫,5例完全脱离输血,HbF升至38%3. Science(4.9)干细胞表观遗传—炎症记忆致患癌风险增2.3倍,新防癌靶点4. NEJM(4.9)优时比 Bimekizumab—银屑病关节炎结构损伤进展降62%5. Lancet(4.11)卫材 Tilzapirumab—早期AD认知衰退降28% 六、大额融资、并购、License(≥5亿/1亿美元,本周) (一)并购 1. 健适医疗→图格医疗:4月7日,控股收购,内窥镜影像,金额未披露2. 珠峰医药→Hasten Biopharma:4月8日,2.5亿美元,亚太8国慢病营销权3. 豪洛捷(Hologic):4月7日,黑石/TPG183亿美元私有化 (二)融资 1. 英矽智能:4月7日,3亿美元D轮,高瓴、淡马锡领投(AI制药)2. Neuralink:4月8日,2.8亿美元E轮,Founders Fund领投(脑机)3. 瑞迪奥医药:4月10日,1.2亿美元B轮,奥博、礼来亚洲(核药) (三)License 1. 荣昌生物→艾伯维:4月9日,PD-1/VEGF双抗(纬迪西妥单抗) 全球授权:首付8亿+里程碑20亿美元2. 劲方医药→罗氏:4月11日,KRAS G12C抑制剂海外授权:首付2.8亿+里程碑12亿美元 七、重点赛道竞争格局简评 1. ADC:国内HER2(恒瑞、映恩、科伦)、Nectin-4(百力司康)白热化;双抗ADC(百利天恒)成国际差异化核心2. GLP-1:口服化决战(礼来Orforglipron、诺和口服司美);双靶点/三靶点(礼来Mazdutide、信达IBI362)迭代提速3. CGT:碱基编辑(博雅辑因、Beam)临床验证;体内递送(LNP、AAV)成胜负手,国产效率追平国际4. AI制药:跨国巨头(礼来、罗氏)大额合作;靶点发现→临床验证周期缩短50%,商业化加速5. 脑机接口:侵入(Neuralink、博睿康)、无创双线推进;中国临床转化全球领先,政策+资本双轮驱动6. 合成生物学:医疗级材料、CGT载体成热点;AI+合成生物学提升效率,罗氏、Moderna加速布局 八、政策面关键变化 (一)国内 1. 医保:4月6日—AI辅助诊断纳入全国医保乙类,报销70%-85%,覆盖837家三甲2. 北京(4.7):创新药审评70→40天;国谈药15天挂网;政府引导基金100亿支持医药创新3. NMPA(4.10):《药物上市申请临床评价指导原则》发布,全球多中心数据直接认可 (二)海外 1. FDA(4.6):CGT加速通道扩围,罕见病/未满足需求临床60天审评2. 欧盟(4.9):CGT临床费用减免50%,专利延至15年
100 项与 南京图格医疗科技有限公司 相关的药物交易
登录后查看更多信息
100 项与 南京图格医疗科技有限公司 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2026年06月17日管线快照
无数据报导
登录后保持更新
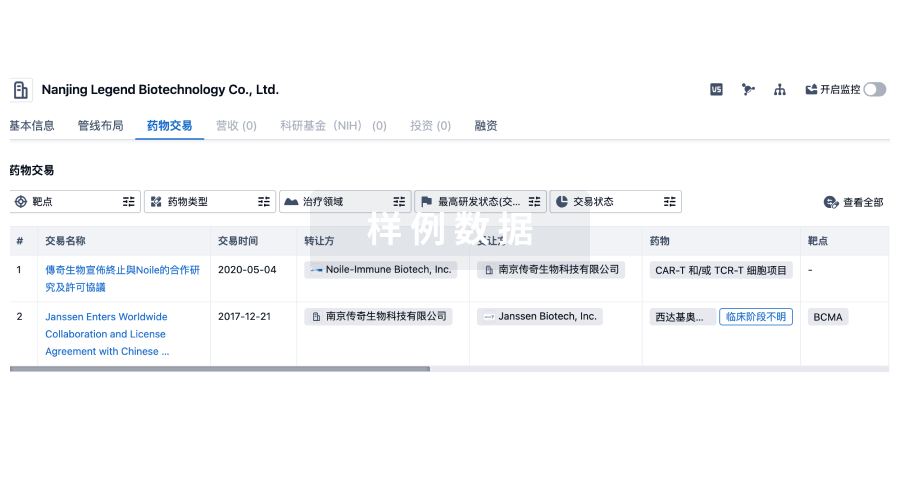
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
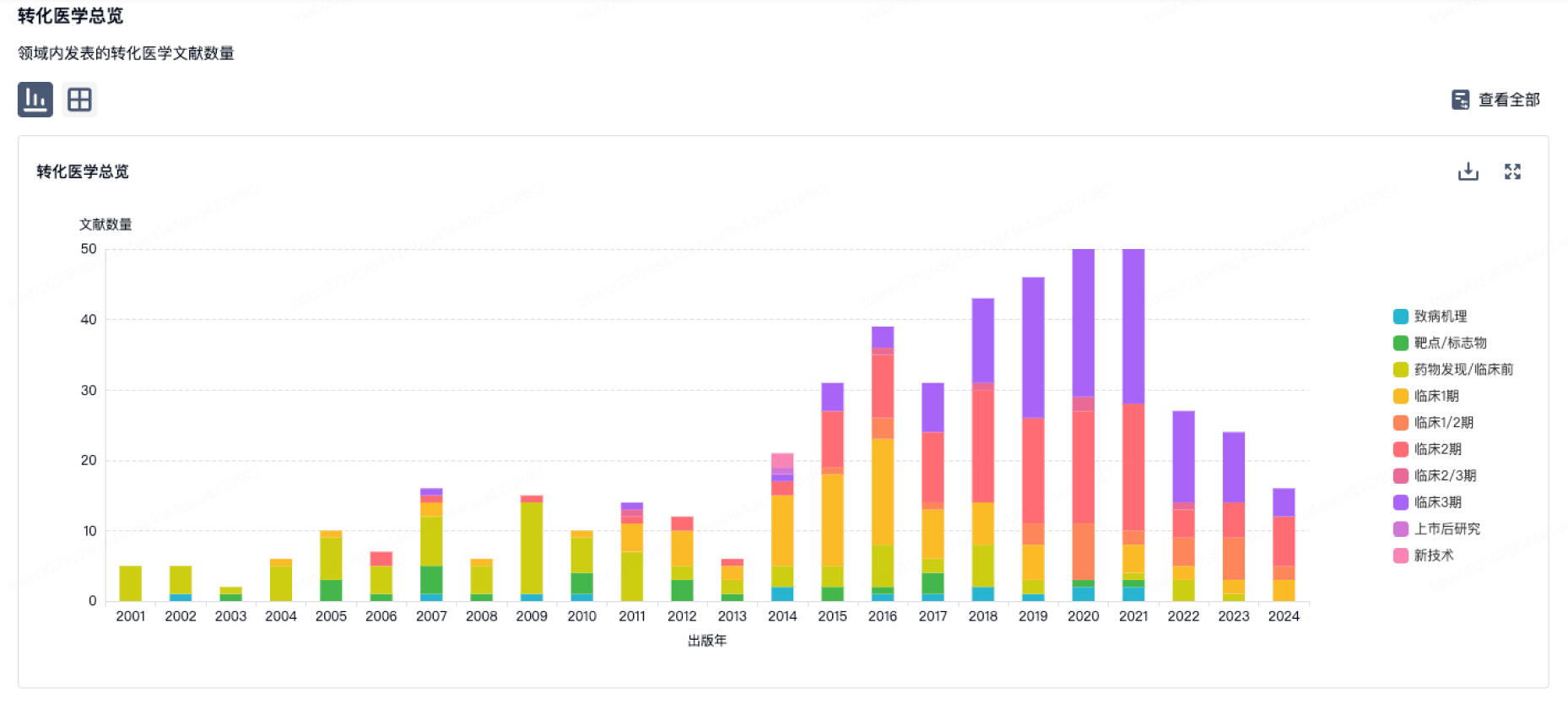
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
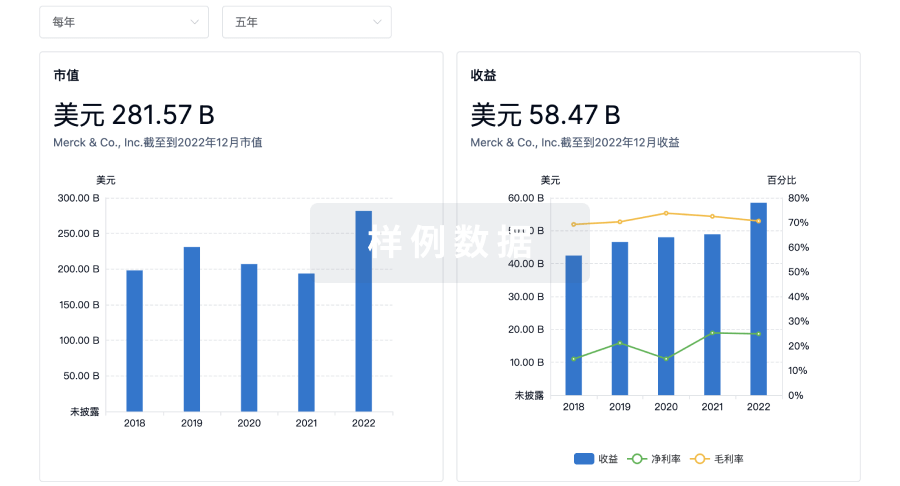
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
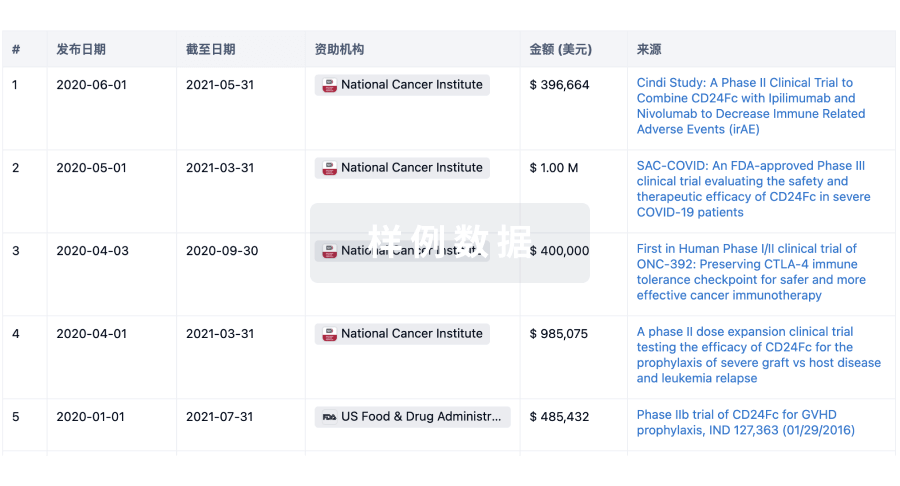
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
生物医药百科问答
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用