预约演示
更新于:2025-12-06
Wuhan ABclonal Biotechnology Co., Ltd.
更新于:2025-12-06
概览
关联
100 项与 武汉爱博泰克生物科技有限公司 相关的临床结果
登录后查看更多信息
0 项与 武汉爱博泰克生物科技有限公司 相关的专利(医药)
登录后查看更多信息
3
项与 武汉爱博泰克生物科技有限公司 相关的文献(医药)2023-08-01·Human vaccines & immunotherapeutics
Monovalent Omicron COVID-19 vaccine triggers superior neutralizing antibody responses against Omicron subvariants than Delta and Omicron bivalent vaccine
Article
作者: Zhao, Liwei ; Li, Wanting ; Zhao, Tiantian ; Wu, Chao ; Li, Chuang ; Ding, Xinyu ; Chen, Yuxin ; Xiao, Hang ; Cheng, Hao ; Tao, Bai ; Lou, Yang ; Chen, Lin
The continuous evolution of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) variants poses a challenge to determine the optimal updated composition of the coronavirus disease 2019 (COVID-19) vaccine. The present study aimed to investigate the immunogenicity of the Delta monovalent vaccine, the Omicron monovalent vaccine, and the Delta and Omicron BA.1 bivalent vaccine. Three COVID-19 vaccines were designed using the heterologous DNA prime-protein boost strategy, with each vaccine containing either Delta receptor-binding domain (RBD) of the spike protein, Omicron RBD, or both Delta and Omicron antigens. Temporal serum antibody binding titers and neutralizing antibody titers induced by the three vaccines in New Zealand White rabbits were analyzed. To further dissect the vaccine elicited antibodies (mAb) responses at the molecular level, a panel of rabbit monoclonal antibodies (RmAbs) was generated by a high-throughput single B cell sorting and discovery pipeline and further comprehensively characterized. The Omicron monovalent vaccine induced higher antibody binding titers and neutralization activities than the Delta and Omicron bivalent vaccine. Four RmAbs with robust neutralization capacity were isolated from rabbits immunized with the Omicron or Delta monovalent vaccine. Notably, 9E11 isolated from the Omicron monovalent vaccine group neutralized all the Omicron subvariants with an IC50 value ranging from 1.5 to 503.6 ng/mL; thus, this vaccine could serve as a prophylactic and therapeutic intervention. Given the increasing incidence of COVID-19 cases due to the Omicron variant, RBD from the Omicron strain could serve as a candidate immunogen that can induce higher neutralization activities against the SARS-CoV-2 Omicron sublineages.
2021-01-01·Emerging microbes & infections2区 · 医学
Potent RBD-specific neutralizing rabbit monoclonal antibodies recognize emerging SARS-CoV-2 variants elicited by DNA prime-protein boost vaccination
2区 · 医学
ArticleOA
作者: Huang, Weijin ; Chen, Jiachen ; Wu, Chao ; Tong, Xin ; Zhao, Xiangan ; Huang, Haibin ; Tong, Bei ; Chen, Yuxin ; Wu, Hai ; Zhu, Liguo ; Lou, Yang ; Tao, Yue
Global concerns arose as the emerged and rapidly spreading SARS-CoV-2 variants might escape host immunity induced by vaccination. In this study, a heterologous prime-boost immunization strategy for COVID-19 was designed to prime with a DNA vaccine encoding wild type (WT) spike protein receptor-binding domain (RBD) followed by S1 protein-based vaccine in rabbits. Four vaccine-elicited rabbit monoclonal antibodies (RmAbs), including 1H1, 9H1, 7G5, and 5E1, were isolated for biophysical property, neutralization potency and sequence analysis. All RmAbs recognized RBD or S1 protein with KD in the low nM or sub nM range. 1H1 and 9H1, but neither 7G5 nor 5E1, can bind to all RBD protein variants derived from B.1.351. All four RmAbs were able to neutralize wild type (WT) SARS-CoV-2 strain in pseudovirus assay, and 1H1 and 9H1 could neutralize the SARS-CoV-2 WT authentic virus with IC50 values of 0.136 and 0.026 μg/mL, respectively. Notably, 1H1 was able to neutralize all 6 emerging SARS-CoV-2 variants tested including D614G, B.1.1.7, B.1.429, P.1, B.1.526, and B.1.351 variants, and 5E1 could neutralize against the above 5 variants except P.1. Epitope binning analysis revealed that 9H1, 5E1 and 1H1 recognized distinct epitopes, while 9H1 and 7G5 may have overlapping but not identical epitope. In conclusion, DNA priming protein boost vaccination was an effective strategy to induce RmAbs with potent neutralization capability against not only SARS-CoV-2 WT strain but also emergent variants, which may provide a new avenue for effective therapeutics and point-of-care diagnostic measures.
2020-10-01·Monoclonal antibodies in immunodiagnosis and immunotherapy
Development of Rabbit Monoclonal Antibodies for Quantitation of Therapeutic Human Antibodies in Preclinical Non-Human Primate Studies
Article
作者: Gerstein, Jacob ; Chu, Ruiyin ; Huang, Haibin ; Lou, Yang ; Wu, Hai ; Palmer, Rachel
During preclinical studies, there is a great need to develop monoclonal antibodies (mAbs) that are specific to human immunoglobulin (IgG), without binding to monkey IgG, to detect therapeutic human mAb in non-human primates. We took advantage of the latest rabbit B cell cloning technology to develop six unique rabbit anti-human IgG mAb clones for this purpose. These clones are capable of binding to both human IgG and Fab with high affinity without nonspecific binding to cynomolgus monkey IgG. These clones have been evaluated as a generic capture reagent for the detection of human IgG and Fab, in the presence of cynomolgus monkey serum, by Gyrolab™ immunoassay. They may be used in singlet or as pairs for the detection of human IgG, in any host animal, to meet the need for therapeutic mAb development in preclinical studies.
354
项与 武汉爱博泰克生物科技有限公司 相关的新闻(医药)2025-11-30
·雪球
之前专栏有几篇体内Car-T的文章可以翻阅欢迎关注我,了解行业和公司信息蓬勃生物(ProBio)为金斯瑞旗下生物药CDMO业务板块本文相关个股:$金斯瑞生物科技(01548)$$东富龙(SZ300171)$$和元生物(SH688238)$以下信息来源:同写意当细胞疗法开始突破体外制备的传统范式,当“体内给药”与“CAR-T”这两个关键词在医药领域的交汇处激起涟漪——我们正在见证一场技术范式的革新。体内CAR-T,这一将CAR结构直接递送至患者体内、利用自身免疫细胞在体内生成CAR-T细胞的前沿技术,正以惊人的速度从实验室走向临床,其发展轨迹让人不禁回想起生物医药领域几次重大的技术革命。然而,这条创新之路并非坦途。从载体设计的精准性到靶向递送的特异性,从生产工艺的稳定性到临床应用的可靠性,体内CAR-T的发展面临着与传统体外CAR-T截然不同的挑战。这些挑战不仅关乎技术本身,更涉及整个产业生态的支撑能力——包括关键试剂、递送系统、生产工艺、质控标准等基础设施的完善程度。正如任何革命性技术的成熟都需要与之匹配的产业生态,体内CAR-T的进一步发展同样呼唤着更加完善、协同的创新基础设施。在此背景下,同写意作为中国新药研发领域的重要思想策源地,始终关注并推动着这一前沿领域的发展。在第16届泰州医博会期间,同写意专门设置体内CAR-T技术专场交流活动,汇聚了该领域的顶尖专家与创新企业,共同探讨技术突破与产业化路径。更为重要的是,现场,同写意联合优秀会员机构正式成立了“体内CAR-T新基建联盟”。该联盟的诞生,标志着中国体内CAR-T领域进入一个全新的发展阶段。联盟将致力于构建覆盖研发、生产、质控全链条的产业协同网络,推动关键试剂、材料、设备的国产化与标准化,建立符合体内CAR-T特点的技术规范与质量体系,助力中国企业在这场全球性的技术竞赛中把握先机。从体外到体内,从复杂到简洁,从高昂到可及,体内CAR-T技术正引领着细胞治疗走向新的时代。而“体内CAR-T新基建联盟”的建立,恰如为这场变革铺设了坚实的跑道,让中国的创新力量能够在这条新兴的赛道上加速奔跑,最终让更多患者受益于这一突破性技术的治疗价值。对于此次联盟成立,联盟单位分别从不同角度分享了各自的布局。东富龙东富龙凭借多年CGT细胞制药装备的制造底蕴和战略性深耕,在invivoCAR-T赛道已构建了领先的一体化布局:在LVV与mRNA的生产制备上积累了从质粒构建、细胞培养、分离纯化及递送的核心工艺与装备技术,多年来所提供的各类形式的上游细胞培养/微生物发酵、下游分离纯化到LNP递送相关实验室及GMP设备解决方案已助力了多家国内外知名CGT企业的申报及上市。东富龙是能够提供覆盖“上游培养/发酵与下游纯化、递送的全自动、一体化设备解决方案”的伙伴。从质粒制备、病毒生产、mRNA制备,到最终的LNP递送与制剂灌装,可实现全流程的自动化、封闭化生产,极大提升了生产效率,降低了污染风险与生产成本,为加速invivoCAR-T药物的研发与商业化提供了强有力的端到端支持。基锘威目前,体内CAR-T被视为肿瘤与自免疾病最具潜力的治疗策略之一,而任何治疗产品在临床前阶段的药效与安全性预测都至关重要。基锘威生物基于重度免疫缺陷小鼠BRGSF构建的PBMC或HSC人源免疫系统重建模型,可在模型体内不同程度重现人源免疫细胞的分布、功能及其与药物的相互作用,且模型状态稳定、实验窗口期长。该类模型能够支持体内/外CAR-T、TCR-T、NK等多类型细胞治疗产品的体内药效及安全性评价。我们希望以系统化、可预测的人源化模型体系,为体内细胞治疗的临床前研发提供可靠支撑,助力合作伙伴加速高质量创新疗法顺利迈向临床。益诺思益诺思在体内CAR-T等创新细胞基因治疗产品的非临床研究领域已建立起从早期研发至临床申报的全流程服务能力,涵盖成药性评估、模型构建、生物分布与安全性评价等关键环节,目前已承接多款体内CAR-T产品的非临床评价项目,公司始终聚焦前瞻性技术布局,持续提升对CGT产品的评价体系与监管认知,致力于推动此类划时代疗法加速完成申报并进入临床阶段。依科赛依科赛生物耕耘CAR-T领域多年,凭借AI赋能的高通量培养基筛选研发实验室与自动化的商业化生产基地,与国内多家CAR-T领军企业建立了长期战略合作伙伴关系。在invivoCAR-T领域,依科赛生物主要专注于LVV路线,提供慢病毒包装生产所必须的高品质、GMP级别的商业化293无血清培养基,产品均已在FDA进行DMF备案,还可以根据不同的研发和生产需求提供定制化服务,助力企业实现高滴度、批次间稳定性好的慢病毒生产,加速体内CAR-T项目在中国、美国等多个国家的申报进程。蓬勃生物在invivoCAR-T领域,蓬勃生物已形成了LVV与LNP双轨驱动下的产业化之路。在LVV领域,蓬勃生物拥有全面的慢病毒载体技术平台,提供从临床前、IND申报、临床试验阶段到商业化生产的一站式服务。其完善的质量体系已帮助客户完成80+包括NMPA、FDA、EMA、MFDS等全球多个地区的IND或BLA申请,为LVV路线企业提供了在全球合规和规模化生产上的强大支持。在LNP药物工艺方面,蓬勃生物依托其mRNACDMO和抗体蛋白药CDMO平台,可提供Ab-LNPs一站式工艺开发和GMP生产整体解决方案。这种整合能力大幅降低了客户在项目沟通、技术整合方面的难度,避免了因多方协作带来的进度延误和质量偏差,从而加速创新RNA药物的临床转化进程。妙顺生物妙顺(上海)生物科技有限公司成立于2014年,专注提供原代细胞整体解决方案,以新质生产力打破进口垄断。针对体内CAR-T研发核心痛点,公司构建全链条业务矩阵:高活性PBMC保障CAR基因稳定表达与T细胞体内扩增,精准分选的免疫细胞亚型筑牢疗法靶向性基础,合规单采血服务提供高浓度细胞原料,高质量疾病患者样本支撑临床相关研究新增NOG-DKO小鼠免疫重建预筛服务,进一步适配长效疗效与安全性验证需求。深研生物EuLV®是深研生物面向下一代基因细胞治疗及invivoCAR-T所打造的稳定细胞系慢病毒生产平台。依托自主开发的诱导型稳定生产细胞系(SPCL),配合高密度悬浮培养体系及CDM化学成分限定培养基,EuLV®实现了远超瞬转工艺的高滴度、高一致性与可规模化生产能力。通过分子开关、转座子整合及高通量自动化单克隆筛选,EuLV®能稳定输出适用于invivo递送需求的高性能LVV,滴度提升可达20倍以上,且具备更高的特异活性与更佳的PBMC/初始T细胞转导表现。作为解决病毒载体生产瓶颈的核心技术,EuLV®将帮助客户加速invivoCAR-T与基因细胞治疗产品的临床转化与产业化推进。行诚生物杭州行诚生物科技有限公司专注于病毒载体和非病毒载体领域,为全球药企提供高效的CRO和CDMO解决方案。从2022年创立至今,我们致力于革新病毒载体全生产流程,核心优势在于:质粒生产采用独创的“无层析工艺”,通过线性流纯化技术,大幅缩短生产周期,降低生产成本,显著提高回收率与质量,完美契合快速、高产的临床与商业化需求。慢病毒上游环节,我们拥有成熟的高滴度悬浮培养及多质粒瞬转技术平台,确保稳定产出高感染活性的病毒。下游环节采用全封闭,一次性工艺,提升病毒质量与安全,为体内CAR-T的高效递送提供坚实保障。行诚生物以前沿工艺,与合作伙伴一起加速下一代基因药物的开发与商业化。聚茵生物聚茵生物拥有全球首创的独家超支化聚β氨基酯HPAE基因递送技术平台,提供超高效可降解的产毒转染试剂BrPERfect®和精准体内基因递送聚合物平台。和元生物在体内CAR-T这一颠覆性技术领域,和元生物依托深厚的载体工艺积累与前瞻性技术布局,已构建起以精准递送载体为核心,病毒与非病毒双线战略并行的多元化技术平台,为全球药企提供覆盖从靶点验证、工艺开发到规模化生产的全周期CDMO解决方案。在病毒载体路径上,和元生物凭借国内领先的病毒载体平台,提供包括衣壳优化、靶向性提升、工艺放大与GMP生产在内的一站式服务。我们成熟的质量体系与丰富的申报经验,已成功支持多个病毒载体项目完成CMC研究及中美双报,为客户扫清了从实验室到临床的产业化障碍。在非病毒路径上,我们通过战略合作积极布局LNP递送技术,可提供从mRNA序列优化、靶向LNP筛选到GMP生产的整合式开发方案。病毒与非病毒载体双轨并行策略,为体内CAR-T的快速迭代与临床转化提供了高效、可靠的解决方案。和元生物期待以坚实的平台能力,赋能创新构想,共同加速下一代细胞药物的上市进程。途深智合途深智合由上海交通大学王宇光副教授创立,核心团队成员来自上海交通大学、约翰霍普金斯大学、剑桥大学、伦敦大学学院等顶尖高校及产业界,具备深厚的AI蛋白设计与生物研发经验。公司在生物药方向积累了系列模型与丰富的产品实践。很荣幸加入Car-T新基建联盟,希望途深的AI能力能够加速Car-T细胞的研发设计和实验验证。爱博泰克爱博泰克依托成熟的抗体开发技术平台,在体内CAR-T开发中聚焦早期发现与质控评价两大核心环节。依托单B细胞平台,我们已构建成熟且可追溯的高亲和力流式抗体序列库,可基于既有序列快速完成scFv工程化改造,并提供序列授权服务,显著缩短从靶点发现到药效验证的研发周期。在功能验证与质量控制方面,爱博泰克隆重推出“无爱博,不流式”全方位流式解决方案。该方案囊括流式分析与检测所需的试剂及仪器,涵盖细胞流式与微球流式两大方向,全面支持T细胞活化、扩增、耗竭以及细胞因子释放等关键指标的检测,为体内CAR功能状态与安全性研究提供系统、可靠的技术保障。恒驭生物恒驭生物以国际合规标准为依托,凭借卓越的质量和技术实力,为invivoCAR-T等创新药企业提供涵盖细胞库检定(CLC)、复制型病毒检测(RCV)等生物安全检测服务,满足NMPA、FDA、EMA等全球监管机构的严格要求,为创新药企提供可靠的保障。恒驭生物已累计服务300多家生物药企,项目及申报实战经验丰富,已经为110+生物药的申报案例及30+BLA和商业化项目提供关键性支持(其中50+海外申报案例),致力于加速创新疗法在全球范围内的临床转化与商业化进程,是药企在创新药上市出海过程中值得信赖的合作伙伴。
细胞疗法免疫疗法信使RNA
2025-11-28
·同写意
当细胞疗法开始突破体外制备的传统范式,当“体内给药”与“CAR-T”这两个关键词在医药领域的交汇处激起涟漪——我们正在见证一场技术范式的革新。
体内CAR-T,这一将CAR结构直接递送至患者体内、利用自身免疫细胞在体内生成CAR-T细胞的前沿技术,正以惊人的速度从实验室走向临床,其发展轨迹让人不禁回想起生物医药领域几次重大的技术革命。
然而,这条创新之路并非坦途。从载体设计的精准性到靶向递送的特异性,从生产工艺的稳定性到临床应用的可靠性,体内CAR-T的发展面临着与传统体外CAR-T截然不同的挑战。
这些挑战不仅关乎技术本身,更涉及整个产业生态的支撑能力——包括关键试剂、递送系统、生产工艺、质控标准等基础设施的完善程度。正如任何革命性技术的成熟都需要与之匹配的产业生态,体内CAR-T的进一步发展同样呼唤着更加完善、协同的创新基础设施。
在此背景下,同写意作为中国新药研发领域的重要思想策源地,始终关注并推动着这一前沿领域的发展。在第16届泰州医博会期间,同写意专门设置体内CAR-T技术专场交流活动,汇聚了该领域的顶尖专家与创新企业,共同探讨技术突破与产业化路径。
更为重要的是,现场,同写意联合优秀会员机构正式成立了“体内CAR-T新基建联盟”。
同写意程博士为联盟机构代表颁发证书
该联盟的诞生,标志着中国体内CAR-T领域进入一个全新的发展阶段。联盟将致力于构建覆盖研发、生产、质控全链条的产业协同网络,推动关键试剂、材料、设备的国产化与标准化,建立符合体内CAR-T特点的技术规范与质量体系,助力中国企业在这场全球性的技术竞赛中把握先机。
从体外到体内,从复杂到简洁,从高昂到可及,体内CAR-T技术正引领着细胞治疗走向新的时代。而“体内CAR-T新基建联盟”的建立,恰如为这场变革铺设了坚实的跑道,让中国的创新力量能够在这条新兴的赛道上加速奔跑,最终让更多患者受益于这一突破性技术的治疗价值。
对于此次联盟成立,联盟单位分别从不同角度分享了各自的布局。
东富龙
东富龙凭借多年CGT细胞制药装备的制造底蕴和战略性深耕,在in vivo CAR-T赛道已构建了领先的一体化布局:在LVV与mRNA的生产制备上积累了从质粒构建、细胞培养、分离纯化及递送的核心工艺与装备技术,多年来所提供的各类形式的上游细胞培养/微生物发酵、下游分离纯化到LNP递送相关实验室及GMP设备解决方案已助力了多家国内外知名CGT企业的申报及上市。
东富龙是能够提供覆盖“上游培养/发酵与下游纯化、递送的全自动、一体化设备解决方案”的伙伴。从质粒制备、病毒生产、mRNA制备,到最终的LNP递送与制剂灌装,可实现全流程的自动化、封闭化生产,极大提升了生产效率,降低了污染风险与生产成本,为加速invivoCAR-T药物的研发与商业化提供了强有力的端到端支持。
基锘威
目前,体内CAR-T被视为肿瘤与自免疾病最具潜力的治疗策略之一,而任何治疗产品在临床前阶段的药效与安全性预测都至关重要。基锘威生物基于重度免疫缺陷小鼠BRGSF构建的PBMC或HSC人源免疫系统重建模型,可在模型体内不同程度重现人源免疫细胞的分布、功能及其与药物的相互作用,且模型状态稳定、实验窗口期长。该类模型能够支持体内/外CAR-T、TCR-T、NK等多类型细胞治疗产品的体内药效及安全性评价。我们希望以系统化、可预测的人源化模型体系,为体内细胞治疗的临床前研发提供可靠支撑,助力合作伙伴加速高质量创新疗法顺利迈向临床。
益诺思
益诺思在体内CAR-T等创新细胞基因治疗产品的非临床研究领域已建立起从早期研发至临床申报的全流程服务能力,涵盖成药性评估、模型构建、生物分布与安全性评价等关键环节,目前已承接多款体内CAR-T产品的非临床评价项目,公司始终聚焦前瞻性技术布局,持续提升对CGT产品的评价体系与监管认知,致力于推动此类划时代疗法加速完成申报并进入临床阶段。
依科赛
依科赛生物耕耘CAR-T领域多年,凭借AI赋能的高通量培养基筛选研发实验室与自动化的商业化生产基地,与国内多家CAR-T领军企业建立了长期战略合作伙伴关系。在in vivo CAR-T领域,依科赛生物主要专注于LVV路线,提供慢病毒包装生产所必须的高品质、GMP级别的商业化293无血清培养基,产品均已在FDA进行DMF备案,还可以根据不同的研发和生产需求提供定制化服务,助力企业实现高滴度、批次间稳定性好的慢病毒生产,加速体内CAR-T项目在中国、美国等多个国家的申报进程。
蓬勃生物
在in vivo CAR-T领域,蓬勃生物已形成了LVV与LNP双轨驱动下的产业化之路。在LVV领域,蓬勃生物拥有全面的慢病毒载体技术平台,提供从临床前、IND申报、临床试验阶段到商业化生产的一站式服务。其完善的质量体系已帮助客户完成80+包括NMPA、FDA、EMA、MFDS等全球多个地区的IND或BLA申请,为LVV路线企业提供了在全球合规和规模化生产上的强大支持。
在LNP药物工艺方面,蓬勃生物依托其mRNA CDMO和抗体蛋白药CDMO平台,可提供Ab-LNPs一站式工艺开发和GMP生产整体解决方案。
这种整合能力大幅降低了客户在项目沟通、技术整合方面的难度,避免了因多方协作带来的进度延误和质量偏差,从而加速创新RNA药物的临床转化进程。
妙顺生物
妙顺(上海)生物科技有限公司成立于2014年,专注提供原代细胞整体解决方案,以新质生产力打破进口垄断。针对体内CAR-T研发核心痛点,公司构建全链条业务矩阵:高活性PBMC保障CAR基因稳定表达与T细胞体内扩增,精准分选的免疫细胞亚型筑牢疗法靶向性基础,合规单采血服务提供高浓度细胞原料,高质量疾病患者样本支撑临床相关研究新增NOG-DKO小鼠免疫重建预筛服务,进一步适配长效疗效与安全性验证需求。
深研生物
EuLV®是深研生物面向下一代基因细胞治疗及in vivo CAR-T所打造的稳定细胞系慢病毒生产平台。依托自主开发的诱导型稳定生产细胞系(SPCL),配合高密度悬浮培养体系及CDM化学成分限定培养基,EuLV®实现了远超瞬转工艺的高滴度、高一致性与可规模化生产能力。通过分子开关、转座子整合及高通量自动化单克隆筛选,EuLV®能稳定输出适用于in vivo递送需求的高性能LVV,滴度提升可达20倍以上,且具备更高的特异活性与更佳的PBMC/初始T细胞转导表现。作为解决病毒载体生产瓶颈的核心技术,EuLV®将帮助客户加速in vivo CAR-T与基因细胞治疗产品的临床转化与产业化推进。
行诚生物
杭州行诚生物科技有限公司专注于病毒载体和非病毒载体领域,为全球药企提供高效的CRO和CDMO解决方案。
从2022年创立至今,我们致力于革新病毒载体全生产流程,核心优势在于:质粒生产采用独创的“无层析工艺”,通过线性流纯化技术,大幅缩短生产周期,降低生产成本,显著提高回收率与质量,完美契合快速、高产的临床与商业化需求。慢病毒上游环节,我们拥有成熟的高滴度悬浮培养及多质粒瞬转技术平台,确保稳定产出高感染活性的病毒。下游环节采用全封闭,一次性工艺,提升病毒质量与安全,为体内CAR-T的高效递送提供坚实保障。
行诚生物以前沿工艺,与合作伙伴一起加速下一代基因药物的开发与商业化。
聚茵生物
我们拥有全球独家的HPAE基因递送技术平台,提供超高效可降解的产毒转染试剂和精准体内基因递送聚合物平台。
和元生物
在体内CAR-T这一颠覆性技术领域,和元生物依托深厚的载体工艺积累与前瞻性技术布局,已构建起以精准递送载体为核心,病毒与非病毒双线战略并行的多元化技术平台,为全球药企提供覆盖从靶点验证、工艺开发到规模化生产的全周期CDMO解决方案。
在病毒载体路径上,和元生物凭借国内领先的病毒载体平台,提供包括衣壳优化、靶向性提升、工艺放大与GMP生产在内的一站式服务。我们成熟的质量体系与丰富的申报经验,已成功支持多个病毒载体项目完成CMC研究及中美双报,为客户扫清了从实验室到临床的产业化障碍。
在非病毒路径上,我们通过战略合作积极布局LNP递送技术,可提供从mRNA序列优化、靶向LNP筛选到GMP生产的整合式开发方案。
病毒与非病毒载体双轨并行策略,为体内CAR-T的快速迭代与临床转化提供了高效、可靠的解决方案。和元生物期待以坚实的平台能力,赋能创新构想,共同加速下一代细胞药物的上市进程。
途深智合
途深智合由上海交通大学王宇光副教授创立,核心团队成员来自上海交通大学、约翰霍普金斯大学、剑桥大学、伦敦大学学院等顶尖高校及产业界,具备深厚的AI蛋白设计与生物研发经验。公司在生物药方向积累了系列模型与丰富的产品实践。很荣幸加入Car-T新基建联盟,希望途深的AI能力能够加速Car-T细胞的研发设计和实验验证。
爱博泰克
爱博泰克依托成熟的抗体开发技术平台,在体内CAR-T开发中聚焦早期发现与质控评价两大核心环节。依托单B细胞平台,我们已构建成熟且可追溯的高亲和力流式抗体序列库,可基于既有序列快速完成scFv工程化改造,并提供序列授权服务,显著缩短从靶点发现到药效验证的研发周期。在功能验证与质量控制方面,爱博泰克隆重推出“无爱博,不流式”全方位流式解决方案。该方案囊括流式分析与检测所需的试剂及仪器,涵盖细胞流式与微球流式两大方向,全面支持T细胞活化、扩增、耗竭以及细胞因子释放等关键指标的检测,为体内CAR功能状态与安全性研究提供系统、可靠的技术保障。
恒驭生物
恒驭生物以国际合规标准为依托,凭借卓越的质量和技术实力,为in vivo CAR-T等创新药企业提供涵盖细胞库检定(CLC)、复制型病毒检测(RCV)等生物安全检测服务,满足NMPA、FDA、EMA等全球监管机构的严格要求,为创新药企提供可靠的保障。恒驭生物已累计服务300多家生物药企,项目及申报实战经验丰富,已经为110+生物药的申报案例及30+BLA和商业化项目提供关键性支持(其中50+海外申报案例),致力于加速创新疗法在全球范围内的临床转化与商业化进程,是药企在创新药上市出海过程中值得信赖的合作伙伴。
细胞疗法免疫疗法信使RNA
2025-11-19
宫颈癌是常见的妇科恶性肿瘤[1],全球女性癌症死亡的主要原因之一。在发展中国家,宫颈癌新增病例为45.2万例,排名第二[2],其病理类型分为鳞癌、腺癌、鳞腺癌,持续感染高危人乳头瘤病毒(HPV)是最主要的原因。宫颈癌的远处转移扩散与长期生存率低有关,其中原位癌高发年龄为30~50岁,浸润癌为45~55岁[3],近年来发病有年轻化趋势。手术仍然是早期宫颈癌患者的主要治疗方法,局部晚期肿瘤采用化疗[4]。
白桦脂酸(Betulinic acid,BA)是存在于蒲桃树叶、白桦树皮、酸枣仁等植物中的天然羽扇豆烷型五环三萜化合物,具有抗肿瘤、抗HIV、调血脂、抗糖尿病等作用[5]。BA通过体内与体外机制抑制癌细胞的生长,并靶向mTOR信号来抑制胰腺癌的凋亡[6]。BA通过抑制炎症与血管的生成,导致细胞周期停滞,最终导致三阴性乳腺癌细胞凋亡[7]。BA可以通过线粒体依赖性和非依赖性途径诱导卵巢癌细胞的凋亡[8]。本文探索BA对宫颈癌细胞的抑制作用。1 材料与方法1.1 实验材料
人宫颈癌细胞株SIHA购自斑马生物技术公司,RPMI1640培养基、胎牛血清购自佰欧公司,双抗为碧云天生物技术公司生产,96孔板为Corning公司产品。BA购自上海源叶生物公司,MTT、二甲基亚砜(DMSO)为SIGMA公司产品。GAPDH Rabbit Monoclonal Antibody(AF0006)购自Beyotime生物公司,c-Myc单克隆抗体购自Abclonal公司。酶标仪(天逸510S-07ICB)购自美国 Bio-Tek公司,BCA蛋白定量检测试剂盒、蛋白提取试剂盒和 SDS-PAGE 凝胶试剂盒均购自白鲨生物。电泳仪(EPS 600)购自雅酶公司,倒置显微镜(TH4-200)购自奥斯维公司。其他试剂为生化分析纯。
A.BA作用于SIHA细胞24h后的细胞白光图(×100);B.一定浓度梯度的BA与SIHA细胞孵育24h后,BA对SIHA细胞增殖的影响(与空白对照组比较,*P<0.05,**P<0.01,***P<0.001,****P<0.0001,n=3);C.与10μmol/L BA孵育24、48、72h后,SIHA细胞的增殖能力被时间依赖性抑制(与24h组比较,**P<0.01,****P<0.0001,n=3)。
图1 BA对SIHA细胞增殖的影响
A.不同浓度的BA对SIHA细胞的克隆形成图;B.不同浓度BA对SIHA细胞的克隆数统计图(与空白对照组比较,*P<0.05,**P<0.01,****P<0.0001,n=3)
图2 BA对SIHA细胞克隆形成能力的影响
A.不同浓度的BA对SIHA细胞纵向迁移的抑制情况(×20);B.BA对SIHA细胞纵向迁移能力抑制的统计图(与空白对照组比较,*P<0.05,****P<0.0001,n=3)
图3 BA抑制SIHA细胞纵向迁移
A.不同浓度的BA对SIHA细胞侵袭能力的影响(×20);B.BA对SIHA细胞侵袭抑制的统计图(与空白对照组比较,**P<0.01,****P<0.0001,n=3)
图4 BA抑制SIHA细胞侵袭
A.Western blot结果图;B.Western blot结果统计图(与空白对照组相比,**P<0.01,****P<0.0001,n=3)。
图5 BA对SIHA细胞c-Myc蛋白表达的影响
A.c-Myc-saRNA可以在SIHA细胞中上调c-Myc的蛋白表达水平;B.c-Myc-saRNA可以部分抵消BA对宫颈癌SIHA细胞的抑制作用(与对照组相比,*P<0.05,**P<0.01,***P<0.001,**** P<0.0001,n=3)。
图6 c-Myc-saRNA BA对宫颈癌SIHA细胞的作用1.2 方法
1.2.1 细胞培养
使用RPMI-1640完全培养基(10% FBS、90%RPMI-1640、1%青霉素链霉素双抗)在37℃、5% CO2细胞培养箱内培养宫颈癌SIHA细胞。
1.2.2 MTT法
待SIHA细胞长至对数增长期时,消化传代后,以每孔5×105接种于96孔板中。接下来的实验步骤参照以前发表的文献[9],BA的浓度设定为0、5、10、20、40、80μmol/L。
1.2.3 平板克隆形成实验
实验步骤参照以前发表的文献[9],BA的浓度设定为0、5、10、40μmol/L。
1.2.4 Transwell小室侵袭和迁移实验
具体实验步骤参照以前发表的文献[9],BA的浓度设定为0、5、10、40μmol/L。
1.2.5 蛋白检测
用不同浓度梯度(0、5、10、40μmol/L)BA与SIHA细胞孵育24h后,提取细胞总蛋白,接下来的实验步骤参考以前发表的文献[10]。
1.2.6 saRNA激活c-Myc蛋白的表达
根据文献报道的方法[11],设计针对人c-Myc基因启动子区的三对小激活RNA(small activating RNA,saRNA),saRNA的序列见表1。以Lipofectamine2000为转染试剂,采用瞬时转染的方法转进宫颈癌SIHA细胞。Western blot检测转染效果。
表1 saRNA序列
1.3 统计学方法
采用GraphPad Prism 9.0进行统计分析及绘图,数据结果以表示,并用One-Way ANOVA进行分析,P<0.05表示差异具有统计学意义。2 结 果2.1 BA对宫颈癌SIHA细胞存活和增殖能力的影响
通过MTT实验检测BA对SIHA细胞存活和增殖能力的抑制情况,如图1所示,不同浓度BA(0、5、10、20、40、80μmol/L)作用24h后,SIHA细胞的活力逐渐被抑制,普通光学显微镜下观察表现为细胞逐渐失去典型多角形,变圆、漂浮,光密度值(OD)逐渐减少,说明其抑制程度随BA浓度的增加而增强,呈现剂量依赖性,经计算,IC50为28.52μmol/L。为了评估时间依赖性,本文选取10μmol/L的浓度来测算。随着时间的推移,10μmol/L BA呈时间依赖性抑制人宫颈癌细胞株SIHA的增殖,表现为OD值逐渐减少。2.2 BA对宫颈癌SIHA细胞克隆形成能力的影响
平板克隆形成实验反映了单个癌细胞的增殖和集落形成能力。如图2所示,一定浓度梯度的BA(0、5、10、40μmol/L)孵育后,BA浓度依赖性抑制了宫颈癌SIHA细胞的克隆形成能力,表现为肉眼可见的克隆逐步减少。2.3 BA对宫颈癌SIHA细胞的纵向迁移能力的影响
Transwell小室迁移实验反映了癌细胞的纵向迁移能力。如图3所示,与一定浓度梯度的BA(0、5、10、40μmol/L)孵育后,BA浓度依赖性抑制了宫颈癌细胞的迁移,表现为穿过基底膜的细胞逐渐减少。2.4 BA对宫颈癌SIHA细胞侵袭的影响
Transwell小室侵袭实验反映了癌细胞的侵袭能力。如图4所示,一定浓度梯度的BA(0、5、10、40μmol/L)与SIHA细胞孵育后,BA浓度依赖性抑制了宫颈癌SIHA细胞的侵袭能力,表现为穿过基底胶和基底膜的细胞逐渐减少。2.5 BA对宫颈癌SIHA细胞c-Myc蛋白表达的影响
如图5所示,从经BA孵育后的宫颈癌细胞中提取总蛋白,实施Western blot检查,发现BA剂量依赖性(0、5、10、40μmol/L)下调了宫颈癌SIHA细胞c-Myc蛋白的表达,说明BA可能通过下调c-Myc基因的翻译抑制宫颈癌SIHA细胞的增殖和迁移。2.6 c-Myc表达的抑制作用
转染c-Myc-saRNA后,发现SIHA细胞的c-Myc蛋白表达显著上调,其中c-Myc-saRNA-1激活效果最明显(图6A),利用20μmol/L BA与SIHA细胞共孵育,发现c-Myc-saRNA-1可以部分抵消BA对SIHA细胞的抑制作用(图6B),说明BA对宫颈癌SIHA细胞的抑制作用部分通过下调c-Myc蛋白的表达而实现。3 讨 论
宫颈癌是全球女性常见的癌症之一,也是导致癌症相关死亡率的原因。几乎所有的宫颈癌都与人类乳头瘤样病毒(HPV)感染有相关性[12]。包括宫颈鳞状细胞癌(70%)、宫颈腺癌(25%)和混合组织学肿瘤。虽然能在普查中及时确诊,但后期的生存率却很低,仅为一年半时间[13]。
BA是一种五环三萜类化合物,在天然界中广泛存在,最早分离于生长在非洲东部的鼠李科常绿植物的树皮,但主要来源是从白桦树皮中提取纯化[14]。BA及其衍生物作为生物制剂在抗癌和HIV治疗等方面表现出了巨大的潜能,在机体的抗氧化损伤和免疫调节过程中发挥着重要作用,是一类很有潜力的药物先导化合物,对于新药的研发具有重要价值[15]。BA在许多癌细胞中具有抗增殖特性,如胰腺癌、胃癌、乳腺癌、肾癌、卵巢癌等。Guo等[16]通过体外和体内实验证明BA均以剂量依赖性方式明显抑制胰腺癌,并且通过特异性靶向mTOR信号而不是Nrf2或JAK2来抑制胰腺癌。Atas等[17]通过评估EMT标志物SNAIL-1和SDC-2来研究BA对肾细胞癌细胞的抗转移作用,通过细胞毒性测定剂量处理BA,SDC-2的表达水平降低,并且抑制癌细胞的增殖与迁移。
许多原癌基因已被证明参与调节细胞凋亡和癌症发生。Myc原癌基因编码一个转录因子家族,是人类癌症中常见的激活癌基因之一。在正常生长的成人组织中,c-Myc表达与生长有关,当静止的组织被刺激生长时,如在再生肝中,c-Myc表达增加,当扩增时表明预后不良。细胞生长的停止通常伴随着c-Myc表达的大幅降低。当使用反义寡核苷酸来降低c-Myc的表达水平,即使不会导致死亡,也会导致生长抑制。它在癌症中的作用是普遍存在的,因为它促进生长、细胞周期进程、代谢和生存[18]。c-Myc涉及多种细胞过程,包括增殖、分化、细胞凋亡和代谢[19],长期培养的成纤维细胞中加入生长因子,诱导c-Myc基因表达,可刺激细胞增殖;当撤去生长因子,且下调c-Myc基因表达时,细胞停滞在G1期而不增殖。c-Myc与正调节的生长因子协同促进细胞增殖,而与具有负调控的生长抑制基因Fas、Fasl和Bas结合后可加速细胞凋亡。成骨肉瘤、软骨肉瘤、脊索瘤、脂肪肉瘤、横纹肌肉瘤、何杰金氏病及头部肿瘤等都有Myc基因的扩增或过度表达[20]。
尽管越来越多的研究提示[21-22]原癌基因c-Myc卷入宫颈癌的发生和发展,但是c-Myc的突变更多地是被作为宫颈癌预后不良的生物标志物,而不是起决定作用的驱动因素[23]。Riou等[24]发现在93例早期宫颈癌中c-Myc基因的过表达预示着更高的复发风险,c-Myc基因的检测联合淋巴结转移状况分析能更精确地预测复发风险。Ocadiz等[25]发现在35例墨西哥妇女宫颈癌样本中约90%存在c-Myc基因的扩增和/或重排。但是迄今为止,似乎未见c-Myc基因在宫颈癌中的功能学研究。本研究发现BA可以剂量依赖性下调宫颈癌SIHA细胞的恶性生物学行为和c-Myc蛋白的表达,且saRNA上调c-Myc基因的表达后可以部分抵消BA的抑制作用,间接地证实了c-Myc基因在宫颈癌中的致瘤性功能。
总之,本研究通过体外实验证明BA能有效抑制宫颈癌细胞的增殖和迁移,且通过下调c-Myc蛋白的表达得以实现。后续将进行裸鼠体内实验验证BA是否在体内具有相同的作用,为临床应用提供理论依据。
参考文献(略)
来源:祝巧云,朱波.白桦脂酸通过下调c-Myc抑制宫颈癌SIHA细胞的增殖与迁移[J].湖北科技学院学报(医学版),2025,39(5):386-390.
申请上市诊断试剂临床结果
100 项与 武汉爱博泰克生物科技有限公司 相关的药物交易
登录后查看更多信息
100 项与 武汉爱博泰克生物科技有限公司 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2026年06月03日管线快照
无数据报导
登录后保持更新
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
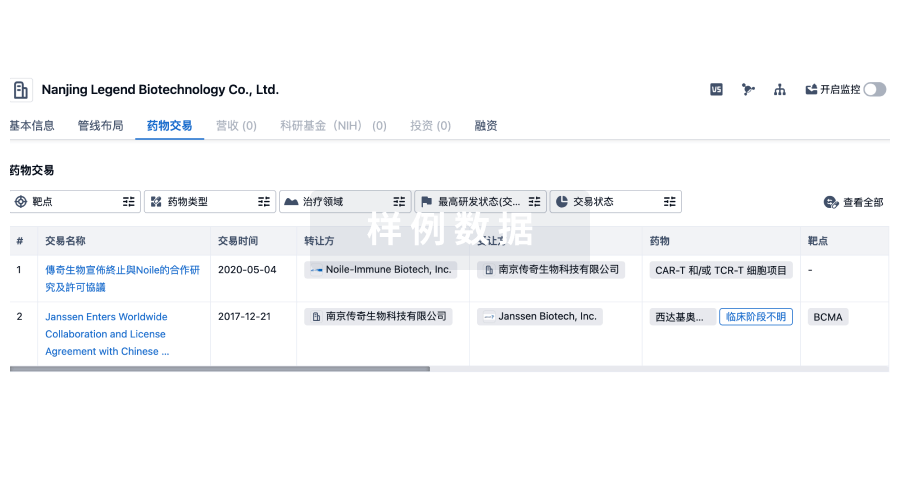
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
生物医药百科问答
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用