预约演示
更新于:2025-08-11

Marcus Research Laboratory, Inc.
更新于:2025-08-11
概览
关联
100 项与 Marcus Research Laboratory, Inc. 相关的临床结果
登录后查看更多信息
0 项与 Marcus Research Laboratory, Inc. 相关的专利(医药)
登录后查看更多信息
7
项与 Marcus Research Laboratory, Inc. 相关的新闻(医药)2025-07-29
·药研网
2025年7月29日,跨国药企诺和诺德宣布,Maziar Mike Doustdar将于2025年8月7日起出任公司总裁兼首席执行官。Maziar Mike Doustdar将接替同日卸任的Lars Fruergaard Jørgensen,同时还公布了将于8月7日生效的其他高管变动的一系列消息。回顾Lars Fruergaard Jørgensen的任期,自1991年加入公司以来,他在多个关键岗位历练,并于2017年1月起出任CEO。在他领导下,诺和诺德不仅巩固了糖尿病治疗领域的领先地位,还在肥胖症治疗上成功开拓市场,并逐步扩展至其他严重慢性疾病的治疗领域。Lars Fruergaard Jørgensen/图自路透社此次人事调整发生在诺和诺德面临多重市场挑战的背景下。尽管公司在GLP-1类药物(如Ozempic和Wegovy)推动下实现了显著增长,销售额、利润和市值在过去八年中几乎翻倍,但自2024年中期以来,股价持续走弱。在激烈的市场竞争下,礼来在GLP-1领域强势追赶,蚕食了诺和诺德的市场份额,而诺和诺德自己的新药管线也没能打动投资者。今年第一季度销售额达781亿丹麦克朗(约合113亿美元),按丹麦克朗计算增长19%,按不变汇率(CER)计算增长18%。不及预期的787亿丹麦克朗。同时诺和诺德还下调了全年销售额预测,原先预计销售额增长16%到24%,现在调整为13%到21%。将运营利润预期从之前的19%到27%下调至16%到24%。也令市场对其增长能力产生担忧。在此背景下,诺和诺德基金会董事会与公司董事会启动了有关加快CEO更替的对话。诺和诺德此前公告称,考虑到市场挑战、股价下跌等原因,寻找新的CEO最符合公司和股东利益。即将上任CEO的Mike Doustdar现任诺和诺德国际运营执行副总裁,拥有卓越的业绩记录。在过去十年里,其领导的国际运营业务销售额增长逾一倍,2024年达到约1,120亿丹麦克朗。该业务涵盖美国以外所有诺和诺德子公司,员工近2万人。此前,Mike Doustdar常驻马来西亚领导东南亚及大洋洲商业区域,并曾负责中东业务。在这之前,他担任土耳其总经理,并在财务与IT领域担任领导职务。Mike Doustdar于2015年升任执行副总裁,同时担任Orion Corporation董事会成员。诺和诺德将于8月7日生效的其他组织变动还有:诺和诺德决定将公司的“研发与早期开发”与“开发”两大职能合并为一个新的、统一的研发部门,由 Martin Holst Lange博士领导。现任开发执行副总裁 Martin Holst Lange 将于8月7日起被任命为首席科学官(CSO)。Martin Holst Lange将专注于无缝整合两个职能,快速推进新疗法创新,确保早期及后期管线的成功,并重点关注糖尿病与肥胖领域。此外,他还将与Mike Doustdar 紧密合作,推动公司内外部的管线开发与创新。现任研发与早期开发执行副总裁兼首席科学官Marcus Schindler 已决定退休。董事会主席 Helge Lund 表示:“我们感谢 Marcus 对诺和诺德及科学卓越追求的深度投入,并祝愿他未来一切顺利。”现任欧洲及加拿大区域高级副总裁 Emil Kongshøj Larsen 将加入执行管理团队,接替Mike Doustdar,担任国际运营执行副总裁。Emil Larsen 目前领导覆盖40个国家的区域,占诺和诺德全球销售额约20%。他曾领导欧洲、非洲及中东其他重要业务区域,以及国际运营的商业事务与战略。此次高层调整也被视为诺和诺德在快速变动的市场环境中主动求变的举措。在GLP-1药物领域,礼来公司正加速追赶,两者在糖尿病与肥胖治疗赛道上的竞争日趋激烈。End声明:本公众号所有发文章(包括原创及转载文章)系出于传递更多信息之目的,且注明来源和作者。本公众号欢迎分享朋友圈或大群,谢绝媒体或机构未经授权以任何形式转载至其他平台。转载/商务/投稿 | 联系微信15618157102(sum_Gmi)商务合作稿件征集点击了解详情往期回顾1港股创新药走出至暗时刻2License-out潮来袭:谁是下一个出海爆款?32025 ASCO热点速递 | 信达、荣昌、科伦集体登场!(附ADC/双/多抗最新进展一览表)
高管变更财报
2024-12-18
观点速览:
1、减重效果不是终点,而是起点,更关注体重维持、给药便利性;
2、严重肥胖人群减重效果依然未被满足;
3、从药物研发端去解决产能不足问题;
4、提出减重疫苗概念,或实现1年1次给药;
5、与欧美不同,亚洲人群更喜欢口服日制剂;
6、正在改良制剂技术从而解决司美格鲁肽口服原料药不足问题;
7、司美格鲁肽后,未来管线为复方Cagrisema、双靶GLP-1/Amylin Amycretin;
8、收购了Dicerna,且持续看好小核酸药物;
9、2025 NVO Shopping list:寻找哪些外部创新合作伙伴。
Marcus Schindler
Executive vice president, Research & Early Development and chief scientific officer (CSO) of Novo Nordisk
https://omny.fm/shows/the-top-line/novo-nordisk-s-chief-scientific-officer-looks-to-t
GLP-1 受体激动剂席卷了全球制药界,其中诺和诺德凭借其重磅药物司美格鲁肽(商品名 Ozempic、Wegovy、Rybelsus)一直处于全球领先地位。司美格鲁肽和其他 GLP-1 药物作为2型糖尿病和肥胖的治疗药物已广受欢迎,但诺和诺德并不满足于此,更是在各种新治疗领域、新剂量、新剂型进行探索和临床研究。12月14日,诺和诺德首席科学官兼研究和早期开发执行副总裁 Marcus Schindler 讨论了 GLP-1 的下一步发展以及诺和诺德最关心的系列问题。
Q1、请介绍一下自己
Marcus:我在诺和诺德有双重身份,首先,作为公司高层,会深度参与公司战略规划的所有决策;其次,我分管了早期研究,从一个想法、分子筛选、药物设计、直到概念验证完成(POC,Proof of Concept),然后转移给负责药物开发的团队继续药物的后续开发及临床研究。
Q2、关注到司美格鲁肽11月公开的三期临床数据在治疗肝病取得了积极进展,这是司美格鲁肽接下来的主要研究方向吗?可否告诉我们整体未来策略是什么?
Marcus:GLP-1受体激动剂药物(包括司美格鲁肽)在今天能够成为颠覆式的创新药物,是基于过去几十年的研究和沉淀所换来的。不局限于2型糖尿病、肥胖、心血管,诺和诺德还同步在许许多多的疾病领域展开了人体临床研究。今年,我们尤其是在肥胖领域,明显感受到,GLP-1药物不再是为了解决未满足临床需求,而是更多来自(消费)市场的需求。在我(首席科学)的观点看来,这只是一个开始,我们还有很多可以展开研究的地方,比如:
1)如何实现更好的减重效果,严重肥胖患者减重效果不达预期;
2)如何长期维持减重后的体重,不仅减少体重,更重要是终身保持健康的体重,这是一个很大的话题;
3)如何从研发端解决产能瓶颈问题,比如对药物进行新的设计(新的原料药/分子),从而彻底解决放大生产产能不足问题,减少对高剂量原料药的依赖;
4)我们在开发减肥疫苗,希望能够实现半年或一年给药一次,类似于疫苗,从而大大提高患者的药物依从性,已经发现了一些潜在的技术可以实现这个想法,并相信这会是下一个颠覆式的创新!
Q3、说到长效制剂/疫苗的开发,但我注意到,在今年八月诺和诺德终止了超长效GLP-1/GIP双效激动剂开发(一个月一次),所以接下来计划是什么呢?
Marcus:很好的问题,首先,在今年11月我们与Ascendis签订了全球独家许可协议,根据协议,Ascendis授予诺和诺德使用其革命性的TransCon技术平台,以开发、制造、和商业化诺和诺德在代谢性疾病领域的专有产品,包括肥胖和2型糖尿病。此次合作的主要项目为每月一次的GLP-1受体激动剂候选产品,用于治疗肥胖和2型糖尿病进行治疗;其次,与Ascendis合作的项目,很重要的使命是在于从源头(药物设计)解决原料药产能问题。
Q5、不同国家是否对长效制剂的临床需求是一样的?
Marcus:我们观察到,有很大一部分亚洲人群,对日制剂(口服)的需求量是更大的,而非超长效注射剂,这为我们的全球开发战略提供了更多来自市场终端需求的启发。
Q6、司美格鲁肽产能问题下一步怎么解决?
Marcus:口服制剂对原料药的需求量很大,因为口服片剂每天需要摄入的剂量(3mg、7mg、14mg/次/天)比起一周打一次注射剂(0.25mg、0.5mg、1mg、1.7mg、2.4mg/次/周)大太多了,我们正在努力研究如何减少司美格鲁肽口服制剂对原料药的依赖,比如新的载药技术。
延伸阅读:
礼来多肽供应商9亿欧扩产, 多肽原料药市场缺口到底有多大?
Q7、GLP1+Amylin激动剂Amycretin是否为继司美格鲁肽之后的下一个开发方向?
Marcus:司美格鲁肽之后,我们认为下一个药物开发的方向为胰淀素Amylin,包括复方制剂Cagrisema(即,司美格鲁肽和长效胰淀素类似物卡格列肽Cagrilintide的复方组合),以及双靶点GLP-1/Amylin胰淀素药物Amycretin。
Jo 在此补充一下,MNC对下一代减重Rx的不同开发思路:GLP-1本质是个结构并不复杂的多肽,技术壁垒并不算高,目前已经涌现了大量的仿制药。根据弗若斯特沙利文预测,2030年全球GLP-1激动剂市场规模将达到1150亿美元(2023~2030年复合增长率14.3%)。司美格鲁肽固然不可能一直维持高速增长和超高市占率,且从替尔泊肽开始,GLP-1管线之间的方向就已经发生了分歧。
礼来:第一代GLP-1/GIPR的双靶点药物 替尔泊肽、靶向GLP-1/GCGR的双靶点药物 玛氏度肽、下一步礼来开始卷GLP-1R/GIPR/GCGR三靶点药物 瑞他鲁肽 Retatrutide,在靶点上卷到了极致。
诺和诺德:第一代GLP-1单靶点日制剂 利拉鲁肽、第二代GLP-1单靶点周制剂 司美格鲁肽、下一步Amylin(胰淀素),复方制剂Cagrisema作为两种单靶点联用的复方制剂,可以说是迭代中的中间产物,真正的第三代诺和诺德降维打击药物,应该是双靶点GLP-1/Amylin胰淀素药物Amycretin。
延伸阅读:
附报告|2023世界肥胖地图发布,你想要的超重和肥胖流行病学数据都在这里啦!
Q8、GLP-1药物除了减重还有其他适应症吗?
Marcus:对一些目前小分子/大分子药物无法解决的未满足临床需求,多肽药物或许会给到一个很好的解决方案,我们已经在临床阶段研究多肽药物用于治疗包括神经退行性疾病(AD,阿尔茨海默氏症)、非酒精性脂肪肝、心衰、卒中等。
延伸阅读:
重磅|司美格鲁肽除了减重还能做什么?15种疾病临床进展及专利分析,你想要知道的都在这里了!
Q10、怎么看小核酸药物?
Marcus:小核酸药物是诺和诺德创新药未来版图的重要组成,Dicerna(2021年11月,诺和诺德以33亿美元收购Dicerna)小核酸技术平台拥有能够实现肝脏靶向(GalXC™)和肝外靶向(GalXC-Plus™)的递送平台,这2款递送平台能极好地补充诺和诺德现有的治疗管线。区别于Alnylam公司的GalNAc递送系统,我们Dicerna的GalXC™在过客链中引入了单价GalNAc偶联的独特四环(tetraloop)结构,可增强偶联物的稳定性,同时,还能准确定位多个GalNAc配体,将siRNA成功递送至肝细胞,这种药理学特性极大降低了药物开发的风险。
Q11、AI在药物开发方面是否为公司带来了实质性的帮助?
Marcus:一方面生产效率的提升;另一方面对Wet-Lab(即,实验湿室,需要处理多种化学物质和潜在液体危险的类型实验室)测试、动物试验量能够有一定优化作用;以及包括办公的便利性。但现在下结论,AI技术能够给药物开发和整个产业链带来革命性的帮助,还为时过早。
Q12、未来诺和诺德在寻找合作方或收购标的还有什么计划吗?
Marcus:现有的合作方或已完成收购的标的,目前与合作伙伴们都推进的非常愉快和顺利。早期项目和晚期我们都是关注的。如果一定要给出一个寻找外部合作机会的方向,回到一开始讨论的GLP-1减重领域,我认为是超长期给药的技术合作伙伴,这是我们会乐于寻找的外部合作机会;其次不仅是减重,我们也在积极寻找心血管疾病、MASH(Metabolic dysfunction-associated steatohepatitis 代谢功能障碍相关的脂肪性肝炎)疾病的优秀外部合作机会,以满足未满足的临床需求。
Q13、2024年即将结束,在2025年里,诺和诺德有什么新项目是建议大家关注的吗?
Marcus:当然,2025年我们有新的药物分子陆续进入人体临床阶段,等到时机成熟(有数据)我们将与大家分享更多信息;当然,大家关心的复方制剂Cagrisema临床数据,我们也会与大家分享临床结果;还有,在减重领域,大家不妨关注除了GLP-1以外的更多有趣的药物分子,我们会通过发表更多的papers(文献)与大家分享。
Jo
2024年12月16日晨
END
免责声明:本文仅作知识交流与分享及科普目的,不涉及商业宣传,不作为相关医疗指导或用药建议。文章如有侵权请联系删除。
IND2025主会场授课已被预定,大会第一天下午茶歇前授课时间段预定开始,咨询热线:177 0186 0390。点击图片,查看IND2025首批重磅嘉宾
并购疫苗临床2期临床3期核酸药物
2024-10-07
摘要:病毒样颗粒在大小、形状和分子组成上类似于感染性病毒粒子;然而,它们无法有效地感染宿主细胞。历史上,病毒样颗粒的存在是通过显微镜下的总粒子计数推断出来的,而感染性粒子计数或形成斑块单位(PFUs)则是通过斑块实验得出的;得到的粒子与PFUs的比率通常大于一,很容易达到10或100,表明大多数粒子是非感染性的。尽管它们无法劫持细胞进行繁殖,病毒样颗粒及其携带的缺陷基因组可以表现出广泛的生物学行为:在共感染期间干扰正常病毒生长、细胞杀伤以及激活或抑制先天免疫信号。此外,一些病毒样颗粒随着感染倍数的增加而变得具有生产力,这是粒子间合作的标志。在这里,我们回顾了计数病毒样颗粒和表征它们生物学功能的既定和新兴方法。我们批判性地审视了自然和临床分离物中缺陷干扰病毒基因组的证据,并回顾了它们作为抗病毒治疗的潜力。简而言之,我们强调迫切需要更好地理解病毒样基因组和颗粒在共感染期间与完整功能病毒的相互作用,以及它们对病毒传播、严重程度和持续性的影响。
1.引言
当可以在相同或相关的现象上进行多次观察或测量时,它们的组合或比较可能会导致新的理解或洞见。例如,在天文学中,对行星在不同季节的位置进行多次测量提供了有力的证据,表明宇宙是日心的而非地心的。在生物学中,观察到的在不同岛屿之间和岛屿内部的雀鸟多样性为达尔文关于物种起源的理论提供了证据。在病毒学中,发展焦点形成和斑块形成试验使得能够定量测量感染性病毒粒子水平为焦点形成或斑块形成单位(PFUs);当这些测量与估计总病毒样粒子的估计结合起来,最初使用电子显微镜,粒子与PFU比例的不同显示大多数病毒样粒子不是感染性的。例如,粒子与PFU比例从噬菌体和天花病毒的1-2到疱疹病毒的10,流感的20-50和脊髓灰质炎病毒的30-1000,到水痘-带状疱疹病毒的40,000。SARS-CoV-2的比例基于RNA作为总粒子的代理(基因组RNA到PFU)在10^4到10^6之间。尽管病毒学家知道不同的病毒可以表现出非常不同的粒子到PFU比例,但人们不太了解非感染性病毒样粒子可以表现出多种生物学活性。我们在这里关注作为病毒生长副产品的非感染性病毒样粒子,它们可以在易感细胞中产生,可以在自然界或实验室培养中找到;我们在这里不回顾作为亚单位疫苗或药物传递载体应用的工程化病毒样粒子的广泛文献。通过回顾病毒样粒子的检测和定量表征方法,我们旨在突出它们的多样化功能和活动。
2.计数粒子
2.1. 病毒和病毒样粒子
利用实验室、环境或临床样本中的病毒和病毒样粒子的数量,利用病毒的物理、化学或生物学方面。这些包括病毒粒子的大小、形状和电导率,它们倾向于被一个或多个荧光团特异性标记,以及病毒粒子感染宿主细胞、引起细胞死亡或产生病毒后代的能力。粒子定量的化学和物理方法总结在表1。
表1. 病毒粒子量化技术及其特性。
2.1.1. 透射电子显微镜
透射电子显微镜(TEM)被用来基于形态学识别、分类和定量不同的病毒。历史上,第一个通过TEM可视化的病毒是正痘病毒。其他发现包括腺病毒、肠道病毒、副黏病毒和呼肠孤病毒。TEM成像在诊断天花和水痘等病毒以及识别西尼罗河和严重急性呼吸综合征(SARS)等新出现疾病和爆发中至关重要。最近,通过TEM可视化确认了与COVID-19相关的新型人类冠状病毒的形态特征。
TEM将加速的电子束聚焦在薄的样品上,将图像传输到屏幕上。由于电子的波长较短(0.1埃),它比光学显微镜具有更高的分辨率。因此,TEM可以捕捉到通常大小为20-200纳米的病毒的原子级分辨率。在电子显微镜确定粒子到PFU比例之前,首次测量感染性粒子计数与物理病毒粒子计数之间的比例是针对天花病毒的,使用样本的干重计算“基本体”的数量。基本体最初被观察为来自禽痘病变刮片的微米级粒子,后来被意识到是病毒粒子的组成部分。基本体可以通过离心和沉降分离,基本体的干重除以一个基本体的计算质量。这种计算允许估计误差,反映了样本的异质性。斑块试验用于确定样本的感染滴度;对于天花病毒,这导致平均每一个感染单位有4.2个粒子。多年后,使用电子显微镜发现天花病毒的粒子到PFU比例为每个感染单位1.5个粒子。
对于病毒定量,样本通常与已知大小和浓度的乳胶珠混合。通过手动计数,可以量化几个视场中的病毒颗粒和珠子的数量,并使用病毒颗粒与库存珠子的相对计数,可以确定每毫升病毒颗粒的滴度。在某些情况下,可以通过透射电子显微镜(TEM)检测到有缺陷的颗粒,因为它们的大小和形态与野生型颗粒不同(图1);水泡性口炎病毒(VSV)的干扰缺陷颗粒通常明显截断,呈圆形,直径为76 ± 8纳米,比野生型VSV颗粒的原型子弹形状70 ± 8纳米短204 ± 14纳米。
图1. 透射电子显微镜图像显示了水泡性口炎病毒的缺陷干扰(DI-T)粒子和标准(VSV)粒子。与完整的VSV粒子相比,DI-T粒子是截断的。
虽然透射电子显微镜(TEM)可以产生高分辨率的图像,但其视野范围限制了其通量。更高的放大倍数可以增加对颗粒识别的信心,但采样视野会变小。TEM图像的解释取决于操作者,在病毒定量过程中,意外地计算碎片可能会导致高估。由于依赖人工计数,TEM定量的通量相对较低。颗粒计数需要足够多的颗粒以反映样本中的平均数量,但不要多到让手动计数器难以应对。实际上,这种计数的最低检测限为每毫升10^9个颗粒;样本可能需要在分析前浓缩。样本制备可能复杂且繁琐,通常需要几个小时,图像中的伪影可能来自制备步骤。病毒从细胞碎片中纯化出来,需要准备合适的支撑膜来承载样本,通常使用涂有铜网的支撑膜。比较了三种方法,采用蔗糖密度纯化后负染色、沉淀的树脂包埋上清液的薄片电子显微镜观察,以及负染色后的直接计数;直接计数,即向样本中添加已知滴度的乳胶珠,被发现是最准确和可重复的。
2.1.2. 荧光显微镜
在荧光显微镜(EFM)中,病毒颗粒被荧光染料染色,这些染料可以结合病毒的核酸和蛋白质。荧光显微镜使用能够产生可区分信号的染料,要求是信噪比高于5。DAPI、Yo-Pro-1、SYBR Green和SYBR Gold是常用于染色核酸的染料。蛋白质通过单克隆抗体(MAb)特异性标记;通常,MAb或初级抗体进一步结合一个与荧光染料共价连接的二级抗体。当暴露于不同激发波长的光时,不同的染料会发出荧光。传统上,荧光显微镜受到光的衍射限制,分辨率为200-350纳米。
与TEM相比,EFM设备价格更便宜,样本制备相对简单,这使得EFM成为现场分析样本的首选方法。EFM最常用于计数海洋样本中的病毒。收集和固定样本需要15-20分钟,制备载玻片需要一个小时,计数需要30分钟。使用Yo-Pro或DAPI染色核酸的EFM方法比TEM方法更精确,TEM低估了样本中病毒的数量;使用直接荧光显微镜计数SYBR Gold染色的病毒颗粒是TEM计数的1.34倍。
2.1.3. 电阻脉冲传感
电阻脉冲传感(RPS)是一种基于库尔特原理的颗粒检测方法,该原理由Wallace Coulter六十年前首次开发并发布。该原理利用细胞或颗粒相对于它们悬浮其中的导电水性盐溶液的低电导率;它指的是当颗粒通过孔隙时,阻抗的变化与颗粒体积相当,暂时减少了通过溶液的电流流动,称为阻塞事件。设备的孔足够大,可以让颗粒通过,但又足够小,以至于颗粒通过时孔截面上的电流流动可以被明显干扰。设备通过含有已知大小和浓度的纳米颗粒的溶液进行校准;需要适当的稀释,以最小化两个颗粒同时占据孔隙的可能性。像烟草花叶病毒这样的杆状病毒颗粒可能会旋转以通过固态纳米孔,这在电流读数中产生噪声和不清晰的信号。对于大小不一和聚集的病毒群体,颗粒可能会卡在孔中。这可以通过调整纳米孔的大小或使用可调弹性纳米孔来避免。可调电阻脉冲传感(TRPS)是RPS技术的一种变体,它使用一个包含可以拉伸或放松的孔的弹性膜,与标准的固定固态孔不同。由此产生的孔口大小可以微调,以更好地匹配不同大小的颗粒,这对于具有广泛大小范围的多分散病毒群体尤为重要。确定阻塞事件的大小的能力也允许确定巧合和聚集事件,例如,当脉冲幅度是典型单个脉冲的整数倍时。脉冲形状取决于颗粒的轨迹和移动速度。孔的可拉伸性还允许从阻塞中恢复,以防止堵塞。这项技术主要由Izon Science开发,他们的qViro平台能够提供颗粒浓度的估计,以及颗粒的大小分布和表面电荷。目前,最小的商业纳米孔直径为100纳米,最小检测限为70纳米。在实践中,TRPS的定量范围为每毫升10^7到10^10个颗粒。
2.1.4. 流式细胞术和病毒测量学
流式细胞术将库尔特原理与荧光检测相结合,作为一种高通量方法来测量细胞的核酸含量和大小。细胞通过层流鞘流系统逐个被引导到流动流中,并被激光束激发。产生的偏转光被表征为前向散射(FSC),测量细胞的大小,和侧向散射(SSC),反映其粒度,是表面不规则性或粗糙度的度量。值得注意的是,流式细胞术测量的感染细胞粒度与单纯疱疹病毒产生的病毒滴度相关。标准流式细胞仪无法有效检测500纳米以下的颗粒;包括使用高瓦数激发激光和减小流动室直径、更宽角度的散射光采样以及荧光标记的改进,已使病毒颗粒定量成为可能。基于其起源于流式细胞术,这项技术被称为流式病毒测量学。流式病毒测量学的示意图如图2所示。
图2. 通过流式病毒测量学计数类病毒颗粒。该设备采用流聚焦流将颗粒排成单列通过强激光束,随后进行增强的宽角前向散射(FSC)检测。在传统细胞计数中使用的FSC信号被阻断,通过设置更高的检测阈值来增强宽角FSC信号,降低噪声,从而实现对纳米级颗粒的敏感检测。
流式病毒测量学的一个强大特性是能够在单个病毒粒子的层面上表征异质性。通过利用多种荧光标记,流式病毒测量仪可以表征单个病毒粒子的多种特征。例如,与特定单克隆抗体结合的磁性纳米粒子(MNPs)已被用于捕获和分离病毒粒子,并通过流式病毒测量学分析分离的复合物。这种技术已被用于分析不同来源产生的登革病毒粒子的成熟过程,以及HIV-1粒子的抗原和包膜蛋白组成。MNP捕获的当前限制是只有与MNPs复合的病毒粒子才能被分析,以及立体干扰,这限制了每个病毒粒子上可由抗体标记的抗原数量。许多现代流式病毒测量仪能够使用荧光激活细胞分选(FACS)对异质样本进行排序,这允许对排序的粒子进行进一步表征。流式病毒测量学检测被用于表征经过排序和分析的胡宁病毒粒子的RNA和糖蛋白含量,这些粒子根据不同的大小具有不同的感染性特征。
因为流式病毒测量学是基于流动的且不依赖可视化,所以它是一种快速技术,对操作者的依赖性比EFM少,通量高达每秒2000-6000个粒子,并在不到1小时内为样本产生结果。操作病毒流式细胞仪的技术要求比其他方法如TEM低,并且许多过程步骤都是自动化的。流式病毒测量学的缺点包括由于背景噪声可能导致对粒子水平的高估。巧合事件将低估粒子计数,这就是为什么确定正确的稀释度对于基于流动的分析很重要。巧合事件的概率可以用泊松分布来描述:
其中 r 是通过检测体积的流量(每秒的粒子数),t 是粒子在检测体积内停留的时间。通常,大约 6 × 10^8 个粒子/mL 的高浓度给出两个粒子同时占据检测体积的 10% 概率,浓度越高,巧合的概率越大。流式病毒测量学的最低检测限约为 100 纳米,限制了对更小病毒粒子的检测。
2.2. 感染性病毒粒子
感染性病毒粒子通过空斑测定或终点稀释来定量,两者都涉及可视化细胞病变的宏观区域。能够感染细胞并杀死它们而不产生病毒后代的病毒粒子可以通过它们阻止细胞集落形成的能力来定量,称为克隆形成测定。这些方法在表 2 中进行了总结。
表2. 基于生物学功能和特性的病毒粒子量化技术。
2.2.1. 空斑测定
空斑测定是用于定量感染性粒子或病毒滴度的最广泛使用的方法。该测定通过准备未知滴度的病毒库存的连续稀释,并将其应用于易感细胞单层来进行。吸附和感染启动后,细胞被琼脂覆盖,以将随后的感染传播局限在最初感染细胞的附近;通过结晶紫使细胞死亡的宏观区域称为“空斑”,它使完整细胞着色,未染色的死细胞或感染区域。由于每个空斑都来自一个初始单一感染性粒子的放大,空斑计数,结合已知稀释体积,可以用来计算库存样本中感染性粒子的浓度。感染性病毒滴度以每毫升溶液的空斑形成焦点(PFF)或空斑形成单位(PFU)报告。
2.2.2. 终点稀释
空斑测定的替代方法,终点稀释测定,也用于定量感染性病毒滴度。终点稀释根据使一半的细胞培养物感染的稀释度来测量病毒滴度,以每毫升 50% 组织培养感染剂量(TCID50)表示。使用未知滴度的病毒库存产生 10 倍稀释,将每个稀释度的 1 毫升应用于多个(比如说十个)细胞培养物。孵育后,检查平板是否有细胞死亡或细胞病变效应(CPE)。50% 的培养物表现出 CPE 的稀释度被视为终点;例如,如果在 10^4 倍稀释的 50% 的培养物表现出 CPE,那么库存大约有每毫升 10^4 TCID50 的滴度。在实践中,TCID50 是通过使用 Spearman–Karber 或 Reed–Muench 方法计算得出的,尽管存在其他方法。TCID50/mL 和 PFU/mL 的值不等价,但是可比的。为了一致性,并且由于空斑测定的更广泛使用,我们这里重点关注 PFU 或 PFF 测量的病毒滴度。
2.3. 细胞杀伤粒子(克隆形成测定)
克隆形成测定,也称为集落形成测定,最初由 Marcus 和 Puck 开发,用于确定辐射对细胞的影响。该程序后来被改编为测量流感病毒 A 的细胞杀伤粒子(CKPs)。在克隆形成测定中,细胞在单层中生长。病毒粒子以各种已知的空斑形成粒子(PFP)的倍数附着在单层上,感染的细胞被单分散并种植到培养板中以允许集落形成。集落产生后,它们被固定和染色以计数。从每个 PFP 的倍数中剩余的存活细胞集落分数产生存活曲线。使用生存曲线函数的泊松分布,可以计算 CKPs 的滴度,假设病毒附着接近 100%,并且每个 CKP 将杀死它感染的细胞,导致没有可见的集落。Marcus 和 Sekellick 首次使用细胞杀伤测定来测量 DIP 和 PFP 活性的三个不同峰之间的 CKPs 和 PFPs 的比率;最近克隆形成测定被用来显示流感病毒 A 的一个变体,CKPs 比 PFPs 多七倍。
3.类病毒粒子:出现、功能和普遍性
尽管它们通常被视为标准或感染性病毒培养中烦人的非感染性副产品,但类病毒粒子可以表现出多样的生物学功能;这些包括干扰正常感染、诱导凋亡或宿主细胞杀死,以及激活先天免疫信号。虽然下一代和单分子测序揭示了自然和临床病毒分离物的异质性,无论是在基因组序列还是长度上,研究都集中在携带完整基因组并具有感染性的亚群上。了解遗传变异如何在不同的细胞和感染环境中贡献不同的功能,仍然是自然病毒群体中大部分未探索的方面。了解如何量化类病毒粒子的功能多样性,可能为它们在病毒自然持续存在中的生态和进化角色提供见解,并为它们的管理提出更强大的策略。下面我们讨论不同的类病毒粒子以及如何表征它们的生物学功能。
3.1.干扰缺陷粒子
无法独立复制的病毒粒子被称为缺陷或非感染性粒子。最广泛研究的一类缺陷粒子可以干扰正常或标准病毒感染,如图 3 所示。干扰缺陷粒子,通常缩写为 DI 粒子或 DIPs,最早由 Preben von Magnus 描述,他发现接种了流感病毒第二和第三次培养物的小鼠感染迹象很少;不完整的流感病毒粒子干扰了野生型复制。它们的名字,干扰缺陷粒子,在 1970 年被创造,反映了这些粒子的非感染性但功能。DIPs 及其相关的缺陷基因组在 1980 年代和 1990 年代重组 DNA 技术兴起之前和期间,是复制机制的有用探针。克隆的水泡性口炎病毒(VSV)和其他 RNA 病毒的 cDNA 样本被用来恢复 DIPs,并且在 1992 年开发了聚合酶链反应测定法来测量复制回退和其他缺陷病毒基因组。
图3. 缺陷干扰粒子(DIPs):出现和生物学。DIPs源于正常病毒感染(左侧),它们在与完全感染性粒子共感染的细胞中放大(中间),单独的DIPs无法有效感染细胞(右侧)。
3.1.1. DIP的出现
DIPs作为病毒复制和感染的副产品而出现。当细胞被病毒感染时,病毒复制复合体使用病毒基因组或反基因组作为模板,指导合成全长基因组模板,最终包装成病毒后代粒子。模板中被相似或相同的短序列(少于20个核苷酸)所夹的区域可以使得延伸中的复制复合体在重复之间跳跃,导致链内或链间重组,留下在结果基因组模板中的删除。如果删除或其他突变导致一个基本病毒功能的丧失,那么复制产物在功能上就是有缺陷的;产物是DVG或缺陷病毒基因组。DVGs长期以来与DIPs有关。最近,DVGs在自然和临床病毒分离物中被发现,部分得益于深度和单分子测序技术的进步。人们对DVGs的生物学功能越来越感兴趣,特别是它们在疾病发展和严重性方面的可能作用;这些细节在其他地方进行了审查。DVGs的两个方面有助于它们在自然中的持续存在至今得到了较少的关注:它们的出现和潜在的进化能力。
DVG种群的结构和进化将取决于它们的生成率。在这里,区分突变率和频率(重组或删除)是有用的,这是由Drake和Holland定义的。具体来说,突变率反映了由复制机制及其处理模板引起的生化事件的机会,作为细胞内环境的函数。突变率通常报告为点突变的发生概率每复制一个核苷酸;对于删除,它是每个基因组复制的概率。例如,T7噬菌体的聚合酶复合体的删除率估计为每复制10−8到10−6;更大的删除在更低的速率下丢失,基于与噬菌体连接酶中设计的不同长度序列的删除相关的功能恢复的测量。
与突变率相比,突变频率取决于突变率以及由此产生的DVG相对于种群中其他基因组的富集能力。简而言之,给定DVG的频率是一个群体水平的度量,受达尔文选择的影响。DVGs在实验室和自然中的选择和持续部分取决于它们作为DIPs在细胞间复制和传播的能力。突变频率的估计可以通过量化DIP水平相对于来自单个感染细胞的完整感染粒子的后代来获得;在实践中,这是对来自小斑点的VSV种群进行的,估计为10−8到10−7,这个值将反映RNA重组形成DVGs的速率组合,以及DVGs可以被包装并与完整病毒共感染细胞以复制和作为DIPs传播的比例。同样,噬菌体T7的DVGs的删除频率大约为10−8,它们删除了编码T7 RNA聚合酶的基因。这样的DVGs及其DIPs可以在重组宿主细胞中比全长基因组更快地繁殖,这些宿主细胞通过转录提供这种必需的噬菌体酶。这种删除频率的估计是基于在小斑点噬菌体群体中尽早检测到DIPs的量化。如果允许小斑点扩大,就会出现不同的DVG和DIP谱系,每个谱系都源自相同的祖先噬菌体基因组;它们沿着不同的空间方向丰富到不同的程度,因为斑点呈放射状扩展。通过使用工程化和人工宿主细胞环境,这项工作揭示了在自然感染中否则无法检测到的DVGs的一个子集的出现和进化。DVG和DIP进化的其他方面已经通过其他连续传代或持续的细胞和病毒培养揭示。这样的培养已被证明促进了较长DVGs的出现、富集和由较短的更高适应性DVG变体取代。番茄丛矮病毒(TBSV)在植物上的连续传代、噬菌体T7在细菌上的连续培养以及VSV在哺乳动物细胞上的连续传代培养期间的DVGs进化已经得到证明。对于TBSV,增强DVG稳定性和包装效率的机制并没有解释较短DVGs的适应性优势。这些结果表明,由于复制速率更高,较短的DVGs比较长的DVGs获得了选择优势。一种基于库的方法,为不同流感A病毒的不同片段生成并评估了广泛的DVGs长度,支持这一趋势,较短基因组片段的复制效率更高。然而,在其他情况下,流感A的选择过程可能有利于DVGs的包装而不是完整基因组。在小鼠肝炎病毒、伪狂犬病病毒和禽类呼肠孤病毒中发现了有效包装的DI基因组,它们在包装竞争力上有所不同。进一步的现象包括超级启动子的出现,它创造了对有效感染所需的基因组片段的不平衡,或者一些不需要删除就可以有效复制和干扰正常病毒生长的因素的组合。
通过荧光原位杂交(FISH),已经阐明了全长病毒基因组及其DVGs在细胞中的空间分布对于仙台病毒。在富含全长病毒基因组的细胞中,病毒基因组与细胞运输机制相互作用,并聚集在核周区域。相比之下,在富含DVGs的细胞中,缺陷基因组没有显示出与运输机制的相互作用,它们在细胞质中均匀分布。结果,含有全长基因组的细胞产生了DVGs和完全感染性的病毒颗粒,而DVG含量高的细胞病毒颗粒产生较差。这种差异可能由DVGs在细胞内的定位差异来解释。
病毒及其缺陷基因组的更复杂的进化动态可以表现出共同进化。DVGs和DIPs的出现可以创造一个选择环境,选择抵抗干扰的完整病毒。例如,完整的VSV突变体可以通过影响基因组复制和包装的突变来抵抗DVG和DIP干扰。此外,这种对DIP有抵抗力的完整病毒进一步创造了一个环境,促进了新变体DIPs的出现和富集;在数百个未稀释的连续传代培养中,完整病毒及其相关的DIPs表现出多轮共同进化。其他对DIP有抵抗力的完整病毒,包括狂犬病、淋巴细胞性脑膜炎、辛德比斯和西尼罗河病毒,具有表现出类似共同进化动态的潜力。
最后,DVGs可以进化为以一种方式合作,使它们摆脱对完整病毒的任何依赖以实现生长。在高感染度下连续传代的口蹄疫病毒(FMDV)培养物使DVGs和DIPs的出现和富集成为可能,它们通过互补感染具有传染性;通过感染同一细胞,它们在转录中提供了复制、包装和其他传播其DVGs及其相关缺陷合作颗粒所需的功能。这种单独包装的病毒基因组片段之间的合作在自然中多部分病毒中发生;多部分病毒的生态和进化最近已经回顾。功能分离过程的反面,即单独的病毒衍生缺陷RNA片段重组形成完整的完全感染性单一部分病毒也已经得到证明;与仙台病毒相关的黄瓜坏死番茄病毒的DVGs和相关病毒在植物衍生的原生质体中共感染,形成了在整株植物中具有感染性的嵌合体重组体。同样,工程化的辛德比斯病毒DVGs在宿主细胞中无法自行复制,但它们可以相互重组形成完全感染性的病毒。保留或缺乏复制和包装功能的遗传元素之间的结果合作具有超越病毒学的更广泛和更深远的意义,包括生命起源中的关键转变。
3.1.2. DIP干扰的度量
DIP对病毒生长干扰的最简单模型假设单次打击行为,即与完整病毒和至少一个DI颗粒共感染的细胞只能产生DI颗粒后代,但实验观察到的干扰行为并不那么简单。干扰的程度取决于剂量,即共感染细胞的标准病毒和DI颗粒的相对水平。例如,在与DI颗粒共感染期间,较高浓度的标准流感A病毒可以部分恢复标准病毒的产生。此外,通过干扰对标准病毒复制至关重要的资源的生产,DI颗粒和基因组在较高剂量下可以抑制它们自己的复制。对VSV的定量研究提供了干扰过程对DI基因组和颗粒产生的复杂反馈的证据。使用放射性同位素对DI和标准颗粒进行代谢标记,通过超速离心进行基于密度的分离,以及对DI和标准颗粒群体的定量,发现增加DI颗粒的输入显著降低了标准颗粒的产生,但最高产量的DI颗粒是在输入剂量下得到的,这些剂量对标准颗粒产生表现出最小的干扰。使用空斑减少测定和双层滴定的进一步定量研究揭示了DI输入剂量下高度非线性的病毒和DI颗粒产生,如图4所示。具体来说,对于固定的病毒剂量和增加的DIP输入,标准病毒产生水平下降,正如预期的那样;更多的DVG可以竞争复制和包装资源,导致标准病毒产生资源减少。然而,在最高水平的DIP输入下,标准病毒产生表现出部分恢复,并且增加的DIP剂量导致DI颗粒产量下降。这种现象以前被描述为“干扰的干扰”。类似的现象也被描述为工程化的条件复制或有缺陷的干扰HIV,它“在分子上自伤”;DIP如此有效地抑制病毒,以至于它自己的产生被抑制。
图4. 共感染细胞产生的(a)标准病毒和(b)DI颗粒的产量是DI颗粒输入的复杂函数。所有细胞都与标准病毒(MOI 20)和不同输入水平的DI颗粒共感染。通过空斑测定和空斑减少测定确定了标准病毒和DI颗粒的水平。
3.1.3. 体外的DIPs
已发现许多DNA和RNA病毒形成DIPs。形成DIPs的DNA病毒包括单纯疱疹病毒、伪狂犬病病毒[82]和Ageratum黄脉病毒。体外产生DIPs的RNA病毒包括水泡性口炎病毒、新城疫、麻疹、流感、小鼠肝炎病毒,以及最近的SARS-CoV-2。SARS-CoV-2的缺陷病毒基因组在培养中容易形成,但尚未显示它们在多大程度上干扰或与完整的冠状病毒生长相互作用。
3.1.4. 体内的DIPs
最初认为大多数病毒的DIPs的发现与自然感染无关,因为它们起源于体外,但在过去15年中,初步证据显示体内存在DIPs。例如,在感染患者的血清中发现了只包含30和50端的包装关键调控元件和大量内部删除的登革病毒RNA短片段。使用患者血清感染蚊虫培养物,培养物中发展出了相同的RNA片段,这为RNA片段作为完全包装的DIPs传播提供了可能性。体外转录的RNA与体内样本相对应,被证明可以被包装成类似病毒的颗粒,并在存在野生型病毒的情况下传播了三次。在含有这些短RNA片段的登革病毒制剂中,野生型病毒的产量降低了,为干扰提供了支持,也是体内DIPs最有说服力的证据。最近,相同的RNA来自血清被用来感染支持登革病毒生长的细胞系,同样,感染性病毒的产量降低了;此外,RNA被证明可以诱导RIG-I、MDA5和干扰素的活性,符合DIPs的免疫刺激性活性(见第3.1.5节)。来自流感A患者鼻咽样本的RNA携带了大量内部删除,其30和50端的重叠序列,并保留了病毒包装信号,类似于体外产生的DIPs的RNA。在同一个接触组的两个患者中发现了相同的RNA序列,指向了缺陷病毒传播的可能性。对于流感A病毒自然DIPs更有说服力的案例将随着进一步的研究而出现;例如,干扰标准病毒复制或生长尚未得到证明,DI颗粒而不是DVGs仍然需要被分离和表征。在缺陷病毒基因组数量与流感A病毒感染的严重程度之间发现了反向关系;具体来说,临床分离物显示与致命感染相关的DVGs水平较低,而轻症病例中的DVGs水平较高。这些DVGs通过在宿主中诱导保护性先天免疫反应而起作用。目前尚不清楚这些DVGs是否被包装并与活病毒一起传播。在人类呼吸道合胞病毒(RSV)中发现了免疫刺激性缺陷病毒基因组(iDVGs),它们在人类细胞中触发了抗病毒免疫反应,这是DIPs的特征(见第3.1.5节)。然而,研究集中在DVGs而不是DI颗粒上;体内RSV的DIPs仍有待分离和表征。对中东呼吸综合征(MERS)冠状病毒感染患者的鼻咽样本进行直接测序,发现了两种病毒变体,它们内部删除导致病毒蛋白的截断,生物信息学分析表明这些变体在包装上存在缺陷。尚未进行功能性测试以提供证据,证明这些变体是DIPs。在SARS-CoV-2的情况下,在只有轻微症状或无症状的多名患者中发现了携带大量相同删除的基因组。目前尚不清楚DVGs在COVID-19的严重性中可能扮演什么角色。
3.1.5. DIPs与免疫反应
最近的综述总结了DIPs在激活免疫反应中的作用。具体来说,DIPs诱导产生I型干扰素(IFN),在先天免疫中发挥关键作用。从机制上讲,模式识别受体RIG-I更倾向于与较短的基因组结合,如DIPs中的DVGs,然后诱导产生I型IFN和其他促炎细胞因子。除了激活免疫反应外,RIG-I的激活还可以诱导凋亡;因此,DIPs有潜力作为抗病毒以及抗肿瘤治疗。
除了激活与IFN相关的先天免疫反应外,DIPs还可以抑制IFN。在细胞系中超量产生IFN的条件下,对8种流感A病毒(IAV)菌株进行了检测,使用干扰素剂量-反应曲线量化了它们对IFN水平的影响。被命名为IFN诱导颗粒(IFPs)的菌株诱导了IFN,而被命名为IFN诱导抑制颗粒(ISPs)的颗粒抑制了IFN。IFP活性似乎是由于存在双链RNA分子,因为单链RNA颗粒没有诱导IFN;双链RNA对IFN诱导的这种影响已经得到了很好的证实。通过紫外线照射增强了IFN的诱导,这有助于将ISPs转化为IFPs,并证实IFN活性不依赖于病毒复制或感染性。在NS1上有缺失的IAV菌株被发现比亲本菌株高20倍地诱导IFN;NS1对IFN诱导的抑制作用现在已经得到了很好的证实。这些发现突出了DIPs如何通过它们的双链RNA结构或表达的抗IFN功能来激活或抑制宿主防御反应。
最近,在RSV鼻分泌物中检测到的DVGs与临床患者对病毒的反应有关。在儿童中,DVGs与更高的病毒载量和更强大的促炎反应相关。然而,在成人中,临床反应是基于检测到DVGs的时间,而不仅仅是它们的存在。在感染过程中早期检测到的DVGs与轻度疾病相关,而晚期检测到的DVGs与严重疾病相关。有DVGs的患者细胞因子表达增强,包括与DVGs和DIPs诱导IFN一致的IFN alpha。
除了在触发先天免疫反应中的作用外,对仙台病毒的研究还提供了DIPs和DVGs在适应性免疫中发挥作用的证据。富含DIPs的仙台病毒库存被证明可以通过帮助上调细胞因子活性来诱导人和小鼠细胞的树突状细胞成熟。这条途径独立于IFN工作,并表明DIPs可能作为疫苗佐剂,刺激树突状细胞成熟。随后的研究表明,DIPs可以上调树突状细胞上的模式识别受体活性,然后刺激T细胞激活。此外,仙台DIPs被用作灭活流感A疫苗的佐剂,其中DIP RNA增强了抗流感血凝素特异性IgG的产生,表明这些DIPs具有广泛的佐剂活性。
3.1.6. DIPs作为抗病毒治疗
由于它们能够干扰标准病毒感染,DIPs被提出作为潜在的可传播抗病毒治疗。例如,一种带有大内部删除的IAV克隆DI病毒,称为244 DI病毒,已经通过鼻腔给药给小鼠,并发现可以保护免受几种IAV菌株的感染。在雪貂中,通过鼻腔给药的244 DI减少了发热、体重减轻、呼吸症状和与感染对照相比的标准病毒的传染性负荷,为244 DI病毒可以作为有效的抗病毒提供了进一步的证据。另一种源自IAV的DI病毒,OP7,当与IAV共感染时表现出强烈的干扰,基于释放的病毒粒子的感染性降低,支持OP7作为潜在的抗病毒治疗。此外,已经设计了治疗性IAV颗粒,将DI基因组片段传播,将正常的IAV感染转向非传染性颗粒的产生,在动物模型中证明了对致命病毒的保护。
从人类免疫缺陷病毒(HIV)中工程化的DIPs已被证明可以减少野生型HIV的复制。基于来自撒哈拉以南非洲的HIV数据的多尺度模型表明,所谓的治疗干扰颗粒(TIPs)可以在接下来的50年内将HIV/AIDS的流行率降低30倍。提出的TIPs将是慢病毒载体,它们缺乏自我复制所需的基因,但保留了HIV包装信号,因此载体可以从感染HIV的宿主中移动,并与野生型病毒竞争资源。正常DIP与这些TIPs的区别在于,TIPs将被工程化以具有大于1的基本生殖数(R0);TIP将产生比野生型病毒更多的基因组RNA(gRNA),利用由野生型病毒产生的病毒资源,大规模降低野生型病毒复制、疾病进展和传播。
最近,DIPs作为针对黄病毒的疗法已被探索。一个生产细胞系使用慢病毒和逆转录病毒载体的组合稳定地产生无病毒的登革病毒DIPs;经过纯化和浓缩后,DIPs在与登革病毒共感染的细胞中显示出抗病毒活性。这样的生产系统和DIP活性可能对多种病毒都是可行的,为基于DIP的抗病毒治疗提供了潜在的平台技术[150]。对于寨卡病毒,DVGs通过计算分析来确定哪些DVG序列作为TIPs最有效。在连续传代过程中频率增加的DVG序列被识别并测试了对野生型病毒的干扰活性。然后,DVGs被工程化到VLPs中,显示出可比的干扰,证实了它们作为TIPs的可能用途。VLPs被用来感染小鼠和蚊子,减少了蚊子中野生型病毒的传播高达90%,并减少了小鼠大脑和卵巢中的病毒载量。在西尼罗河和黄热病病毒的传代培养中也发现了类似的DVGs,表明这种方法可以推广到虫媒病毒,可能推广到其他病毒。
COVID-19大流行促进了针对人类冠状病毒,特别是SARS-CoV-2的合成DVGs和DIPs的发展。基于已知的冠状病毒DIPs,设计了一个设计,结合了50和30端,以及来自SARS-CoV-2基因组的假定包装信号。片段被组装在一个框架中,作为DNA合成,插入到质粒中,转录形成gRNA,并电穿孔到感染SARS-CoV 2的细胞中。DI基因组被发现比野生型复制速度大约3倍,同时在24小时内将野生型病毒的数量减少大约一半;下一步将在动物模型中评估该策略。一项更深入的研究设计了基于机制建模的治疗性干扰颗粒,合成了包括SARS-CoV-2基因组的50和30区域、包装信号和荧光报告器的TIPs。TIPs在细胞培养、人类肺类器官和仓鼠中进行了测试。工程化的TIPs被发现在细胞中抑制SARS-CoV-2复制10到100倍,在仓鼠的肺部抑制病毒负荷10倍,并在感染前或感染后给药时减少炎症和严重疾病。这些令人鼓舞的结果为最终在人类临床试验中的测试和优化提供了希望。
最终,工程化的DIPs可能有助于促进公共卫生的新范式:具体来说,是那些因其广泛的保护效果而减少传染病的疫苗。例如,基于脊髓灰质炎DVGs的肠道病毒治疗干扰颗粒(eTIP1)在小鼠的呼吸道内触发了一种抗病毒状态,这种状态抑制了病毒复制,并保护小鼠免受肠道病毒、流感和SARS-CoV-2的感染。通过在病毒感染前24至48小时内或感染后给予eTIP1,实现了保护,这种保护是通过I型IFN信号和持续数周的病毒特异性中和抗体反应介导的。这类策略因其广泛的抗病毒效果,有潜力对抗病毒变种的出现。
3.2. 半感染性粒子
流感A病毒(IAV)通常表现出粒子与PFU(每毫升菌落形成单位)比率为10到100,所以IAV群体中超过90%的粒子是非感染性的。在一个关键实验中,IAV粒子可以通过启动基因表达来在单个易感细胞中启动感染,但大多数粒子随后未能表达一个或多个必需蛋白;基于对邻近细胞缺乏感染传播,这些细胞也没有产生病毒后代。少数被单个粒子感染的细胞显示出了有效的感染。然而,当细胞被多个粒子感染(MOI为5)时,大多数细胞都是有效的。由于被单个或多个粒子感染的细胞表现出了要么效率低下要么高效的产生,这些入侵实体被命名为半感染性粒子(SIPs);见图5。
图5. 半感染性粒子的三种可能命运。(a) 一个粒子感染了它的宿主细胞,但至少有一个必需基因未能表达,所以细胞没有产生后代,(b) 一个粒子感染了它的宿主细胞,所有必需基因都得到了表达;细胞产生了后代,(c) 两个或更多的粒子共感染一个宿主细胞,基因或功能缺陷通过互补得到克服,细胞产生了病毒后代。
通常,SIPs与DIPs不同。SIPs在共感染期间不会干扰标准病毒的产生,并且它们缺乏作为DIPs基因组特征的大型内部缺失。此外,大多数IAV的SIPs似乎是完全完好无损的;它们携带了八种病毒RNA基因组片段中的每一种。那么,为什么只有一小部分IAV粒子能有效地感染细胞呢?
高颗粒与PFU(每毫升菌落形成单位)的比率可能源于缺乏感染所必需功能的粒子,如DIPs,但也可能源于SIPs,根据它们的基因组序列、生物分子组成和结构,人们可能会期望它们是完全功能的。然而,单细胞测量和定量模型表明,在IAV SIPs和其他病毒的混合开关行为中,随机或嘈杂过程扮演了重要角色。
从细胞群体中测量的平均细胞行为掩盖了可以通过单细胞水平测量最好理解的显著异质性。在可能最极端的掩盖行为案例中,大量感染细胞的亚群未能产生感染性病毒后代;具体来说,表现出早期病毒基因表达的感染细胞未能为30%的VSV感染细胞、80%的牛痘感染细胞和90%的IAV感染细胞产生可检测的病毒后代。此外,剩余产生感染性病毒后代的细胞表现出产量分布,从10倍到1000倍不等;这样的广泛分布已观察到在被噬菌体、VSV、脊髓灰质炎和IAV感染的细胞中。
计算模型揭示了与病毒-细胞相互作用相关的不同细胞内过程的嘈杂行为的显著效应:在裂解-溶原性决定中的随机基因表达、病毒中间体内部水平的异质性,以及感染对进入病毒基因组降解的敏感性。IAV进入病毒基因组的随机降解的证据结合了来自单个细胞的病毒RNA、IAV细胞的病毒蛋白和计算机制建模的实验测量。进一步的实验和细胞质扩散传输的随机建模已经确定了病毒-内吞体融合的时间点和释放病毒基因组到细胞核的相关扩散距离,作为有效病毒感染的关键瓶颈。为了克服基因组片段的降解,需要多个病毒粒子进行有效感染;具体来说,对于IAV,大约需要2到5个病毒粒子进入一个细胞以使其有效感染。除了克服基因组片段降解的损失外,IAV的研究还展示了病毒RNA依赖的RNA聚合酶的二聚化如何可能需要克服病毒RNA复制的宿主特异性障碍,为集体分子相互作用在必需病毒处理中的作用提供了证据。
病毒粒子之间需要合作的需求并不仅限于IAV。对牛痘病毒的优雅单细胞研究结合了纳米级流体操控、检测重组荧光病毒粒子和原子力显微镜(AFM)来剖析感染的早期阶段。基于单粒子输入的效率,人们可以预测两粒子、三粒子及更多粒子输入的效率;与预测的偏差表明整体感染是合作的。然而,只有48%的表面结合病毒进入,与结合的总粒子数无关,表明进入不是合作的。此外,进入的那些中,80%无法直接检测到基因表达,不到2%的表面结合病毒粒子能够完成整个病毒生命周期并指导后代病毒粒子的组装。
除了不可避免的单感染细胞的嘈杂或随机行为外,其他机制也可能促成半感染表型。例如,电子显微镜显示IAV的基因组片段包装可能是不完整的,经常省略编码PB2的片段。此外,IAV具有高突变率,IAV的多个基因突变可能使其易受先天免疫系统的关闭。IAV突变也可能对IAV基因产物与其许多必需宿主因子的相互作用产生不利影响。
3.3. 非感染性细胞杀伤粒子
感染性病毒粒子通常在产生病毒后代的同时作为副产品杀死宿主细胞。那些杀死宿主细胞但未能产生可检测后代的粒子被称为非感染性细胞杀伤粒子(NiCKPs),如图6所示。一般来说,这种粒子可以通过集落形成测定法(第2.3节)来量化,未被杀死的细胞会产生可以轻易量化的菌落。
图6. 一个非感染性细胞杀伤粒子可以在细胞中触发凋亡。
3.3.1. NiCKPs特征
NiCKPs的证据基于VSV粒子的细胞杀伤和形成空斑的测量差异。未能通过空斑测定产生可检测后代的CKPs是“有缺陷的细胞杀伤粒子”或NiCKPs,它们的产量比PFPs高出5到9倍。此外,NiCKPs和DIPs都未能产生病毒后代,但DIPs也未能杀死细胞,并且DIPs未能干扰NiCKPs对宿主细胞的杀伤。IAV的NiCKPs和DIPs表现出类似的行为。单个NiCKP足以杀死一个细胞,并且UV灭活研究表明,不同IAV粒子的UV灭活理论靶标大小不同。更具体地说,正常的感染粒子的UV靶标约为13,600个核苷酸(nt),NiCKPs的UV靶标约为2400 nt,与其中一个聚合酶亚基基因一致,DIPs的UV靶标约为350 nt,与与干扰相关的最小缺陷病毒基因组一致。最后,将感染力、干扰和细胞杀伤的测定结合起来,用于表征在高倍数传代期间IAV群体的动态;一个纯粹的感染粒子初始群体在第一次传代期间下降了100倍以上,被DIPs(68.5%)和非感染性CKPs(31%)所取代。在第二和第三次传代期间,DIPs继续丰富(超过90%),以牺牲非感染性CKPs(低于10%)和感染性粒子(~0.01%)为代价,而总粒子计数保持相对稳定和高,超过109,在传代过程中。
3.3.2. 粒子适应度和毒力
尽管将病毒粒子描述为感染性或细胞杀伤性暗示了全有或全无的行为,但这样的描述是连续行为的简化。病毒在特定条件下的适应度通常指的是其复制能力,而其毒力指的是其杀死细胞的能力。病毒的适应度和毒力取决于其培养条件。例如,当一个口蹄疫病毒(FMDV)克隆在大量群体中传代培养时,产生了高适应度高毒力的病毒;然而,随后的空斑到空斑传代产生了适应度的显著损失,毒力损失很小。这种低适应度高毒力的FMDV粒子类似于为IAV描述的非感染性CKPs。同样,对FMDV适应度和毒力影响的点突变研究提供了这些表型是如何编码的指示;适应度可能受到基因组任何区域突变的影响,而毒力似乎局限于病毒基因的子集。因此,毒力可能比适应度更能抵御有害突变的影响。对其他病毒的适应度和毒力的研究表现出各种行为;病毒适应度和毒力正相关,但VSV有值得注意的例外,并且它们在烟草蚀纹病毒对植物的感染中似乎缺乏相关性。
3.3.3. NiCKPs的应用
跨越广泛毒力或细胞杀伤范围的病毒样粒子可能在人类健康方面有有用的应用。非感染性CKPs,其适应度低但仍然有毒力,可能在细胞杀伤是可取的但不希望感染扩散的疗法中有用,如用于治疗癌症的溶瘤疗法。例如,工程化的高减毒VSV,产生很少后代并且小空斑,选择性地感染并杀死植入SCID小鼠的人胶质瘤;在美国,VSV的溶瘤疗法已在I期临床试验中进行了测试。其他人主张采用在宿主中迅速传播的溶瘤策略,因为它们具有高适应度,但毒力最小。除了病毒适应度和毒力水平之外,其他因素,包括在健康和目标宿主细胞和组织中先天免疫激活的程度,也将在溶瘤疗法的设计中很重要。
4. 讨论
粒子与PFU比率。我们应该给远大于1的粒子与PFU或基因组与PFU比率赋予什么意义?量化这些比率需要在相同样本中测量PFUs之外,还需要总粒子或基因组水平。这种多重测量最常在“干净”的实验室培养的细胞和病毒样本上进行,而不是在“脏”的或未培养的环境样本上。这些培养通常也使用转化的细胞系作为感染宿主,因为它们可以产生强大的病毒滴度或通过空斑计数容易地可视化和量化传染性粒子。这些病毒株通常与它们的自然对应物相去甚远,经过多代传代培养在实验室中筛选出来,以产生强大的滴度或容易观察到的空斑。报道的粒子与PFU比率的广泛范围也反映了缺乏参考比率和标准化。尽管如此,我们认为这些比率的效用不在于它们的具体数值,而在于当这些比率大于一时,它们突显了两个可能对自然界中的病毒及其宿主具有普遍性的特征:(i) 非感染性病毒样粒子容易作为感染的副产品产生,(ii) 这些粒子可以表现出多样化的生物活性。
自然界中的缺陷病毒基因组。我们已经强调了体内或来自自然和临床分离物的病毒样粒子和DVGs的多样化生物活性的引人注目的例子。这些包括半感染性粒子,这些粒子必须与多个粒子感染同一细胞,才能在小鼠中对流感A病毒产生有效的感染。此外,具有相似或相同缺陷的病毒基因组已被从不同地理位置的人类患者和蚊媒中分离出来,为登革病毒感染提供了长期跨时空传播的证据。最近,人类中产生的DVGs的存在已与先天免疫反应的诱导相关,并与不同程度疾病严重程度的呼吸道合胞病毒的年轻和老年患者联系起来。最后,值得注意的是,与临床样本中更高病毒载量相关的多瘤病毒的DVGs。我们预计,在未来几年中,对患者和环境分离物的深度测序和单分子测序将揭示更多与不同临床结果相关的DVGs的例子。一个持续的挑战将是表征这些DVGs可能具有的多种生物活性及其潜在机制。这些机制可能在分子到粒子的尺度上起作用,甚至影响多粒子群体。
病毒和病毒样粒子的工程研究。工程化的实验系统可以比自然或患者感染更深入地研究病毒-细胞行为。例如,工程包括设计、合成和应用报告基因组。单报告病毒可用于在低MOI下感染细胞,结合荧光激活细胞分选(FACS)分离被单个病毒粒子感染的单细胞,这些细胞可以进一步研究,以研究感染启动病毒基因表达但未能产生病毒的感染,以及有效感染的产量分布。病毒和DIPs的工程双色报告可以使得在不同共感染条件下,病毒编码和DIP编码基因表达的量化成为可能;微孔技术和相关的图像获取和分析流程可以促进对这种单细胞行为随时间的跟踪。或者,工程可能涉及将湿实验室感染“滴定”与数学或计算模型相结合;DIP剂量对共感染细胞产生病毒和DIP活性的影响揭示了非直观的行为,特别是在高DIP剂量下,数据的数学建模表明单个VSV DIP无法完全抑制活病毒的产生。最后,工程化人造环境以探测孤立的单细胞随时间变化,与更常见的培养细胞群体相比,可以揭示DIPs的微妙之处;群体中的细胞共感染比它们的单细胞对应物受DIPs的抑制要小,这表明细胞群体内的营养或信号效应可以调节DIPs对病毒基因表达和生长的抑制作用。
病毒如何成为“半感染性”的?单细胞行为的数学建模突出了病毒基因组的嘈杂或随机降解如何导致原本相同的感染细胞要么产生病毒,要么不产生。更机械和复杂的计算模型表明,在单细胞感染期间,转录本、蛋白质和基因组的随机合成可能导致病毒产量的广泛分布,这已经在实验中观察到。这些模型为半感染性粒子表型提供了一个合理的贡献因素。更广泛地说,Klasse很好地阐述了与粒子相关的感染挑战;“关于病毒粒子感染性的全有或全无的假设是有缺陷的,应该被允许感染倾向谱的描述所取代”。换句话说,病毒-细胞相互作用的结果可以表现出感染相关行为的广泛分布,包括未能产生病毒后代。
回归自然。通常用于培养病毒的转化细胞系在先天免疫信号和病毒生长抑制方面常常存在缺陷;这些包括,例如,Vero、BHK、HEK293T和HeLa。尽管已知在这些转化细胞系上病毒基因组复制、病毒蛋白的合成和处理以及病毒关闭宿主细胞过程与它们在原代细胞中的行为相当的案例,但这类研究很少;病毒生长和感染传播的比较研究突出了在宿主细胞保留先天免疫信号时更大的抑制作用。在原代细胞和组织上病毒传播产生与转化细胞培养产生的病毒样粒子多样性的程度仍是一个未解决的问题。
培养原代细胞技术上具有挑战性,它们对病毒的感染往往产量较低且高度可变。然而,低产量和可变性可能反映了更接近自然行为的感染特征。其他更自然且难以实施的培养环境包括促进原代细胞分化的条件。例如,原代人类气道上皮细胞的空气-液体界面(ALI)培养促进了它们的分化,形成了假复层上皮;存在杯状细胞和粘液产生细胞,以及功能性纤毛,上皮对人类鼻病毒C型易感。再生医学和药物测试领域,将人类诱导多能干细胞(iPSCs)分化为类组织细胞,提供了更自然和可控的环境,以研究病毒生长和感染传播行为。例如,iPSCs已分化为支持丙型肝炎病毒完整生命周期的类肝细胞,包括对感染的炎症宿主反应。类似的方法已被用于创建人类神经前体细胞以研究寨卡病毒和人类3D肺芽类器官以研究呼吸道合胞病毒。随着这些技术的成熟和更广泛的应用,它们可能有助于揭示在转化细胞系培养中缺失的病毒和病毒样粒子相互作用在生长和传播期间的方面。
结论。对许多病毒测量的大量粒子与PFU比率表明,绝大多数病毒样粒子无法有效地感染它们的宿主细胞。然而,尽管是“死的”,病毒样粒子在其他方面可能非常活跃:利用完整病毒建立的宿主细胞内的资源,干扰正常病毒生长,激活或抑制宿主细胞的先天免疫信号,杀死宿主细胞,并与其他“死”病毒合作产生感染性后代。对病毒样粒子的这种理解主要是从粒子和宿主细胞水平的研究中获得的。一个巨大的挑战仍然是理解这些功能和活动如何影响植物、动物和人类宿主中传染病的发展、严重程度、传播和持续存在。
信使RNA疫苗
100 项与 Marcus Research Laboratory, Inc. 相关的药物交易
登录后查看更多信息
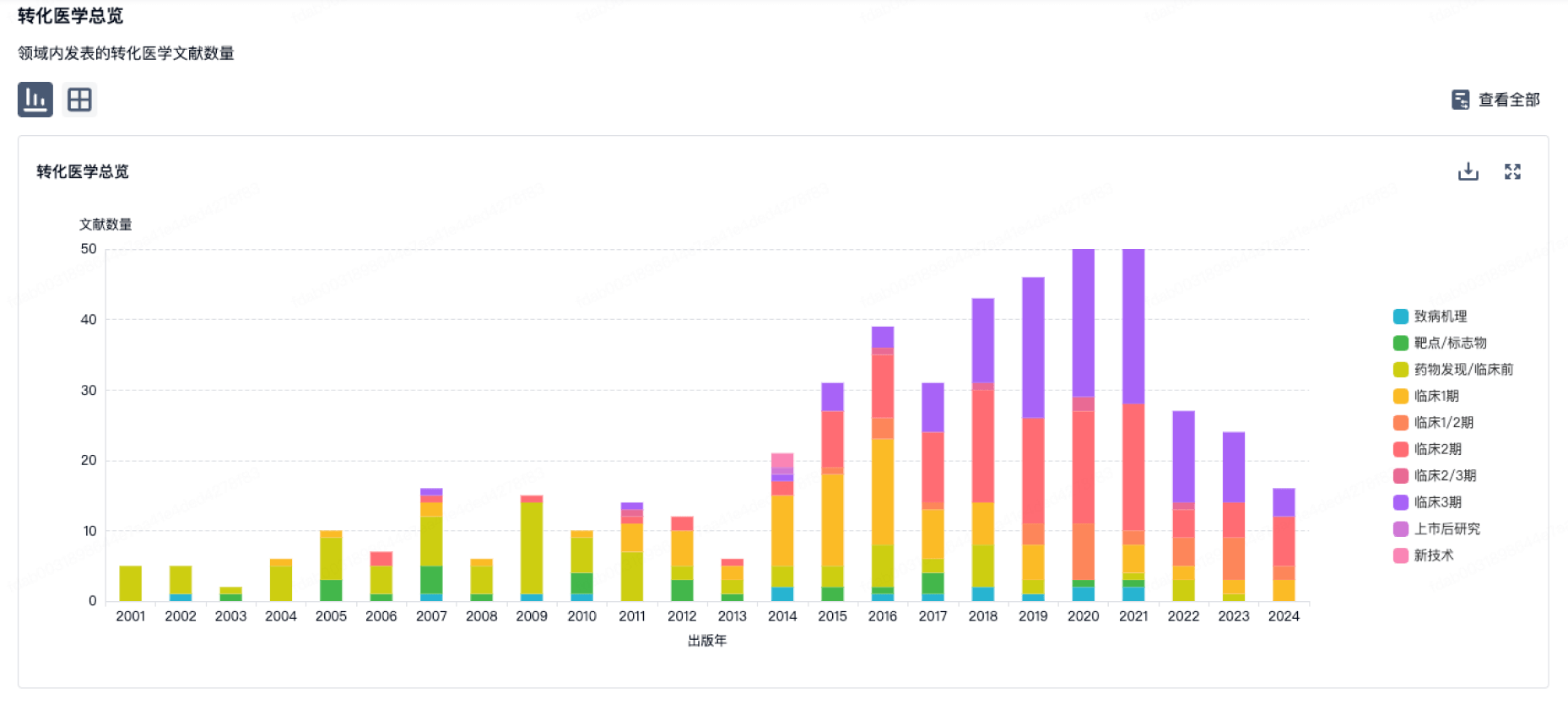
100 项与 Marcus Research Laboratory, Inc. 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2026年02月09日管线快照
无数据报导
登录后保持更新
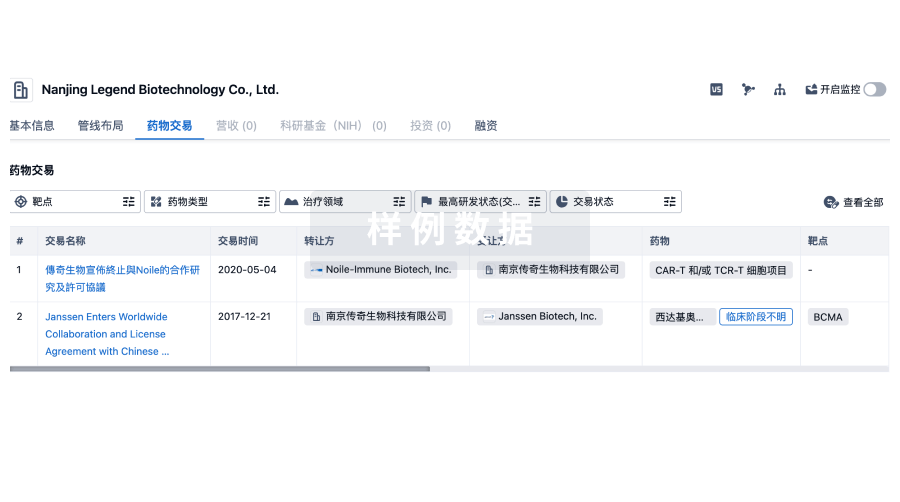
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
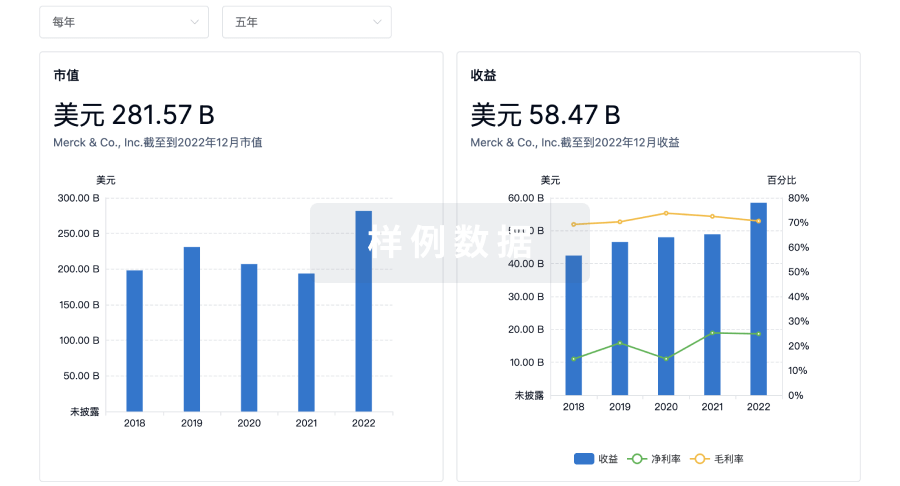
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
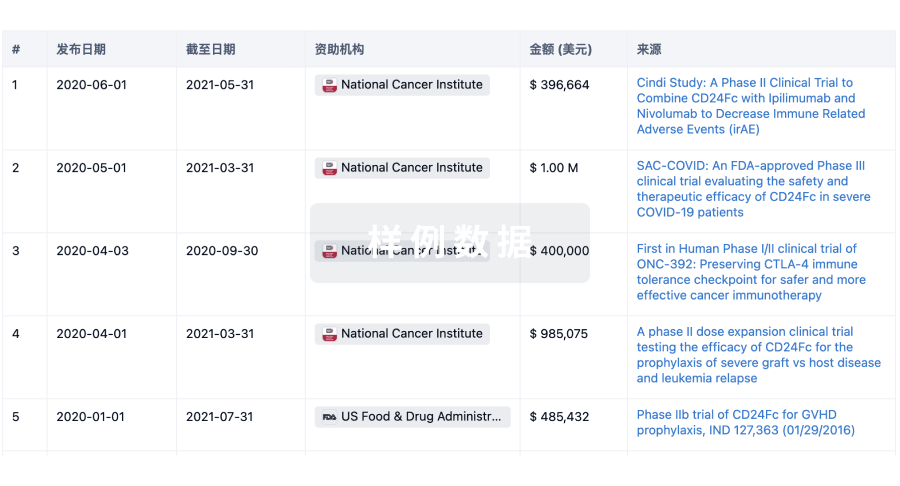
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
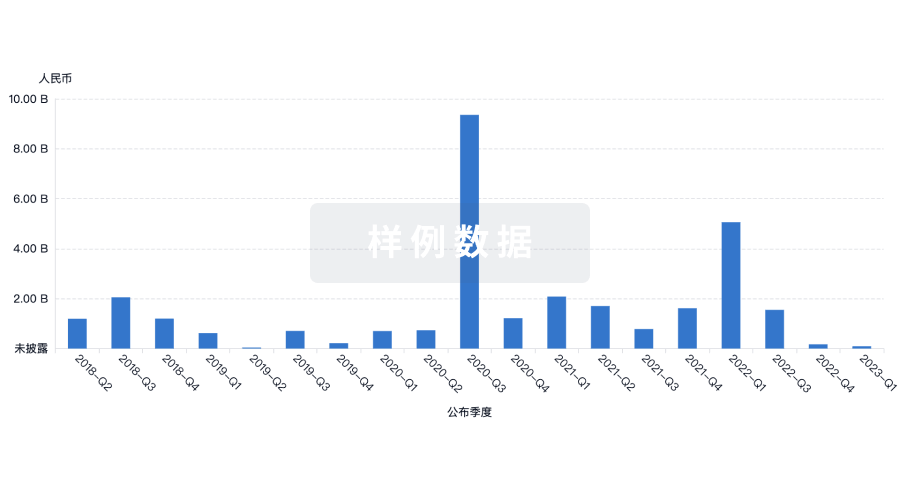
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
生物医药百科问答
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用