预约演示
更新于:2025-05-07
ACSL1
更新于:2025-05-07
基本信息
别名 ACS1、ACSL1、Acyl-CoA synthetase 1 + [16] |
简介 Catalyzes the conversion of long-chain fatty acids to their active form acyl-CoAs for both synthesis of cellular lipids, and degradation via beta-oxidation (PubMed:21242590, PubMed:22633490, PubMed:24269233). Preferentially uses palmitoleate, oleate and linoleate (PubMed:24269233). Preferentially activates arachidonate than epoxyeicosatrienoic acids (EETs) or hydroxyeicosatrienoic acids (HETEs) (By similarity). |
关联
100 项与 ACSL1 相关的临床结果
登录后查看更多信息
100 项与 ACSL1 相关的转化医学
登录后查看更多信息
0 项与 ACSL1 相关的专利(医药)
登录后查看更多信息
1,084
项与 ACSL1 相关的文献(医药)2025-06-01·Phytomedicine
Stevia rebaudiana root polysaccharide modulates liver metabolism, bile acid, and gut microbiota improving HFD-induced NAFLD: Potential roles of ACSL1 and FADS2
Article
作者: Wang, Guodong ; Wang, Jiapeng ; Bao, Yulong ; Lv, Qiuyue ; Liu, Chunyan ; Shang, Xiaolong ; Shao, Taili ; Che, Hui ; Hu, Guangdong ; Han, Jun
2025-06-01·Toxicology
Identification of key genes linking bisphenols exposure and breast cancer
Article
作者: Liu, Jiangzheng ; Yu, Weihua ; Li, Fei ; Liu, Rui ; Wang, Zhen ; Zhang, Xiaodi ; Gao, Tian ; Xu, Xinyue ; Cao, Meng ; Hai, Chunxu
2025-06-01·Molecular Immunology
Methotrexate loaded extracellular vesicles attenuate periodontitis by suppressing ACSL1 and promoting anti-inflammatory macrophage
Article
作者: Tao, Dihao ; He, Xiaoning ; Liu, Zhi ; Tan, Guodong ; Li, Bei ; Wang, Wenzhe ; Jin, Fang ; Yang, Jianhua ; Shi, Yuanyuan
14
项与 ACSL1 相关的新闻(医药)2024-12-30
·学术经纬
▎药明康德内容团队编辑
近期,我们邀请学术经纬的读者朋友评选了生物医学领域的年度突破。根据投票结果,2024年十大科学突破榜单正式发布。在这份榜单中,我们看到了基因疗法、靶向蛋白降解等创新疗法正在改变治疗格局;也看到了癌症、阿尔茨海默病等领域的重大机制突破……接下来,让我们共同揭晓本年度的科学突破榜单。
基因疗法恢复遗传性耳聋患者的听力
图片来源:123RF
遗传性耳聋影响全球高达2600万患者,在听力丧失的儿童中,超过60%源于遗传因素。现在,基因疗法的问世有望从根源上改善患者听力。
常染色体隐性遗传性耳聋9型(DFNB9)由编码耳铁蛋白的OTOF基因发生突变而导致,患者通常从出生起就有严重至极严重的双侧听力丧失。基因疗法AAV1-hOTOF使用腺相关病毒载体(AAV),将编码正常耳铁蛋白的基因递送到患者的内耳毛细胞。
今年1月,由复旦大学附属眼耳鼻喉科医院领衔的研究团队公布了全球首个OTOF基因疗法的临床试验结果,6名接受治疗的DFNB9患儿均未发现剂量限制性毒性和严重不良事件,其中5名患儿的听力得到显著改善,语言识别能力也获得显著提高,显示了基因疗法在遗传性耳聋临床治疗中的安全性和有效性。这项基因疗法的重要应用,也成为本次评选中得票数最高的科学突破。
CAR-T疗法“治愈”自身免疫疾病
今年2月,CAR-T疗法领域迎来了一项具有里程碑意义的研究结果。《新英格兰医学杂志》(NEJM)发表的一项论文显示,首批接受CAR-T疗法的15名患有严重自身免疫疾病的患者,在中位随访时间15个月期间持续维持缓解或症状大幅减轻,并已停止所有免疫抑制与抗炎药物的使用。入组的一名红斑狼疮患者在治疗后生活恢复正常、可以稳定行走,另外一名患有肌炎的患者从无法站立转变成了步行上班。这项“标志性”研究不仅让患者能够重新稳步前进,也代表着自身免疫领域治疗向前行进了一大步。
AlphaFold 3重磅登场
▲AlphaFold 3预测的复合物结构(图片来源:DOI: 10.1038/s41586-024-07487-w)
在2018年的国际蛋白质结构预测竞赛上,一个改变历史的“选手”AlphaFold诞生了;两年后,Google DeepMind研究团队进一步构建出了解决蛋白质折叠问题的新版本AlphaFold 2,但这仍不是它的极限。今年5月,DeepMind团队在《自然》杂志上宣布了AlphaFold 3的诞生,该模型能以前所未有的精度来预测所有生命分子的结构和相互作用。有了AlphaFold 3,我们将能够破解与人类息息相关的各种生物分子复合体结构,这项突破将蛋白质预测带到了一个全新高度。除了蛋白质-蛋白质相互作用,AlphaFold 3还能高效预测核酸分子、小分子、离子和修饰残基的位置、结构和互作,这让新型药物的设计具有了可能性。
首次证明,没有基因突变也会发生癌症
▲研究展示了通过降低Polycomb水平诱发癌症的例子(图片来源:Giacomo Cavalli)
几十年来,科学家认为抑癌基因或原癌基因的DNA序列产生变异往往会驱使细胞癌变。今年4月,法国国家科学研究中心(CNRS)和蒙彼利埃大学的研究者为癌变机制提出了新的见解。他们发表在《自然》杂志的研究显示,即便驱动基因的DNA没有发生突变,仅仅表观遗传上的变异——Polycomb蛋白的可逆损耗也足以导致细胞癌变。这是科学家们第一次证明基因突变并不是癌症发病的必要条件,该发现也让研究者开始重新思考癌症主要是基因疾病的理论。
HIV长效疗法:单次注射长期抑制病毒
▲TIP可以长期有效抑制HIV(图片来源:DOI: 10.1126/science.adn5866)
自发现以来,艾滋病(AIDS)一直是人类健康领域面临的重大挑战。今年,俄勒冈健康与科学大学与加州大学旧金山分校的研究者开发了一种人工合成的治疗性干扰颗粒(简称TIP),通过模拟有缺陷的HIV,TIP能有效压制体内的天然病毒。非人灵长类动物实验显示,单次TIP治疗即可在30周内持续抑制体内的HIV,将病毒水平降至千分之一。此外,长效预防HIV的暴露前预防(PrEP)疗法lenacapavir当选为《科学》杂志的年度科学突破。这些突破标志着艾滋病有望迎来长效预防与治疗的新时代。
找到阿尔茨海默病的隐藏“元凶”
▲脂质累积与AD病理发展相关(图片来源:DOI: 10.1038/s41586-024-07185-7)
阿尔茨海默病(AD)是神经科学领域的重大挑战,只有充分了解该疾病背后的更多机制,才能为AD的预防与治疗带来新希望。今年,斯坦福大学Tony Wyss-Coray教授团队发现了一种由脂滴相关酶ACSL1定义的小胶质细胞状态。AD高风险群体表达ACSL1的小胶质细胞数量更多,并且AD发生的遗传风险与小胶质细胞的脂滴累积存在联系。而在另一项研究中,加州大学伯克利分校的研究人员提出,细胞持续承受的应激压力是直接杀死神经元的“元凶”,因此帮助细胞关闭应激反应的药物或能帮助AD患者。这两项新研究为AD的治疗和干预带来了全新的靶点和方向。
干细胞再生疗法功能性治愈糖尿病
▲基于CiPS细胞治疗1型糖尿病的临床研究示意图(图片来源:DOI: 10.1016/j.cell.2024.09.004)
利用多能干细胞制备的胰岛细胞,为糖尿病提供了新的移植来源。天津市第一中心医院与北京大学、昌平实验室的合作团队利用化学重编程的方法将人成体细胞诱导为多能干细胞(简称CiPS细胞),再制备出功能成熟的胰岛细胞,用于治疗1型糖尿病。在今年的《细胞》杂志上,研究团队发表了首个临床研究结果,首例接受移植的患者已恢复内源自主性、生理性的血糖调控,移植75天后完全稳定脱离胰岛素注射治疗,实现了临床功能性治愈。作者指出,化学重编程有望高效制备各种功能细胞,为细胞治疗在重大疾病治疗中的广泛应用开辟新的道路。
iPSC移植恢复人类视力
▲诱导人类多能干细胞的制备与移植示意图(图片来源:DOI: 10.1016/S0140-6736(24)01764-1)
角膜是保护我们眼睛的重要结构,而作为角膜的生产者,角膜缘干细胞一旦缺失或出现功能障碍,就可能引起视力下降甚至是失明。利用诱导多能干细胞(iPSC)分化的角膜上皮片层,日本大阪大学的研究团队成功实现了角膜缘干细胞缺乏症的临床移植试验。在今年的《柳叶刀》杂志上,该团队发表了4名接受iPSC移植患者的随访情况。所有患者治疗眼的疾病分期均有所改善,矫正远视力增强,角膜混浊情况改善。论文指出,这是首次使用iPSC分化的角膜上皮细胞,用于修复因角膜缘干细胞缺乏症而视力受损患者的角膜。
靶向蛋白降解剂攻克“不可成药”靶点
靶向蛋白降解技术近年来备受关注,其能够针对传统小分子或抗体药物难以靶向的蛋白发挥作用,让曾经的“不可成药”靶点变得可及。今年,基于靶向蛋白降解,来自勃林格殷格翰(Boehringer Ingelheim)、英国邓迪大学的合作团队向长期被视作“不可成药靶点”的KRAS蛋白发起挑战,他们设计了一种全新的蛋白降解靶向嵌合体(Proteolysis-Targeting Chimeras,简称PROTAC)。在17种最常见的KRAS突变体中,这款全新PROTAC分子可降解并灭活其中的13种突变体,有望带来通用型癌症疗法。
此外,Dana-Farber癌症研究中心与哈佛大学医学院的科学家设计了新型靶向蛋白降解技术——转铁蛋白受体靶向嵌合体(TransTAC)。TransTAC在体外细胞实验中可高效降解EGFR、PD-L1、CD20和嵌合抗原受体(CAR)等多种膜蛋白,在癌症等疾病治疗中展现出巨大潜力。
大脑高清细节图来了
▲研究从一个立方毫米大小的脑组织中还原了高分辨率的细节图(图片来源:DOI: 10.1126/science.adk4858)
看清大脑的微观细节,成为脑科学研究者一直追寻的目标。在今年的一篇《科学》论文中,哈佛大学与Google的研究者合作绘制了一幅大脑皮层高分辨率细节图,利用来自手术切除的一立方毫米脑组织揭开大脑的微观世界。图谱展示出了颞叶皮层中每个细胞以及神经元之间的连接网络,还有那些过往从未描述过的颞叶皮层细节,例如其中胶质细胞的数量明显超过了神经元、含有约50个突触连接的特殊轴突。下一步,研究团队希望利用该技术创建完整的大鼠大脑高分辨率图谱。
本文来自药明康德内容微信团队,欢迎转发到朋友圈,谢绝转载到其他平台。如有开设白名单需求,请在“学术经纬”公众号主页回复“转载”获取转载须知。其他合作需求,请联系wuxi_media@wuxiapptec.com。
免责声明:药明康德内容团队专注介绍全球生物医药健康研究进展。本文仅作信息交流之目的,文中观点不代表药明康德立场,亦不代表药明康德支持或反对文中观点。本文也不是治疗方案推荐。如需获得治疗方案指导,请前往正规医院就诊。
点个“在看”再走吧~
细胞疗法基因疗法免疫疗法临床研究
2024-10-08
·梅斯医学
“早上困,中午困,下午困,晚上一拿手机就不困”“熬最长的夜,敷最贵的面膜”“三点睡五点起,阎王夸我好身体”......成了不少年轻人/打工人的真实写照。睡眠的重要性不言而喻,但随着社会飞速发展以及生活节奏加快,“拼命十三郎”们只能舍弃睡眠时间,转而投入996甚至007的“社畜”工作中。
除了繁忙的工作压缩了睡眠时间外,一个更常见的现象是“报复性熬夜”——白天被各种琐事儿缠身,只有当晚上躺在床上的那一刻,时间才开始属于自己,接着就是“无止境”的熬夜。当从手机中“缓过神”的时候,发现自己根本睡不了几个小时又要搬砖了!
今年最新发布的《2024中国居民睡眠健康白皮书》显示,我国居民的平均睡眠时长为6.75小时,甚至有28%的人夜间睡眠时长不足6小时。睡不着、睡不深、睡不够等问题困扰着越来越多的人。
正如大家调侃得那样“晚上不睡,白天崩溃”,崩溃的不仅是精神,还有身体的方方面面——熬夜使得主要心血管不良事件的风险提升92%;增加海马体中抑制神经元活动,破坏记忆巩固;导致癌症的发生等等。早在2007年,国际癌症研究机构(IARC)就把熬夜(包括昼夜节律打乱的轮班工作)定义为2A类致癌物。
近日,来自大连医科大学的研究团队揭示了睡眠不足成为癌症“铺路石”的详细机制——具体来说,当机体睡眠不足时,脂肪酸氧化能够感知到昼夜节律的紊乱,并进一步传递到致癌代谢信号,增强癌症干性和肿瘤发生。但好消息是,有改善方法!
DOI: 10.1016/j.cmet.2024.04.018
“睡眠不足会扰乱昼夜节律”并不是新闻,睡眠不足(SD)会引发多种病理过程,包括:免疫功能紊乱、新陈代谢失调,以及过早死亡。而这种昼夜节律的紊乱同样会作用于癌症发展过程,比如:改变葡萄糖、谷氨酸、胆汁酸和脂质代谢等诸多代谢,进而刺激肿瘤生长、转移和免疫逃逸。但其中的具体机制仍不清楚,也是本次研究的焦点。
第一步,研究者定位到生物机体内感知昼夜节律的关键一环——脂肪酸氧化(FAO)。
具体来说,研究者首先发现,与对照组小鼠相比,SD组有着更长的清醒时间,以及更短的非快速动眼(NREM)和快速动眼(REM)睡眠。由此可见,睡眠不足确实会导致小鼠昼夜节律发生紊乱。
紧接着,研究者分析了野生型C57BL/6J小鼠在SD和对照组培养4周后肺组织中的转录组和代谢组的差异。RNA测序结果显示,3284个基因在SD之后失去了昼夜振荡,而这些基因与脂质代谢的生物过程高度相关,尤其是FAO。
FAO是指,利用脂肪酸转化成的乙酰辅酶A(CoA)亚基来进一步生成烟酰胺腺嘌呤二核苷酸(NADH)、乙酰-CoA和三磷酸腺苷(ATP),参与到产生能量、氧化还原平衡和生物合成反应的过程中。事实上,与FAO相关的代谢酶和代谢物表现出生物节律,以维持生理功能的平衡。
对比SD组和对照组发现,对照组的FAO基因显示出节律模式,但SD组的FAO基因则出现了明显的紊乱。同时,进一步的超高效液相色谱/质谱分析显示,SD小鼠肺组织中振荡脂质代谢物的数量明显减少,并找到了47种节律紊乱的脂质代谢物,其中游离脂肪酸(FFAs)占比最大。
由此可见,FAO能在基因表达和代谢功能水平上感知到SD诱导的生理状态节律紊乱。
FAO能感知到SD情况下的昼夜节律紊乱
第二步,找到FAO在SD促癌过程中担任的“角色”。
与对照组对比,SD明显加速了肺癌的发生和发展,包括促进肺肿瘤的形成、提高Ki-67和CD166的蛋白水平,同时还会在mRNA和蛋白水平上增加MYC、SOX2、CD166和NANOG的表达。
类似的情况也发生在患者源性异种移植模型中,SD导致肿瘤体积明显增加,降低肿瘤小鼠的存活率,并提高了肿瘤组织中干性标记物的表达。
当研究者对肺肿瘤进行RNA-seq分析后发现,SD显著上调了827个基因,而这些上调的基因主要属于脂肪酸代谢过程,尤其是FAO。同时,SD组的肿瘤组织中FAO基因表达量明显高于对照组的,并表现出了节律紊乱。
为了进一步确认FAO在整个过程中的重要作用,研究者给小鼠注射了FAO抑制剂依托莫西(ETO)。结果并不令人意外,ETO逆转了SD增强的肿瘤发生,还阻碍了肺癌中Ki-67、CD166、乙酰-CoA和ATP水平的升高,即ETO逆转了SD增加的肿瘤形成率。
总结来说,FAO感知昼夜节律紊乱是SD增强肺癌干性的基础;相反,当FAO受到抑制,肿瘤进展也会被阻碍。
FAO昼夜节律紊乱维持SD诱导的肺肿瘤病变
第三步,详细定位到FAO节律紊乱维持SD诱导肺癌干性过程中的关键分子。
对比两组情况,研究者定位到13个可能导致SD引起的致癌FAO节律紊乱的常见基因。进一步,在这13个基因中发现,Acsl1 mRNA水平在SD处理的肺肿瘤中显著升高;类似的情况还出现在患者源性异种移植模型里,ACSL1蛋白表达同样有所增加。
那么,ACSL1是不是我们要找的关键分子呢?当研究者敲除了Ascl1之后,SD促进肺癌进展的作用被阻断了,Ki-67、CD166和ACSL1的表达也趋于平常,再一次证实了ACSL1是SD增强肺肿瘤发生和发展的必要条件。
于是,研究者进一步的分析了“可能作为ACSL1底物和产物的长链FAO代谢物”。鉴于棕榈酸(PA)是最主要的饱和脂肪酸,也是长链饱和或不饱和脂肪酸的前体,研究者将目光投向了PA及其代谢产物PA-CoA。
ACSL1具有将PA转化为PA-CoA的能力,而PA-CoA将以棕榈酰肉碱的形式进一步转运到线粒体中,以促进FAO的过程。而本实验结果表明,睡眠不足会扰乱了ACSL1的节律使得更多的PA转化为PA-CoA,从而促进FAO,导致肺癌生长的加速和干性特征的维持。
除此之外,ACSL1还能感知到SD诱导的CLOCK节律失常,同样能促进肺癌的发生。
睡眠不足致癌的信号通路
当然,好消息是,研究者找到了解“癌”方案——β-内啡肽。
在14种与睡眠相关的激素中,研究者发现,注射了β-内啡肽能够显著下调肺癌细胞系中的CLOCK和ACSL1 mRNA的表达,而连续给药甚至能逆转SD诱导的肿瘤生长。
为了评估上述发现的临床价值,他们还开展了前瞻性和回顾性的临床研究,评估了血清β-内啡肽水平、睡眠质量与肺癌发展之间的临床相关性。结果令人“鼓舞”,血清β-内啡肽水平与深度睡眠和REM的比例呈正相关,而与清醒次数呈负相关,即β-内啡肽水平能够很好地预测睡眠质量。
不仅如此,对62例肺癌患者开展的回顾性分析显示,β-内啡肽水平与CLOCK、ACSL1、CD166和Ki-67评分呈负相关,呈现出抗肿瘤效果;而生存期分析结果表明,血清中的高β-内啡肽水平具有较好的预后价值。
临床试验中β-内啡肽水平与指标之间的影响
综上所述,睡眠不足会促进癌症的生长、转移和免疫逃逸,而在这个过程中,FAO的节律紊乱发挥了重要的作用。不过好消息是,补充β-内啡肽或能有效地规避这一风险。
因此,“睡眠不足,癌症来找”是有科学依据的。睡眠的重要性不言而喻,不如就从今天开始早睡计划吧!
参考资料:
[1]Peng F, Lu J, Su K, Liu X, Luo H, He B, Wang C, Zhang X, An F, Lv D, Luo Y, Su Q, Jiang T, Deng Z, He B, Xu L, Guo T, Xiang J, Gu C, Wang L, Xu G, Xu Y, Li M, Kelley KW, Cui B, Liu Q. Oncogenic fatty acid oxidation senses circadian disruption in sleep-deficiency-enhanced tumorigenesis. Cell Metab. 2024 May 14:S1550-4131(24)00138-4. doi: 10.1016/j.cmet.2024.04.018. Epub ahead of print. PMID: 38772364.
撰文 | Swagpp
编辑 | Swagpp
● 重磅!科主任训话:卫健委鼓励医护兼职创业,不是让你去摆摊、送外卖!要和医疗相关!上了热搜后,他从医院离职了!医护兼职到底能干啥?
● 阿司匹林再添新用!JAMA最新:仅低剂量服用,能使肝脏脂肪减少10.2%、糖尿病风险降15%!
● 热议!阳光绩效实行,医生工资升,护士工资降?护士透露:从9千多降到2千多,不怕本科护士全跑光?护士有400万缺口,不能厚此薄彼
版权说明:梅斯医学(MedSci)是国内领先的医学科研与学术服务平台,致力于医疗质量的改进,为临床实践提供智慧、精准的决策支持,让医生与患者受益。欢迎个人转发至朋友圈,谢绝媒体或机构未经授权以任何形式转载至其他平台。
点击下方「阅读原文」 立刻下载梅斯医学APP!
2024-07-30
PHILADELPHIA, July 30, 2024 (GLOBE NEWSWIRE) -- ADvantage Therapeutics today announced groundbreaking results from a study on AD04™, a novel therapeutic candidate for Alzheimer's disease. Scientists from ADvantage presented the research poster "AD04™ - modifying Alzheimer’s disease by modulation of hippocampal lipid metabolism" at the Alzheimer’s Association International Conference, Key Findings: Pleiotropic Activity: Clinical and proteomics-based studies demonstrated that AD04™ exhibits durable and multifaceted effects, including both immediate symptomatic relief and long-term disease modification.Safety Profile: AD04™ boasts an excellent safety profile in previous Phase 2 studies, notably not triggering ARIA-E/H, a common adverse event associated with other Alzheimer's treatments.Mechanism of Action: Treatment with AD04™ significantly restored the expression of proteins involved in lipid metabolism in aged mice, aligning their levels with those of young mice. This restoration is believed to contribute to the therapeutic effects observed.Lipid Metabolism: Key proteins in lipid metabolism, such as ACSL1, were found to be upregulated after AD04™ treatment. This suggests that the therapeutic benefits of AD04™ are closely tied to the modulation of hippocampal lipid metabolism. Clinical Implications: Improved Cognitive Functions: The ability of AD04™ to modulate lipid metabolism and restore protein expression points to potential improvements in cognitive function and overall brain health in Alzheimer's patients.Novel Class of AD Drugs: AD04™ could represent the first of a new class of Alzheimer’s disease drugs, distinguished by its unique mechanism of action and safety profile. "The significant and multifaceted benefits observed with AD04™ treatment mark a pivotal step forward in Alzheimer's research,” said Dr. Carmela R. Abraham, CSO of ADvantage. “We are optimistic that AD04™ may offer a new lifeline to patients and their families, providing both immediate symptomatic relief and modification of the disease progression." The research has been well-received by the scientific community, with anticipation building around the upcoming clinical trials. ADvantage Therapeutics is committed to advancing this promising candidate through rigorous testing and regulatory review to bring a novel, effective, and safe treatment option to those affected by Alzheimer’s disease. About AD04™ ADvantage Therapeutics is developing AD04™ as a new class of therapeutic for mild Alzheimer’s disease that exhibits durable multifaceted effects, including both immediate symptomatic relief and long-term disease-modification. The compound has been used extensively as an adjuvant in human and animal vaccination programs. In a previous trial, AD04™ serving as a control against another compound appeared to demonstrate statistically significant slower decline in cognitive and quality of life clinical measures compared to other treatment groups. AD04™ also showed slower decline in hippocampal volume as a biomarker. The Company believes that rather than being limited to a specific aspect of AD pathology, such as amyloid beta or tau, AD04™ may restore expression of genes in lipid metabolism, improve phagocytosis and reduce inflammation. Through multiple mechanisms AD04™ functions as an immunomodulator, stimulating and/or regulating the immune system to reduce AD pathology. About Alzheimer’s Disease About fifty million people worldwide suffer from Alzheimer’s Disease, which is the sixth leading cause of death in industrialized countries. In 2019, the World Health Organization estimated the total worldwide cost of dementia at $1.3 trillion and expects this cost to rise to $2 trillion by 2030. The socio-economic burden of Alzheimer’s Disease is enormous. AD devastates the lives of patients and their families. AD victims lose their memory and independence. Alzheimer’s Disease is a high unmet medical need as there are currently no disease-modifying drugs approved worldwide. The availability of a safe, effective, affordable drug would transform the life of an AD patient from accepting a debilitating disease to the retention of personality, independence, and dignity. About ADvantage Therapeutics, Inc. Headquartered in the Wynwood neighborhood in Miami, ADvantage Therapeutics is developing therapies to treat neurodegenerative conditions with a central focus on Alzheimer’s Disease. The Company’s lead compound AD04™ is a subcutaneous injectable therapy that entered, in November 2023, confirmatory Phase 2b clinical trials in Europe, to evaluate its safety and efficacy in early AD patients. The Company believes that AD04™ may function as an immunomodulator, stimulating and regulating the immune system to reduce AD pathology, rather than limiting therapy to attack the misfolded proteins, beta amyloid and tau. The Company is exploring additional approaches to mitigating neurodegenerative disease, among them increasing the levels of Klotho, a life and health extending protein which will have an overall impact on longevity. About ADvantage Therapeutics GmbH ADvantage Therapeutics GmbH, founded in 2021, is the Vienna BioCenter-based subsidiary of ADvantage Therapeutics, Inc., where the Company conducts early research and drug development. Safe Harbor-Forward-Looking Statements This press release may contain forward-looking statements, including statements of potential mode of action, potential clinical effect, potential safety, and ADvantage’s potential clinical development program and pipeline program. ADvantage is in the early stages of developing and testing its AD04™ compound and may not receive future regulatory approvals needed for marketing it as a drug. The described clinical effect of our lead compound AD04™ is primarily based on results of a Phase 2 study designed to evaluate a different compound. The described results need to be confirmed for proof of concept, might not be representative of larger scale clinical trials, and do not guarantee future clinical success. Any preclinical results presented here are interim. The mechanism of action of AD04™, as potentially determined in our future investigations, particularly in future clinical trials with patients with Alzheimer’s Disease, might differ from the one presented. For more information, please contact: Dr. Carmela R. Abraham, CSOADvantage Therapeutics, Inc.195 NW 40th StreetMiami, FL 33127Phone: +1-781-424-7090Email: Carmela@advantagetherapeutics.com Disclaimer: The views and opinions expressed herein do not reflect those of the Alzheimer's Association.
临床2期
分析
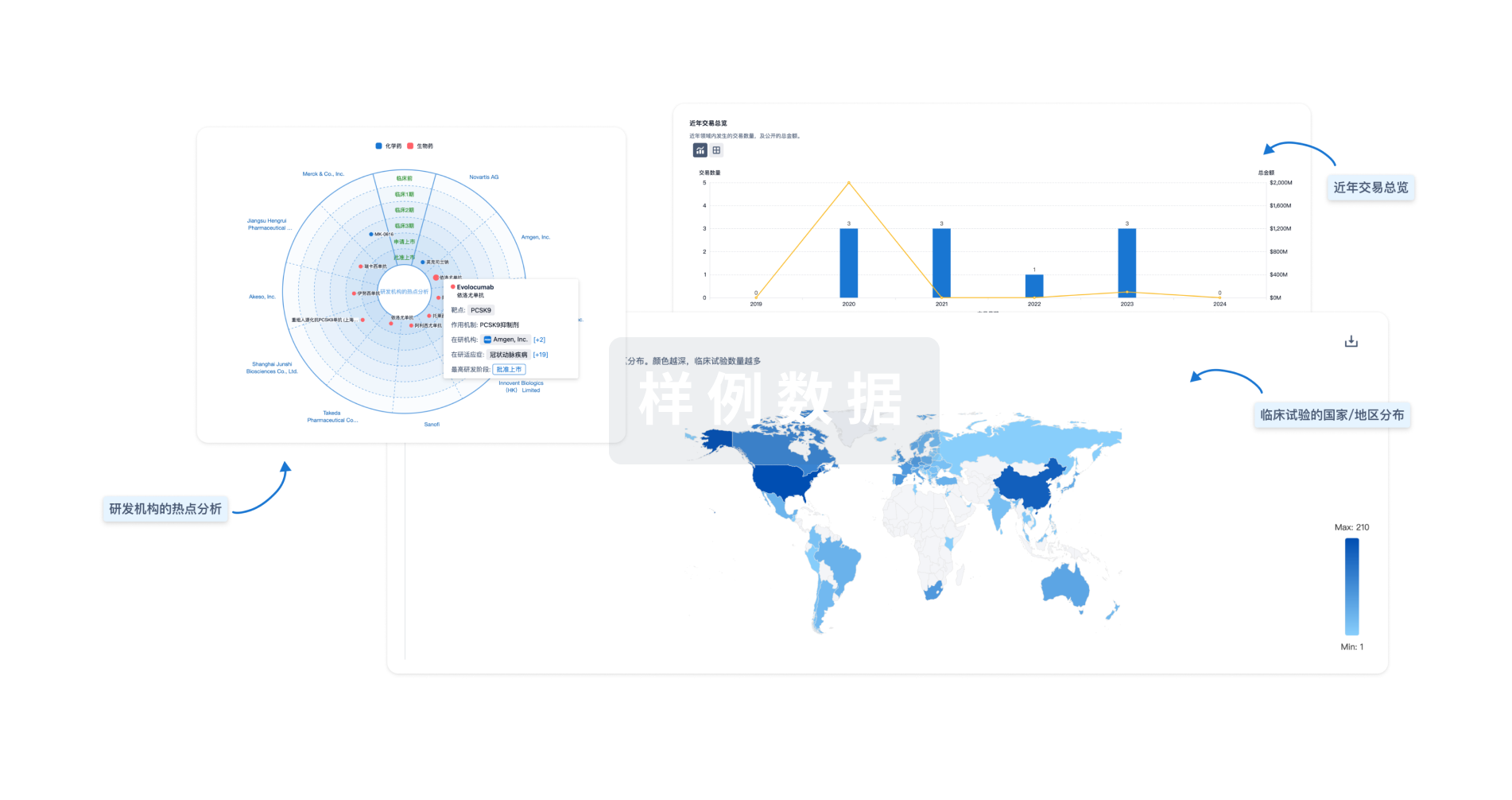
对领域进行一次全面的分析。
登录
或
生物医药百科问答
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用