预约演示
更新于:2026-01-17
Alatrofloxacin Mesylate
甲磺酸阿曲沙星
更新于:2026-01-17
概要
基本信息
药物类型 小分子化药 |
别名 Alatrofloxacin mesylate (USAN)、L-Ala-L-Ala-CP-99219、517-27 + [6] |
作用方式 抑制剂 |
作用机制 Bacterial DNA gyrase 抑制剂(细菌DNA旋转酶 抑制剂)、Bacterial top IV抑制剂(细菌拓扑异构酶IV抑制剂) |
治疗领域 |
在研适应症- |
非在研适应症 |
原研机构 |
在研机构- |
权益机构- |
最高研发阶段撤市 |
首次获批日期 美国 (1997-12-18), |
最高研发阶段(中国)- |
特殊审评- |

登录后查看时间轴
结构/序列
分子式C27H29F3N6O8S |
InChIKeyCYETUYYEVKNSHZ-LGOOQLFJSA-N |
CAS号146961-77-5 |
关联
100 项与 甲磺酸阿曲沙星 相关的临床结果
登录后查看更多信息
100 项与 甲磺酸阿曲沙星 相关的转化医学
登录后查看更多信息
100 项与 甲磺酸阿曲沙星 相关的专利(医药)
登录后查看更多信息
54
项与 甲磺酸阿曲沙星 相关的文献(医药)2022-11-24·Journal of biomolecular structure & dynamics
Virtual screening of quinoline derived library for SARS-COV-2 targeting viral entry and replication
Article
作者: Hussain, Firasat ; Pant, Pradeep ; Anju, Anju ; Mishra, Anil Kumar ; Chaturvedi, Shubhra ; Chaudhary, Vishakha
The COVID-19 pandemic infection has claimed many lives and added to the social, economic, and psychological distress. The contagious disease has quickly spread to almost 218 countries and territories following the regional outbreak in China. As the number of infected populations increases exponentially, there is a pressing demand for anti-COVID drugs and vaccines. Virtual screening provides possible leads while extensively cutting down the time and resources required for ab-initio drug design. We report structure-based virtual screening of a hundred plus library of quinoline drugs with established antiviral, antimalarial, antibiotic or kinase inhibitor activity. In this study, targets having a role in viral entry, viral assembly, and viral replication have been selected. The targets include: 1) RBD of receptor-binding domain spike protein S 2) Mpro Chymotrypsin main protease 3) Ppro Papain protease 4) RNA binding domain of Nucleocapsid Protein, and 5) RNA Dependent RNA polymerase from SARS-COV-2. An in-depth analysis of the interactions and G-score compared to the controls like hydroxyquinoline and remdesivir has been presented. The salient results are (1) higher scoring of antivirals as potential drugs (2) potential of afatinib by scoring as better inhibitor, and (3) biological explanation of the potency of afatinib. Further MD simulations and MM-PBSA calculations showed that afatinib works best to interfere with the the activity of RNA dependent RNA polymerase of SARS-COV-2, thereby inhibiting replication process of single stranded RNA virus. Communicated by Ramaswamy H. Sarma.
2021-07-01·Biochemistry and biophysics reports
Repurposing of approved drugs with potential to interact with SARS-CoV-2 receptor
Article
作者: Sajib, Abu Ashfaqur ; Ahsan, Tamim
Respiratory transmission is the primary route of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) infection. Angiotensin I converting enzyme 2 (ACE2) is the known receptor of SARS-CoV-2 surface spike glycoprotein for entry into human cells. A recent study reported absent to low expression of ACE2 in a variety of human lung epithelial cell samples. Three bioprojects (PRJEB4337, PRJNA270632 and PRJNA280600) invariably found abundant expression of ACE1 (a homolog of ACE2 and also known as ACE) in human lungs compared to very low expression of ACE2. In fact, ACE1 has a wider and more abundant tissue distribution compared to ACE2. Although it is not obvious from the primary sequence alignment of ACE1 and ACE2, comparison of X-ray crystallographic structures show striking similarities in the regions of the peptidase domains (PD) of these proteins, which is known (for ACE2) to interact with the receptor binding domain (RBD) of the SARS-CoV-2 spike protein. Critical amino acids in ACE2 that mediate interaction with the viral spike protein are present and organized in the same order in the PD of ACE1. In silico analysis predicts comparable interaction of SARS-CoV-2 spike protein with ACE1 and ACE2. In addition, this study predicts from a list of 1263 already approved drugs that may interact with ACE2 and/or ACE1 and potentially interfere with the entry of SARS-CoV-2 inside the host cells.
2021-01-01·Computational and Structural Biotechnology Journal
Prediction of repurposed drugs for Coronaviruses using artificial intelligence and machine learning
Article
作者: Kumar, Manoj ; Rajput, Akanksha ; Thakur, Anamika ; Mukhopadhyay, Adhip ; Gautam, Sakshi ; Jassal, Harvinder ; Kamboj, Sakshi ; Rastogi, Amber
The world is facing the COVID-19 pandemic caused by Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2). Likewise, other viruses of the Coronaviridae family were responsible for causing epidemics earlier. To tackle these viruses, there is a lack of approved antiviral drugs. Therefore, we have developed robust computational methods to predict the repurposed drugs using machine learning techniques namely Support Vector Machine, Random Forest, k-Nearest Neighbour, Artificial Neural Network, and Deep Learning. We used the experimentally validated drugs/chemicals with anticorona activity (IC50/EC50) from 'DrugRepV' repository. The unique entries of SARS-CoV-2 (142), SARS (221), MERS (123), and overall Coronaviruses (414) were subdivided into the training/testing and independent validation datasets, followed by the extraction of chemical/structural descriptors and fingerprints (17968). The highly relevant features were filtered using the recursive feature selection algorithm. The selected chemical descriptors were used to develop prediction models with Pearson's correlation coefficients ranging from 0.60 to 0.90 on training/testing. The robustness of the predictive models was further ensured using external independent validation datasets, decoy datasets, applicability domain, and chemical analyses. The developed models were used to predict promising repurposed drug candidates against coronaviruses after scanning the DrugBank. Top predicted molecules for SARS-CoV-2 were further validated by molecular docking against the spike protein complex with ACE receptor. We found potential repurposed drugs namely Verteporfin, Alatrofloxacin, Metergoline, Rescinnamine, Leuprolide, and Telotristat ethyl with high binding affinity. These 'anticorona' computational models would assist in antiviral drug discovery against SARS-CoV-2 and other Coronaviruses.
1
项与 甲磺酸阿曲沙星 相关的新闻(医药)2024-11-27
·药学进展
“
点击蓝字 关注我们
孙丽新
药理学博士,中国药科大学副研究员,硕士生导师。主要从事分子药理学和药物毒理学研究。主要研究方向为:药源性肝肠损伤及肝脏、肠道疾病的分子机制研究;药物对肝脏毒性作用机制和物质基础研究;炎症性肠病的分子机制研究及内源性代谢物对肠道免疫的影响。曾主持国家自然科学基金项目 4 项、江苏省自然基金项目 1 项,发表 SCI 收录论文数十篇。
中性粒细胞在药物性肝损伤中的作用研究进展 PPS
文晴 1,杨梦娇 1,原玉柱 1,赵永睿 1,肖莉 2,孙丽新 1*
(1. 中国药科大学多靶标天然药物全国重点实验室新药筛选与药效评价中心,江苏 南京 210009;2. 湖州师范学院生命科学学院,浙江 湖州 313000)
[ 摘要 ] 在药物性肝损伤(DILI)的疾病发展过程中,被募集到肝脏中的中性粒细胞通过多种途径加重肝损伤的作用受到广泛关注。然而,中性粒细胞在不同种类药物导致的直接型、间接型、特异质型 DILI 中的作用机制不同,尚待进一步研究。综述中性粒细胞在 DILI 中发挥加重肝损伤作用以及中性粒细胞分别在乙酰氨基酚、雷公藤甲素和吡咯里西啶生物碱导致的直接型 DILI,氟烷诱导的特异质型 DILI,免疫检查点抑制剂引起的间接型 DILI 中的作用机制,为探究 DILI 的内在机制和治疗手段提供参考。
药物性肝损伤(drug-induced liver injury,DILI)是一种严重的药物不良反应,轻症可致血清转氨酶浓度升高,重症可致肝衰竭甚至死亡 [1]。DILI 是临床上急性肝衰竭的常见病因,引起人们广泛关注。然而,DILI 内在机制复杂,目前尚未被完全阐明,仍需大量研究。研究表明,多种药物导致的 DILI 中,均存在中性粒细胞在肝脏中蓄积和活化的现象。中性粒细胞在 DILI 的疾病进程中起重要作用。本文重点阐述中性粒细胞在 DILI 中的作用,探讨中性粒细胞作为 DILI 治疗靶点的潜在作用。
1
药物性肝损伤概述
DILI 是一种常见药物不良反应,是多种新药开发受阻和上市药物撤市的原因。导致 DILI 的药物主要包括化学药物、中草药和膳食补充剂。导致 DILI的药物类别存在地区差异。在欧美国家,膳食补充剂导致的肝损伤病例逐渐增多,但其所占比重较小,以抗生素为首的化学药物仍是导致 DILI 的主要药物 [2-5]。在亚洲地区,尤其是东亚地区,中草药导致的 DILI 所占比重较大 [6-8]。在中国人群中导致 DILI的主要药物为中草药,占比为 26.81%,其次是抗结核药物,占比为 21.99%,而抗肿瘤药物、抗感染药物、抗精神病药物的占比均不超过 10%[9]。
DILI 表现为肝功能障碍,可能伴有黄疸,严重时可致急性肝衰竭甚至死亡 [10-12]。根据药物导致肝损伤的不同方式,DILI 可分为直接型 DILI、特异质型 DILI 和间接型 DILI[10]。其中,直接型 DILI 由具有肝毒性的药物直接引起,具有剂量依赖性、重复性和可预测性。引起特异质型 DILI 的药物一般只有极小的直接毒性或无直接毒性。特异质型 DILI 具有不可预测性,动物实验难以复制,个体差异显著,通常与药物剂量无相关性。间接型 DILI 与以上 2 种不同,继发于药物的药理作用,而不是由药物固有的毒性或免疫原性引起,表现为诱发或加重肝脏疾病 [11,13]。
2
中性粒细胞对药物性肝损伤的调节过程
2.1 中性粒细胞的分化和成熟
中性粒细胞是哺乳动物体内数量最多的白细胞,具有强大的抗菌作用,是体内抵御病原体入侵的第一道防线 [14]。然而,中性粒细胞的抗菌活性也使其具有生物毒性,可导致组织损伤和自身免疫性疾病 [15-17]。成熟中性粒细胞以约 1 011 个/d 的速率在骨髓中生成,但在血管内停留的时间平均只有 6 ~8 h。当病原体感染机体时,骨髓中的中性粒细胞将加速生成 [15]。成熟中性粒细胞的形成过程分为 2 个阶段,即分化和成熟。其中,分化是指造血干细胞分化和发育为髓系祖细胞,再分化为中性粒细胞前体的过程。成熟则是指中性粒细胞前体依次生成初级颗粒、次级颗粒、三级颗粒和分泌囊泡,并形成分叶核,成为成熟中性粒细胞的过程 [18-19]。
2.2 中性粒细胞的募集
循环中的中性粒细胞受到损伤相关分子模式(damage-associated molecular patterns,DAMPs)、促炎因子和趋化因子等因素诱导,向肝脏趋化,黏附在内皮上,通过迁移抵达损伤部位,这一过程被称为中性粒细胞的募集(见图 1)。当中性粒细胞被招募时,其在骨髓中的分化和生成速度也加快。在DILI 中,肝细胞释放 DAMPs,如热休克蛋白 60(heat shock protein 60,HSP60)、 高 迁 移 率 族 蛋 白 1(high mobility group proteins,HMGB1)和腺嘌呤核苷三磷酸(adenosine triphosphate, ATP)等,与相应受体结合,介导中性粒细胞的趋化 [20-21]。肝脏中的枯否细胞(Kupffer cell,KC)释放肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)、白介素-1β(interleukin-1β,IL-1β)等促炎因子,诱导中性粒细胞的趋化。同时,CXC 趋化因子配体 1(C-X-C chemokine ligand 1,CXCL1)、CXC 趋化因子配体2/8(C-X-C chemokine ligand 2/8,CXCL2/8)与 CXC趋化因子受体 2(C-X-C chemokine receptor type 2,CXCR2)结合,甲酰肽与甲酰肽受体 1(formyl peptide receptor 1,FPR1)结合,协同招募中性粒细胞至肝坏死部位 [22]。中性粒细胞的黏附和迁移过程由中性粒细胞表达的 β2 整合素(integrin β2,ITGβ2)和内皮细胞表达的黏附因子 [ 细胞间黏附分子 -1(intercellular adhesion molecules-1,ICAM-1)、血管细胞黏附分子 -1(vascular cell adhesion molecule-1,VCAM-1)、血小板内皮细胞黏附分子 -1(platelet endothelial cell adhesion molecule-1,PECAM-1)] 共同调控 [23]。
2.3 中性粒细胞加重肝损伤
中性粒细胞在组织损伤部位,被环境信号激活,表现为在胞内合成颗粒蛋白和分泌囊泡,并转录趋化因子 [24]。在 DILI 的肝脏损伤部位,活化的中性粒细胞产生多种效应:1)脱颗粒,释放髓过氧化物酶(myeloperoxidase,MPO)、中性粒细胞弹性蛋白酶(neutrophil elastase,NE)等颗粒蛋白,对肝细胞产生毒性作用;2)分泌 TNF-α、IL-1β 等炎症因子,加剧肝脏炎症;3)诱导活性氧(reactive oxygen species,ROS)的生成,引起氧化应激,从而损伤肝脏;4)形成中性粒细胞胞外诱捕网,直接杀伤肝细胞,或介导肝血窦中微血栓的形成 [15]。
3
中性粒细胞在药物性肝损伤中的作用
3.1 中性粒细胞在直接型药物性肝损伤中的作用
阿司匹林、对乙酰氨基酚(acetaminophen,APAP)等非甾体抗炎药是造成直接型 DILI 的常见药物。APAP 诱导的急性肝损伤模型广泛应用于直接型 DILI 的机制研究。近年来,不仅在中国,在日本、韩国、新加坡等许多国家,因服用中草药而导致 DILI 的病例所占的比重也逐渐增加 [25]。一些中草药中的特定化学成分已被确定为肝毒性成分,对人体具有内在肝毒性,直接损伤肝脏,例如雷公藤中的雷公藤多苷和雷公藤甲素、蓖麻子中的蓖麻毒素、常山中的常山碱、合欢皮中的合欢皮皂苷、商陆中的商陆碱等 [26]。同时,多种肝毒性中草药(如菊三七、千里光等)中均含有不饱和吡咯里西啶生物碱,其诱导的 DILI 引起广泛关注 [27]。
3.1.1 中性粒细胞在对乙酰氨基酚导致的药物性肝损伤中的作用 APAP 是一种具有解热镇痛作用的非甾体抗炎药,服用过量时可导致急性肝损伤。APAP诱导的急性肝损伤已成为肝衰竭的主要原因之一,严重威胁人类健康。因此,APAP 导致 DILI 的毒理机制和治疗靶点研究引起广泛关注,其诱导的肝损伤动物模型也被大量研究。APAP 诱导的肝损伤发生早期,中性粒细胞迁移到小鼠肝组织中,大多数位于肝脏的健康部位;在肝损伤逐渐加重直至最高峰期间,大部分中性粒细胞迁移至坏死区域;然而在肝损伤恢复期,肝脏中中性粒细胞的数量则降低 [28]。
研究表明,中性粒细胞可加重 APAP 诱导的肝损伤,耗竭小鼠体内的中性粒细胞则可明显减轻APAP 诱导的肝损伤。在 APAP 导致的 DILI 中,中性粒细胞在肝脏中蓄积和活化,分泌 IL-1α 等炎症因子以加剧炎症,并释放髓过氧化物酶、一氧化氮和硝基酪氨酸,从而损伤肝细胞 [29]。研究表明,抑制中性粒细胞的肝脏浸润可减轻 APAP 导致的肝损伤,例如巨噬细胞迁移抑制因子抑制剂 [30]、溶解素 [21]、甘草次酸 [31]、胃泌素释放肽受体(gastrin-releasing peptide receptor,GRPR)拮抗剂均可通过抑制中性粒细胞的浸润,从而减轻肝损伤 [32]。激活白三烯 B4 受 体 1 型(leukotriene B4 receptor 1,BLT1)信号通路和肝线粒体脱氧核糖核酸(mitochondrial DNA,mtDNA)/Toll 样受体 9(Toll-like receptor 9,TLR9)/微小核糖核酸-223(microribonucleic acid-233,miRNA-233)通路也抑制中性粒细胞在肝脏中蓄积,从而保护肝脏免受APAP 肝毒性的影响 [33-34]。综上所述,在 APAP 诱导的 DILI 中,中性粒细胞向肝脏募集,释放炎症因子和颗粒蛋白,从而加重肝损伤。
3.1.2 中性粒细胞在雷公藤甲素导致的药物性肝损伤中的作用 雷公藤甲素是从雷公藤中提取出的主要活性成分,具有抗癌、抗类风湿、抗炎、抗阿尔茨海默病等药理作用 [35]。然而由于雷公藤甲素具有肝毒性、肾毒性和生殖毒性等多种毒性,可导致严重不良反应,其在临床上的应用受到限制。研究表明,在雷公藤甲素诱导的小鼠肝损伤模型中,随着肝损伤的发展,中性粒细胞在肝脏中蓄积和活化。肝脏中 CXCL1、TLR4、TLR9 和单核细胞趋化因子-1(monocyte chemoattractant protein-1,MCP-1)的表达增加,介导中性粒细胞向肝脏募集。肝脏中的中性粒细胞被激活后,释放髓过氧化物酶,分泌炎症因子,从而对肝细胞造成损伤。耗竭小鼠体内的中性粒细胞可使雷公藤甲素导致的肝损伤减轻 [36]。
3.1.3 中性粒细胞在吡咯里西啶生物碱导致的药物性肝损伤中的作用 吡咯里西啶生物碱诱导的肝损伤主要表现为肝窦内皮细胞损伤,肝组织淤血,形成微血栓阻塞肝窦,导致肝窦阻塞综合征 [25, 37]。野百合碱诱导的动物模型是目前研究吡咯里西啶生物碱导致的 DILI 的机制和治疗药物的常用模型 [38]。研究表明,在野百合碱诱导的 C57BL/6 小鼠模型中,肝脏中 HSP60 和 HMGB1 的释放增加,TLR4 的表达也增加,从而诱导中性粒细胞的募集,加剧炎症反应 [39]。肝窦内皮细胞与中性粒细胞之间存在相互作用,一方面肝窦内皮细胞参与中性粒细胞的趋化、黏附、迁移和活化过程,另一方面中性粒细胞也可损伤肝窦内皮细胞以加重肝损伤 [23, 40]。此外,中性粒细胞形成的中性粒细胞胞外诱捕网可促进凝血和抑制纤溶,从而导致血管内微血栓的形成 [41-42]。因此,靶向中性粒细胞可能成为研究吡咯里西啶生物碱导致的 DILI 的内在机制和治疗手段的新思路。
3.2 中性粒细胞在特异质型药物性肝损伤中的作用
特异质型 DILI 的不可预测性和不可重复性,是探究 DILI 的毒性机制和治疗靶点的难题。导致特异质型 DILI 的主要药物包括异烟肼、利福平、吡嗪酰胺等抗结核药和头孢菌素、氟喹诺酮类抗生素、大环内酯类抗生素等抗生素以及氟烷等 [11, 43-44]。氟烷是一种吸入性全身麻醉剂,可导致急性肝损伤,氟烷诱导的 DILI 模型是研究特异质型 DILI 的重要模型之一 [44]。不同品系的小鼠对氟烷肝毒性的易感程度不同,其中 BALB/c 小鼠最敏感,DBA/1 小鼠次之,C57BL/6J 小鼠无明显肝毒性。在氟烷诱导的 BALB/c 小鼠模型中,CXCL2 表达增加以招募中性粒细胞,促炎介质 IL-6、IL-1β、TNF-α、诱导型 一 氧 化 氮 合 酶(inducible nitric oxide synthase,iNOS)表达量增加,引发炎症反应 [45]。研究表明,在 BALB/c 的亚系 BALB/cJ 小鼠中,耗竭自然杀伤T 细胞以抑制中性粒细胞的募集,可降低对氟烷导致的 DILI 的易感性 [46]。在 C57BL/6J 小鼠中,敲除IL-10 以增加 CXCL2 的表达,从而诱导中性粒细胞的募集,则可增加对氟烷导致的 DILI 的易感性 [47]。值得关注的是,性别是氟烷引起 DILI 的风险因素之一,女性更易发生氟烷引起的 DILI。研究表明,在氟烷诱导的 BALB/c 小鼠模型中,雌二醇可使CXCL1、CXCL2 和 ICAM-1 在肝脏中高表达,诱导大量中性粒细胞浸润至肝脏,从而加重氟烷对肝脏的损伤,而黄体酮则可抑制中性粒细胞的募集,减轻氟烷对肝脏的损伤 [48]。女性体内雌二醇和黄体酮的平衡失调导致中性粒细胞向肝脏募集,可能是氟烷在女性中易感程度更高的重要原因。
氟喹诺酮类抗生素曲伐沙星导致的特异质型DILI 也与中性粒细胞相关 [49]。曲伐沙星因导致肝损伤发病率高而于 2001 年退市,距离其上市仅 3 年。研究显示,在中性粒细胞弹性蛋白酶缺陷小鼠中,曲伐沙星对肝脏的损伤作用减轻 [49]。这表明,中性粒细胞在损伤部位释放中性粒细胞弹性蛋白酶,参与曲伐沙星导致的肝损伤。
3.3 中性粒细胞在间接型药物性肝损伤中的作用
间接型 DILI 继发于药物的治疗作用,与药物的固有肝毒性或免疫原性无关,表现为诱发或加重肝脏疾病。其中,免疫检查点抑制剂导致的间接型 DILI 引起广泛关注。靶向的免疫检查点分子主要是程序性细胞死亡蛋白 1(programmed cell death protein 1,PD-1)、程序性死亡配体 1(programmed death-ligand 1,PD-L1)和细胞毒性 T 淋巴细胞相关蛋白 4(cytotoxic T-lymphocyte-associated protein 4,CTLA-4)[50]。随着免疫检查点抑制剂越来越多地用于癌症的治疗,免疫相关不良事件的发生逐渐增多。免疫检查点抑制剂导致的肝损伤主要表现为胆汁淤积、胆管炎和肝功能障碍 [50]。
免疫检查点抑制剂诱导的间接型 DILI,主要是由 T 细胞过度活化引起的细胞毒性导致 [51-52]。临床病例的肝活检结果显示,在抗 CTLA-4 治疗诱导的DILI 中,肝脏浸润的炎性细胞主要是 CD8+ T 细胞,而在抗 PD-1/PD-L1 治疗导致的 DILI 中,肝脏中CD4+ T 细胞和 CD8+ T 细胞的数量相当 [53]。研究表明,抗 PD-1 致 DILI 临床病例的肝脏中也存在中性粒细胞浸润 [54]。在用 B6/lpr 小鼠建立的抗 PD-1 和抗 CTLA-4 抗体共同诱导的肝损伤模型中也观察到CD4+ T细胞和CD8+ T细胞的肝脏浸润[55]。研究显示,中性粒细胞在流感病毒感染动物模型中介导 CD8+ T细胞的反应,在分支杆菌感染动物模型中促进CD4+ T细胞的活化和增殖 [56]。中性粒细胞可以分泌趋化因子以促进 T 细胞迁移到炎症部位,也可作为抗原呈递细胞,表达主要组织相容性复合体Ⅱ类、CD80和 CD86,从而驱动 CD4+ T 细胞和 CD8+ T 细胞活化 [56]。综上可见,在免疫检查点抑制剂导致的肝损伤中,中性粒细胞可能促进 CD4+ 和 CD8+ T 细胞活化,从而加重肝损伤。
4
结语与展望
综上所述,中性粒细胞在骨髓中分化和成熟,受到 DAMPs、趋化因子和炎症因子等环境信号的刺激,从骨髓释放至血液循环,通过肝血窦进入肝脏,抵达损伤部位并活化。活化后的中性粒细胞脱颗粒、促进炎症、生成 ROS 和形成中性粒细胞胞外诱捕网,从而损伤肝细胞。在多种 DILI 中,中性粒细胞均具有加重肝损伤作用。
目前在 APAP 诱导的急性肝损伤中,中性粒细胞对肝损伤的作用仍存在争议,可能与实验观察的时间点差异相关。研究表明,在 APAP 诱导的急性肝损伤修复期,中性粒细胞可通过生成 ROS,介导促炎性巨噬细胞向修复性巨噬细胞转化,从而促进肝脏修复 [57-58]。在 APAP 诱导的 DILI 病理生理过程的不同阶段,中性粒细胞发挥的效能可能不同,以单一时间点作为实验观察点得到的实验结果可能存在片面性。为更深入地探究中性粒细胞在 APAP 诱导的 DILI 的不同阶段中的作用,还需在肝损伤加重期和修复期内设置多个实验观察时间点,考察肝脏损伤和修复情况 [59]。
中性粒细胞弹性蛋白酶是一种由活化的中性粒细胞分泌的颗粒蛋白,其抑制剂西维来司和 N-乙酰半胱氨酸联合治疗可显著减轻肝损伤,比 N-乙酰半胱氨酸单药治疗更有效,为 APAP 诱导的 DILI 的治疗提供新的选择。另外,研究表明,一些小分子物质也可通过抑制中性粒细胞的招募和浸润以减轻APAP 诱导的 DILI,如小檗碱 [60]、栀子苷 [61]、柠檬醛 [62] 等。中性粒细胞可能是预防和治疗 APAP 诱导的急性肝衰竭的潜在靶点。抑制中性粒细胞向肝脏募集,降低其对肝脏的损伤作用可作为 DILI 的潜在治疗方案,也为避免新药肝毒性提供思路。虽然中性粒细胞在 DILI 中的作用机制研究已取得一定进展,但其在 DILI 治疗中的作用仍须进一步探索,与其他损伤相关成分之间的相互作用尚须进一步研究。
参考文献 :
[1] 肇丽梅 , 缪丽燕 . 药物性肝损伤研究进展及展望 [J]. 药学进展 , 2023, 47(2): 81-83.
[2] Brennan P N, Cartlidge P, Manship T, et al. Guideline review: EASL clinical practice guidelines: drug-induced liver injury (DILI)[J].Frontline Gastroenterol, 2021, 13(4): 332-336.
[3] Bessone F, García-Cortés M, Medina-Caliz I, et al. Herbal and dietary supplements-induced liver injury in Latin America: experience from the LATINDILI network[J]. Clin Gastroenterol Hepatol, 2022, 20(3): e548-e563. DOI: 10.1016/j.cgh.2021.01.011.
[4] Björnsson E S, Bergmann O M, Björnsson H K, et al. Incidence, presentation, and outcomes in patients with drug-induced liver injury in the general population of Iceland[J]. Gastroenterology, 2013, 144(7): 1419-1425.
[5] Li X, Tang J, Mao Y. Incidence and risk factors of drug-induced liver injury[J]. Liver Int, 2022, 42(9): 1999-2014.
[6] Devarbhavi H, Aithal G, Treeprasertsuk S, et al. Drug-induced liver injury: Asia pacific association of study of liver consensus guidelines[J].Hepatol Int, 2021, 15(2): 258-282.
[7] Philips C A, Augustine P, Rajesh S, et al. Complementary and alternative medicine-related drug-induced liver injury in Asia[J]. J Clin Transl Hepatol, 2019, 7(3): 263-274.
[8] Wai C T, Tan B H, Chan C L, et al. Drug-induced liver injury at an Asian center: a prospective study[J]. Liver Int, 2007, 27(4): 465-474.
[9] Ma Z T, Shi Z, Xiao X H, et al. New insights into herb-induced liver injury[J]. Antioxid Redox Signal, 2023, 38(16/17/18): 1138-1149.
[10] Björnsson H K, Björnsson E S. Drug-induced liver injury: pathogenesis, epidemiology, clinical features, and practical management[J]. Eur J Intern Med, 2022, 97: 26-31. DOI: 10.1016/j.ejim.2021.10.035.
[11] Hoofnagle J H, Björnsson E S. Drug-induced liver injury-types and phenotypes[J]. N Engl J Med, 2019, 381(3): 264-273.
[12] Ke L, Lu C, Shen R, et al. Knowledge mapping of drug-induced liver injury: a scientometric investigation (2010-2019)[J]. Front Pharmacol, 2020, 11: 842. DOI: 10.3389/fphar.2020.00842.
[13] Miller E D, Abu-Sbeih H, Styskel B, et al. Clinical characteristics and adverse impact of hepatotoxicity due to immune checkpoint inhibitors[J]. Am J Gastroenterol, 2020, 115(2): 251-261.
[14] Burn G L, Foti A, Marsman G, et al. The neutrophil[J]. Immunity, 2021, 54(7): 1377-1391.
[15] Rawat K, Shrivastava A. Neutrophils as emerging protagonists and targets in chronic inflammatory diseases[J]. Inflamm Res, 2022, 71(12): 1477-1488.
[16] Wigerblad G, Kaplan M J. NETs spread ever wider in rheumatic diseases[J]. Nat Rev Rheumatol, 2020, 16(2): 73-74.
[17] Silvestre-Roig C, Braster Q, Ortega-Gomez A, et al. Neutrophils as regulators of cardiovascular inflammation[J]. Nat Rev Cardiol, 2020, 17(6): 327-340.
[18] Lawrence S M, Corriden R, Nizet V. The ontogeny of a neutrophil: mechanisms of granulopoiesis and homeostasis[J]. Microbiol Mol Biol Rev, 2018, 82(1): e00057-17. DOI: 10.1128/MMBR.00057-17.
[19] Yvan-Charvet L, Ng L G. Granulopoiesis and neutrophil homeostasis: a metabolic, daily balancing act[J]. Trends Immunol, 2019, 40(7): 598-612.
[20] Teng T S, Ji A L, Ji X Y, et al. Neutrophils and immunity: from bactericidal action to being conquered[J]. J Immunol Res, 2017: 9671604. DOI: 10.1155/2017/9671604.
[21] Patel S J, Luther J, Bohr S, et al. A novel resolvin-based strategy for limiting acetaminophen hepatotoxicity[J]. Clin Transl Gastroenterol, 2016, 7(3): e153. DOI: 10.1038/ctg.2016.13.
[22] Guo H, Chen S, Xie M, et al. The complex roles of neutrophils in APAP-induced liver injury[J]. Cell Prolif, 2021, 54(6): e13040. DOI: 10.1111/cpr.13040.
[23] Wang Y, Liu Y. Neutrophil-induced liver injury and interactions between neutrophils and liver sinusoidal endothelial cells[J].Inflammation, 2021, 44(4): 1246-1262.
[24] Amulic B, Cazalet C, Hayes G L, et al. Neutrophil function: from mechanisms to disease[J]. Annu Rev Immunol, 2012, 30: 459-489. DOI: 10.1146/annurev-immunol-020711-074942.
[25] Pan X, Zhou J, Chen Y, et al. Classification, hepatotoxic mechanisms, and targets of the risk ingredients in traditional Chinese medicineinduced liver injury[J]. Toxicol Lett, 2020, 323: 48-56. DOI: 10.1016/j.toxlet.2020.01.026.
[26] Jing J, Teschke R. Traditional chinese medicine and herb-induced liver injury: comparison with drug-induced liver injury[J]. J Clin Transl Hepatol, 2018, 6(1): 57-68.
[27] Wei X, Ruan W, Vrieling K. Current knowledge and perspectives of pyrrolizidine alkaloids in pharmacological applications: a minireview[J]. Molecules, 2021, 26(7): 1970. DOI: 10.3390/molecules 26071970.
[28] Williams C D, Bajt M L, Sharpe M R, et al. Neutrophil activation during acetaminophen hepatotoxicity and repair in mice and humans[J].Toxicol Appl Pharmacol, 2014, 275(2): 122-133.
[29] Gong L, Liao L, Dai X, et al. The dual role of immune response in acetaminophen hepatotoxicity: implication for immune pharmacological targets[J]. Toxicol Lett, 2021, 351: 37-52. DOI: 10.1016/j.toxlet.2021.08.009.
[30] Ohkawara T, Okubo N, Maehara O, et al. Protective effect of ISO-1 with inhibition of RIPK3 up-regulation and neutrophilic accumulation on acetaminophen-induced liver injury in mice[J]. Toxicol Lett, 2021, 39: 51-59. DOI: 10.1016/j.toxlet.2020.12.015.
[31] Yang G, Zhang L, Ma L, et al. Glycyrrhetinic acid prevents acetaminophen-induced acute liver injury via the inhibition of CYP2E1 expression and HMGB1-TLR4 signal activation in mice[J].Int Immunopharmacol, 2017, 50: 186-193. DOI: 10.1016/j.intimp. 2017.06.027.
[32] Czepielewski R S, Jaeger N, Marques P E, et al. GRPR antagonist protects from drug-induced liver injury by impairing neutrophil chemotaxis and motility[J]. Eur J Immunol, 2017, 47(4): 646-657.
[33] He Y, Feng D, Li M, et al. Hepatic mitochondrial DNA/Toll-like receptor 9/microRNA-223 forms a negative feedback loop to limit neutrophil overactivation and acetaminophen hepatotoxicity in mice[J].Hepatology, 2017, 66(1): 220-234.
[34] Kojo K, Ito Y, Eshima K, et al. BLT1 signalling protects the liver against acetaminophen hepatotoxicity by preventing excessive accumulation of hepatic neutrophils[J]. Sci Rep, 2016, 6: 29650. DOI: 10.1038/srep29650.
[35] Tong L, Zhao Q, Datan E, et al. Triptolide: reflections on two decades of research and prospects for the future[J]. Nat Prod Rep, 2021, 38(4): 843-860.
[36] Wang X Z, Zhang S Y, Xu Y, et al. The role of neutrophils in triptolideinduced liver injury[J]. Chin J Nat Med, 2018, 16(9): 653-664.
[37] Zhu L, Zhang C Y, Li D P, et al. Tu-San-Qi (Gynura japonica): the culprit behind pyrrolizidine alkaloid-induced liver injury in China[J].Acta Pharmacol Sin, 2021, 42(8): 1212-1222.
[38] Kumar A, Palek R, Liska V. A critical analysis of experimental animal models of sinusoidal obstruction syndrome[J]. J Clin Exp Hepatol, 2019, 9(3): 345-353.
[39] Huang Z, Chen M, Wei M, et al. Liver inflammatory injury initiated by DAMPs-TLR4-MyD88/TRIF-NFκB signaling pathway is involved in monocrotaline-induced HSOS[J]. Toxicol Sci, 2019, 172(2): 385-397.
[40] Hilscher M B, Sehrawat T, Arab J P, et al. Mechanical stretch increases expression of CXCL1 in liver sinusoidal endothelial cells to recruit neutrophils, generate sinusoidal microthombi, and promote portal hypertension[J]. Gastroenterology, 2019, 157(1): 193-209.
[41] Zhou J, Guo P, Hao X, et al. Neutrophil extracellular traps (NETs): a new therapeutic target for neuroinflammation and microthrombosis after subarachnoid hemorrhage?[J]. Transl Stroke Res, 2023, 14(4): 443-445.
[42] Zuo Y, Kanthi Y, Knight J S, et al. The interplay between neutrophils, complement, and microthrombi in COVID-19[J]. Best Pract Res Clin Rheumatol, 2021, 35(1): 101661. DOI: 10.1016/j.berh.2021.101661.
[43] Kumar P S, Vidya R, Tabassum, et al. Anti-tuberculosis treatment: induced hepatotoxicity-a case report[J]. EJIFCC, 2020, 31(3): 242-247.
[44] Jia R, Oda S, Yokoi T. Pharmacological evidence for the involvement of ryanodine receptors in halothane-induced liver injury in mice[J].Toxicology, 2020, 443: 152560. DOI: 10.1016/j.tox.2020.152560.
[45] You Q, Cheng L, Reilly T P, et al. Role of neutrophils in a mouse model of halothane-induced liver injury[J]. Hepatology, 2006, 44(6): 1421-1431.
[46] Cheng L, You Q, Yin H, et al. Involvement of natural killer T cells in halothane-induced liver injury in mice[J]. Biochem Pharmacol, 2010, 80(2): 255-261.
[47] Feng D, Wang Y, Xu Y, et al. Interleukin 10 deficiency exacerbates halothane induced liver injury by increasing interleukin 8 expression and neutrophil infiltration[J]. Biochem Pharmacol, 2009, 77(2): 277-284.
[48] Toyoda Y, Miyashita T, Endo S, et al. Estradiol and progesterone modulate halothane-induced liver injury in mice[J]. Toxicol Lett, 2011, 204(1): 17-24.
[49] Shaw P J, Ganey P E, Roth R A. Trovafloxacin enhances the inflammatory response to a Gram-negative or a Gram-positive bacterial stimulus, resulting in neutrophil-dependent liver injury in mice[J]. J Pharmacol Exp Ther, 2009, 330(1): 72-78.
[50] Remash D, Prince D S, McKenzie C, et al. Immune checkpoint inhibitor-related hepatotoxicity: a review[J]. World J Gastroenterol, 2021, 27(32): 5376-5391.
[51] Da Cunha T, Wu G Y, Vaziri H. Immunotherapy-induced hepatotoxicity: a review[J]. J Clin Transl Hepatol, 2022, 10(6): 1194-1204.
[52] Shojaie L, Ali M, Iorga A, et al. Mechanisms of immune checkpoint inhibitor-mediated liver injury[J]. Acta Pharm Sin B, 2021, 11(12): 3727-3739.
[53] de Martin E, Michot J M, Papouin B, et al. Characterization of liver injury induced by cancer immunotherapy using immune checkpoint inhibitors[J]. J Hepatol, 2018, 68(6): 1181-1190.
[54] Zen Y, Yeh M M. Hepatotoxicity of immune checkpoint inhibitors: a histology study of seven cases in comparison with autoimmune hepatitis and idiosyncratic drug-induced liver injury[J]. Mod Pathol, 2018, 31(6): 965-973.
[55] Adam K, Iuga A, Tocheva A S, et al. A novel mouse model for checkpoint inhibitor-induced adverse events[J]. PLoS One, 2021, 16(2): e0246168. DOI: 10.1371/journal.pone.0246168.
[56] Kalyan S, Kabelitz D. When neutrophils meet T cells: beginnings of a tumultuous relationship with underappreciated potential[J]. Eur J Immunol, 2014, 44(3): 627-633.
[57] Mattos M S, Lopes M E, de Araujo A M, et al. Prolonged neutrophil survival at necrotic sites is a fundamental feature for tissue recovery and resolution of hepatic inflammation[J]. J Leukoc Biol, 2020, 108(4): 1199-1213.
[58] Yang W, Tao Y, Wu Y, et al. Neutrophils promote the development of reparative macrophages mediated by ROS to orchestrate liver repair[J]. Nat Commun, 2019, 10(1): 1076. DOI: 10.1038/s41467-019-09046-8.
[59] Jaeschke H, Ramachandran A. Mechanisms and pathophysiological significance of sterile inflammation during acetaminophen hepatotoxicity[J]. Food Chem Toxicol, 2020, 138: 111240. DOI: 10. 1016/j.fct.2020.111240.
[60] Zhao Z, Wei Q, Hua W, et al. Hepatoprotective effects of berberine on acetaminophen-induced hepatotoxicity in mice[J]. Biomed Pharmacother, 2018, 103: 1319-1326. DOI: 10.1016/j.biopha. 2018.04.175.
[61] Yang S, Kuang G, Jiang R, et al. Geniposide protected hepatocytes from acetaminophen hepatotoxicity by down-regulating CYP 2E1 expression and inhibiting TLR 4/NF-κB signaling pathway[J]. Int Immunopharmacol, 2019, 74: 105625. DOI: 10.1016/j.intimp.2019. 05.010.
[62] Uchida N S, Silva-Filho S E, Cardia G F E, et al. Hepatoprotective effect of citral on acetaminophen-induced liver toxicity in mice[J].Evid Based Complement Alternat Med, 2017: 1796209. DOI: 10.1155/ 2017/1796209.
美编排版:陈鑫茹
感谢您阅读《药学进展》微信平台原创好文,也欢迎各位读者转载、引用。本文选自《药学进展》2024年第 10 期。
《药学进展》杂志由中国药科大学和中国药学会共同主办、国家教育部主管,中国科技核心期刊(中国科技论文统计源期刊)。刊物以反映药学科研领域的新方法、新成果、新进展、新趋势为宗旨,以综述、评述、行业发展报告为特色,以药学学科进展、技术进展、新药研发各环节技术信息为重点,是一本专注于医药科技前沿与产业动态的专业媒体。
《药学进展》注重内容策划、加强组稿约稿、深度挖掘、分析药学信息资源、在药学学科进展、科研思路方法、靶点机制探讨、新药研发报告、临床用药分析、国际医药前沿等方面初具特色;特别是医药信息内容以科学前沿与国家战略需求相合,更加突出前瞻性、权威性、时效性、新颖性、系统性、实战性。根据最新统计数据,刊物篇均下载率连续三年蝉联我国医药期刊榜首,复合影响因子1.216,具有较高的影响力。
《药学进展》编委会由国家重大专项化学药总师陈凯先院士担任主编,编委由新药研发技术链政府监管部门、高校科研院所、制药企业、临床医院、CRO、金融资本及知识产权相关机构近两百位极具影响力的专家组成。
联系《药学进展》↓↓↓
编辑部官网:pps.cpu.edu.cn;
邮箱:yxjz@163.com;
电话:025-83271227。
欢迎投稿、订阅!
往期推荐
聚焦“兴药为民·2023生物医药创新融合发展大会”“兴药为民·2023生物医药创新融合发展大会”盛大启幕!院士专家齐聚杭城,绘就生物医药前沿赛道新蓝图“兴药强刊”青年学者论坛暨《药学进展》第二届青年编委会议成功召开“兴药为民·2023生物医药创新融合发展大会”路演专场圆满收官!校企合作新旅程已启航
我知道你在看哟
临床研究
100 项与 甲磺酸阿曲沙星 相关的药物交易
登录后查看更多信息
研发状态
10 条最早获批的记录, 后查看更多信息
登录
| 适应症 | 国家/地区 | 公司 | 日期 |
|---|---|---|---|
| 细菌感染 | 美国 | 1997-12-18 |
登录后查看更多信息
临床结果
临床结果
适应症
分期
评价
查看全部结果
| 研究 | 分期 | 人群特征 | 评价人数 | 分组 | 结果 | 评价 | 发布日期 |
|---|
No Data | |||||||
登录后查看更多信息
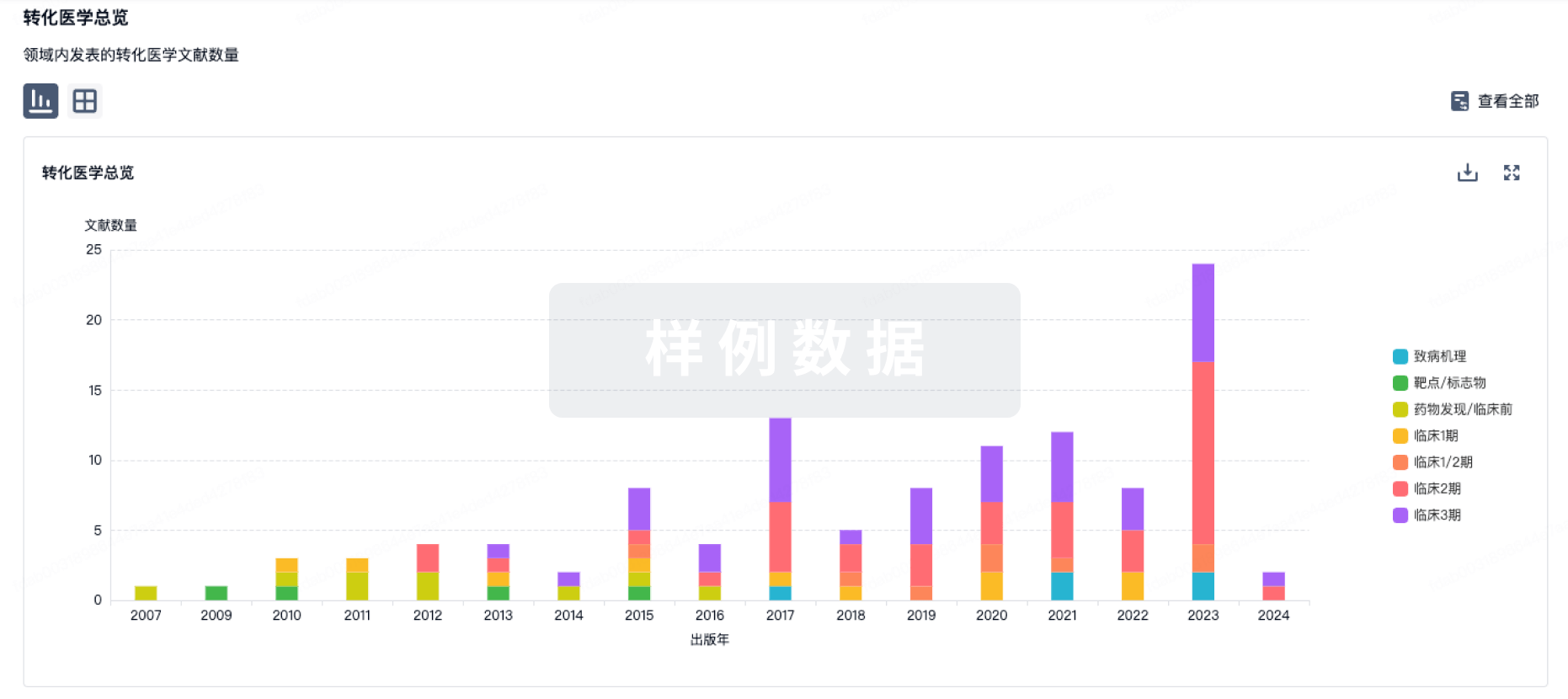
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
核心专利
使用我们的核心专利数据促进您的研究。
登录
或
临床分析
紧跟全球注册中心的最新临床试验。
登录
或
批准
利用最新的监管批准信息加速您的研究。
登录
或
特殊审评
只需点击几下即可了解关键药物信息。
登录
或
生物医药百科问答
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用