预约演示
更新于:2026-05-16
Anti PD-1 antibody(Chulalongkorn University)
抗PD-1抗体(Chulalongkorn University)
更新于:2026-05-16
概要
基本信息
非在研机构- |
权益机构- |
最高研发阶段临床前 |
首次获批日期- |
最高研发阶段(中国)- |
特殊审评- |
关联
100 项与 抗PD-1抗体(Chulalongkorn University) 相关的临床结果
登录后查看更多信息
100 项与 抗PD-1抗体(Chulalongkorn University) 相关的转化医学
登录后查看更多信息
100 项与 抗PD-1抗体(Chulalongkorn University) 相关的专利(医药)
登录后查看更多信息
76
项与 抗PD-1抗体(Chulalongkorn University) 相关的文献(医药)2026-04-01·CANCER SCIENCE
Impact of Sequential Ramucirumab Plus Docetaxel After
PD
‐1 Inhibitors on Anti‐
PD
‐1 Antibody‐Bound T‐Cell Dynamics and Clinical Outcomes
Article
作者: Koyama, Shohei ; Kuge, Tomoki ; Minami, Toshiyuki ; Kumanogoh, Atsushi ; Takakura, Nobuyuki ; Naito, Yujiro ; Satoh, Shingo ; Tamiya, Akihiro ; Mori, Masahide ; Takeda, Yoshito ; Nishino, Kazumi ; Okishio, Kyoichi ; Matsumoto, Kinnosuke ; Kinehara, Yuhei ; Shiroyama, Takayuki ; Tetsumoto, Satoshi ; Tamiya, Motohiro ; Niki, Toshie ; Suzuki, Hidekazu ; Suga, Yasuhiko ; Osa, Akio
ABSTRACT:
Ramucirumab plus docetaxel (RAM+DOC) demonstrates clinical activity after programmed cell death‐1 (PD‐1) inhibitors in advanced non‐small cell lung cancer (NSCLC); however, the underlying mechanisms remain unclear. We aimed to evaluate clinical efficacy and explore immunologic dynamics and benefit‐associated biomarkers. Patients treated with RAM+DOC after PD‐1 inhibitors were enrolled in a multicenter prospective cohort. Anti‐PD‐1 antibody bound (IgG4
+
) T‐cell subsets were measured at baseline (T0) and after 2–3 cycles (T1), reflecting residual anti‐PD‐1 antibody binding on circulating T cells. T1/T0 ratios of immune subsets were calculated to assess dynamics. Landmark analyses at T1 evaluated associations with progression‐free survival (PFS) and overall survival (OS). Prognostic biomarkers were assessed at baseline. Among 27 evaluable patients, the objective response rate was 37.0%, median PFS 5.1 months, and OS 10.4 months. RAM + DOC responders had higher IgG4
+
CD8
+
T‐cell and lower IgG4
+
Treg T1/T0 ratios (both
p
< 0.001). Higher IgG4
+
CD8
+
ratios were associated with longer landmark PFS (
p
= 0.002) and OS (
p
= 0.016) and were inversely correlated with IgG4
+
Treg ratios (
p
= 0.008). Among the baseline factors, high IgG4
+
CD8
+
Temra conferred survival benefits (OS, not reached vs. 8.2 months;
p
= 0.006), and low vascular endothelial growth factor (VEGF)‐C levels were associated with longer OS (not reached vs. 8.5 months,
p
= 0.044). Both variables remained independent prognostic factors of PFS and OS in multivariable analysis. Our findings suggest that sequential strategy administering RAM+DOC during persistent binding of anti–PD‐1 antibody to T cells may be beneficial. IgG4
+
CD8
+
Temra and VEGF‐C levels at RAM+DOC initiation may serve as biomarkers of survival benefit.
Trial Registration:
UMIN‐Clinical Trials Registry (UMIN000050478)
2026-04-01·MedComm
Sulforaphane Synergizes With PD‐1 Blockade Through Activating CD8
+
T Cells in Non–Small Cell Lung Cancer: Preclinical and Clinical Investigations
Article
作者: Liu, Shasha ; Yue, Dongli ; Zhang, Tengyue ; Liu, Yanfen ; Zhang, Yi ; Zhao, Ming ; Fu, Ziyi ; Wang, Liping ; Huang, Jianmin ; Gao, Qun ; Liu, Jinyan ; Guo, Caijuan ; Wang, Zheng ; Li, Yixin ; Li, Jieyao ; You, Mingming ; Yang, Shuangning
ABSTRACT:
Anti‐PD‐1/PD‐L1 therapy has achieved promising success across several tumor types; however, its efficacy is still far from satisfactory in non–small cell lung cancer (NSCLC). Combining therapies have been attempted to synergize anti‐PD‐1/PD‐L1 therapy through activating antitumor response. Previously, we convinced the role of sulforaphane (SFN) in regulating tumor immune microenvironment (TME) to enhance antitumor response. Consistently, here we observed combining SFN with chemotherapy and anti‐PD‐1 therapy achieved the best tumor suppression versus other treatments in mouse models bearing Lewis lung carcinoma cells.
Further, a clinical trial (KY‐2021‐0266) was performed, and the disease control and objective response rates were higher in the experimental group (SFN combined anti‐PD‐1 antibody and chemotherapy group,
n
= 30) compared with the control group (anti‐PD‐1 antibody combined chemotherapy group,
n
= 30) (100% vs. 93.3% and 86.7% vs. 60.0%, respectively). Moreover, the median progression‐free survival was longer (19 vs. 9.5 months, respectively) in the experimental group. After treatment, antitumor response was enriched, while CD8‐related function markers were elevated and myeloid‐derived suppressor cell/M2‐related markers were reduced in the experimental group. Two spurious progressions were observed in the experimental group. In conclusion, this synergistic effect suggests that SFN may be a promising immunosensitizer and a treatment option in NSCLC.
2026-01-01·EXPERIMENTAL DERMATOLOGY
Phase
III
Study Investigating the Safety and Efficacy of
TM5614
in Combination With Nivolumab for the Treatment of Unresectable Malignant Melanoma: Protocol
Article
作者: Matsushita, Shigeto ; Hashimoto, Akira ; Tanemura, Atsushi ; Ohe, Shuichi ; Iwata, Hiroaki ; Takenouchi, Tatsuya ; Kato, Hiroshi ; Yamazaki, Emi ; Namikawa, Kenjiro ; Yoshikawa, Shusuke ; Kambayashi, Yumi ; Nakamura, Yoshiyuki ; Ohno, Fumitaka ; Maekawa, Takeo ; Fujisawa, Yasuhiro ; Yamasaki, Osamu ; Takahashi‐Watanabe, Manami ; Yoshino, Koji ; Fujimura, Taku ; Uhara, Hisashi ; Rokunohe, Daiki ; Asano, Yoshihide ; Takahashi, Akira ; Uchi, Hiroshi ; Fukushima, Satoshi ; Muto, Yusuke ; Ito, Takamichi
ABSTRACT:
TM5614, a plasminogen activator inhibitor‐1 (PAI‐1) inhibitor, has demonstrated potential in overcoming nivolumab resistance in patients with unresectable, anti‐PD‐1 antibody‐refractory malignant melanoma. Previous Phase II trials have shown promising response rates, warranting further investigation through a randomised, placebo‐controlled Phase III trial. This Phase III, randomised, double‐blind, placebo‐controlled, investigator‐initiated clinical trial aims to evaluate the efficacy and safety of TM5614 in combination with nivolumab for treating patients with unresectable, anti‐PD‐1 antibody‐refractory malignant melanoma. The study will enrol 124 participants across 18 medical institutions in Japan. Patients will receive either TM5614 plus nivolumab or placebo plus nivolumab for 48 weeks. The primary endpoint is overall survival (OS), while secondary endpoints include objective response rate (ORR), progression‐free survival (PFS), disease control rate (DCR), and the incidence of treatment‐related adverse events. This study is designed to compare OS between the TM5614 plus nivolumab group and the placebo plus nivolumab group. Statistical analyses will be conducted using the Kaplan–Meier method and Cox proportional hazards models. This Phase III trial will provide critical insights into the efficacy of TM5614 in combination with nivolumab as a novel treatment strategy for unresectable, anti‐PD‐1 antibody‐refractory malignant melanoma. The results may offer new therapeutic options to improve patient outcomes and overcome resistance to immune checkpoint inhibitors.
Trial Registration:
jRCT2021240049 (
https://jrct.niph.go.jp/en‐latest‐detail/jRCT2021240049
). Protocol version: Ver.1.2 (3rd February, 2025)
56
项与 抗PD-1抗体(Chulalongkorn University) 相关的新闻(医药)2026-05-12
·生物探索
引言
为什么有些癌症对免疫疗法似乎具有天生的抵抗力?长期以来,我们习惯于在肿瘤细胞的突变负荷或是免疫细胞的衰竭状态中寻找答案。
然而,5月7日,《Cell》的研究报道“A blood-brain barrier-like vascular gate limits immunotherapy efficacy in neuroendocrine cancers”,这项研究跳出了传统的细胞博弈视角,发现限制免疫系统发挥作用的关键,可能在于肿瘤内部巧妙构建的一套“交通管制系统”。通过深入解析小细胞肺癌(Small cell lung cancer, SCLC)及其微环境,研究人员发现了一种特殊的血管结构。这扇被肿瘤精心打造的“门”,将试图攻击它的免疫细胞死死地挡在了外围。
免疫荒漠中的悖论与反常的血管
在探讨肿瘤免疫治疗时,我们常常会提及一个重要指标:肿瘤突变负荷(Tumor mutation burden, TMB)。通常情况下,TMB越高的肿瘤,其表面呈现的异常抗原就越多,越容易被免疫系统识别并攻击,因此这类患者往往能从免疫检查点抑制剂(Immune checkpoint inhibitors, ICIs)的治疗中获益。然而,小细胞肺癌却是一个巨大的反常悖论。它拥有极高的突变负荷,是侵袭性最强、恶性程度最高的神经内分泌肿瘤(Neuroendocrine cancers, NECs)之一,但它却是一个典型的“免疫荒漠”(Immune-desert)肿瘤,其微环境中的细胞毒性T细胞浸润极少。在IMpower133和CASPIAN等大型临床试验中,即便使用了先进的免疫疗法,患者的总生存期(Overall survival, OS)获益也仅仅徘徊在可怜的2个月左右。
究竟是什么阻碍了免疫细胞进入这片“战场”?研究人员将目光转向了泛癌种的转录组学数据。在对413例接受免疫治疗的泛癌种患者队列进行深度分析时,一个不寻常的信号浮出水面:多种与血管相关的通路,包括内皮细胞形态发生和血管平滑肌细胞发育,与T细胞的浸润程度呈现出显著的负相关。这暗示着,血管本身可能就是阻止免疫细胞进入的元凶。
进一步的单细胞转录组测序对比了小细胞肺癌与非小细胞肺癌(Non-small cell lung cancer, NSCLC)的微环境。结果令人大跌眼镜:在小细胞肺癌所有的微环境细胞群体中,内皮细胞(Endothelial cells)的差异表达基因(Differentially expressed genes, DEGs)数量竟然是最高的,接近800个之多。这些差异基因大量富集在细胞外基质(Extracellular matrix, ECM)和基底膜(Basement membrane)相关的通路上。显然,小细胞肺癌的血管内皮细胞正在秘密执行着某种非同寻常的生物学程序。堪比血脑屏障:探秘血管门的建筑构造
在传统的肿瘤生物学认知中,为了满足快速生长的营养需求,肿瘤往往会诱导生成大量的新生血管。但这些血管通常是杂乱无章、高度渗漏的,基底膜薄弱且缺乏周细胞的包裹。然而,当研究人员将小细胞肺癌的血管组织置于显微镜下时,却看到了截然不同的一幕。
病理学分析显示,小细胞肺癌的血管内皮细胞排列极其规则,更令人惊讶的是,它们被一层异常厚实的基底膜紧紧包裹。数据对比直观地揭示了这种差异:非小细胞肺癌的血管基底膜厚度通常不足2微米,而小细胞肺癌的血管基底膜厚度平均超过了6微米,甚至更厚。透射电子显微镜下的观察进一步证实,这些内皮细胞之间存在着极其频繁且紧密的紧密连接(Tight junctions),由关键蛋白ZO-1标记;同时,血管外围布满了由α平滑肌肌动蛋白(αSMA)和血小板衍生生长因子受体β(PDGFRβ)标记的致密周细胞(Pericyte)。
这种由厚实的基底膜、密集的紧密连接和高度覆盖的周细胞构成的三重防线,在结构上惊人地模仿了人体内最严密的生理屏障——血脑屏障(Blood-brain barrier, BBB)。研究人员将其命名为血脑屏障样血管门(Blood-brain barrier-like vascular gate, BVG)。
这道“血管门”在功能上是否真如其结构般坚不可摧?研究人员通过尾静脉注射伊文思蓝染料(Evans blue)进行了血管通透性测试。在非小细胞肺癌的原位肿瘤中,染料大量渗漏到血管外的肿瘤组织中;但在小细胞肺癌中,染料被死死限制在血管腔内,几乎没有任何渗出。更为关键的是,当研究人员向携带这两种肿瘤的小鼠体内输入特异性识别肿瘤抗原的OT-1 T细胞后,他们发现非小细胞肺癌中每个视野下能有超过40个T细胞成功浸润,而在小细胞肺癌中,成功越过血管壁的T细胞数量寥寥无几,通常不到10个。这道物理屏障,确实将免疫大军拒之门外。幕后黑手现身:肿瘤细胞的基因操控术
既然小细胞肺癌和非小细胞肺癌可以生长在完全相同的肺部器官微环境中,为什么只有小细胞肺癌能长出这种奇特的血管门?为了回答这个问题,研究人员将这两种肿瘤细胞分别注射到小鼠的皮下组织进行培养。结果显示,哪怕离开了肺部环境,皮下生长的小细胞肺癌依然能够构建出带有厚实基底膜和周细胞覆盖的BVG结构。这充分说明,建造这堵高墙的图纸并非来自周围的环境,而是由小细胞肺癌细胞自身主导和设计的。
那么,是谁在肿瘤细胞内部发号施令?通过将小细胞肺癌特异性的基因表达特征与BVG基因标签进行相关性排序,研究人员锁定了一个关键的转录因子:ASCL1(Achaete-scute family bHLH transcription factor 1)。ASCL1不仅是经典神经内分泌亚型小细胞肺癌的诊断标志物,更是决定细胞命运的主调控因子(Master regulator)。
当研究人员利用基因编辑技术敲除肿瘤细胞中的Ascl1基因后,原本坚固的BVG结构轰然倒塌。血管基底膜的厚度显著下降,周细胞的覆盖和紧密连接也随之消散。顺藤摸瓜,研究团队发现ASCL1直接转录调控了其下游的胰岛素样生长因子结合蛋白5(Insulin-like growth factor-binding protein 5, IGFBP5)。IGFBP5在小细胞肺癌中高度表达,而在非小细胞肺癌中几乎难觅踪影。
敲除Igfbp5基因产生了与敲除Ascl1极其相似的破坏力:BVG结构瓦解,血管通透性恢复,原本被挡在门外的T细胞开始大量涌入肿瘤内部。机制上,肿瘤细胞分泌的IGFBP5能够结合IGF1,并特异性地激活血管内皮细胞表面胰岛素样生长因子1受体(Insulin-like growth factor 1 receptor, IGF1R)介导的信号通路。为了证实这一单向的信号传递过程,研究人员巧妙地将缺乏Igf1r的内皮细胞与小细胞肺癌细胞混合移植。不出所料,面对缺乏接收天线的内皮细胞,肿瘤细胞再也无法诱导血管门的形成,肿瘤组织内的T细胞数量从单视野下的个位数猛增至25个以上。
至此,一条清晰的因果链条浮出水面:肿瘤细胞通过ASCL1-IGFBP5-IGF1R轴,远程操控了内皮细胞,硬生生在自己周围筑起了一道“血脑屏障”。推翻高墙:重塑血管以唤醒免疫应答
发现问题是为了解决问题。既然ASCL1-IGFBP5-IGF1R轴是维持这道免疫排斥屏障的生命线,那么阻断这条通路能否逆转小细胞肺癌对免疫治疗的抵抗?
在动物模型中,仅仅敲除肿瘤细胞的Igfbp5基因在体外并不会影响其生长,但在小鼠体内由于微环境的改变,肿瘤生长受到了一定程度的抑制。然而,真正令人振奋的转变发生在联合免疫治疗之时。面对完整的BVG屏障,单用抗PD-1抗体治疗对小细胞肺癌几乎无效;但当面对Igfbp5缺失、失去BVG保护的肿瘤时,抗PD-1抗体展现出了摧枯拉朽的威力,几乎完全抑制了肿瘤的生长。
为了将这一发现推向临床应用,研究人员测试了一种已在早期临床试验中验证过安全性的IGF1R抑制剂——OSI-906(Linsitinib)。正如预测的那样,单独使用OSI-906或单用抗PD-1抗体,SCLC小鼠的肿瘤体积在21天后依然会不可控地疯长至约1000立方毫米。但当OSI-906与抗PD-1抗体联合出击时,肿瘤的生长轨迹被彻底改写,体积被死死压制在300立方毫米以下。
深层数据的分析揭示了联合治疗背后的动态重塑过程:OSI-906的加入显著削薄了血管的基底膜,使得曾经坚不可摧的BVG变得千疮百孔。伴随而来的是CD8+ T细胞数量的急剧攀升,从原本寥寥无几的20余个飙升至超过150个。同时,多重免疫荧光和单细胞转录组分析显示,肿瘤微环境中的巨噬细胞群体也发生了根本性的命运逆转,从具有免疫抑制功能的亚型(macro-c1)大量转化为具备抗肿瘤活性的状态(macro-c0和macro-c2)。
值得思考的是,这种干预方式微微减缓了肿瘤内部的血流速度,但并未改变肿瘤内部的酸碱度(pH)环境。在传统的抗血管生成疗法中,研究者致力于将过度渗漏的血管“正常化”,以改善化疗药物的递送;而在小细胞肺癌的语境下,这种“正常化”却意味着将过度紧密的BVG适度“破坏”,增加其渗漏性,从而为免疫细胞打开一扇大门。这种视角的转换,为克服肿瘤免疫耐药提供了全新的思路。放眼全局:泛神经内分泌肿瘤的通用护盾
小细胞肺癌凭借这套机制在免疫系统的眼皮底下暗度陈仓,那么其他部位的神经内分泌肿瘤是否也掌握了这门“建筑手艺”?神经内分泌肿瘤由于其特殊的起源,广泛分布于人体的各个器官。为了验证这一假设,研究团队将目光投向了食管、膀胱、胃、胰腺、直肠以及去势抵抗性前列腺(CRPC-NE)等多种组织来源的泛神经内分泌肿瘤(Pan-NECs)。
病理切片的测量数据给出了肯定的答案。这些泛神经内分泌肿瘤的血管基底膜厚度无一例外地超过了10微米,部分甚至达到了12到15微米;而与其对应的非神经内分泌肿瘤组织(如胃腺癌、尿路上皮癌等),其血管基底膜厚度仍维持在2到3微米左右的常态水平。同时,在这些部位的NECs中,同样能够检测到高强度的紧密连接蛋白和周细胞标志物。
进一步的基因组学队列分析证实,在这些泛神经内分泌肿瘤中,高水平的ASCL1表达同样严格对应着高水平的IGFBP5表达,并且内皮细胞屏障相关通路的活跃度与T细胞的浸润程度呈现出显著的负相关。
这表明,血脑屏障样血管门并非小细胞肺癌的专属奇招,而是整个神经内分泌肿瘤家族为了抵御免疫系统清剿,在进化过程中保留并共享的一种通用物理护盾。
这项研究为我们描绘了一幅令人震撼的肿瘤微环境图景。肿瘤并非仅仅是失控增殖的细胞团,它们更像是一个个精密运作的组织,懂得如何利用转录因子去遥控远端的正常内皮细胞,为自己搭建起一座抵御外敌的坚固堡垒。但科学的魅力就在于,每一道看似无解的屏障背后,都隐藏着被瓦解的命门。当我们将目光从单一的癌细胞转移到这堵隐形的“血管城墙”上时,或许,敲开神经内分泌肿瘤治愈之门的钥匙,就已经握在了我们手中。
参考文献
Wang Y, Zhong A, Wang B, Zhai X, Lei C, Liang Z, Deng X, Zhong J, Xiao C, Zheng J, Wu B, Zhang L, Wang Y, Luo X, Wang J, Zhang M, Liu H, Wan X, Dai S, Yang Y, Zhang S, Wang W, Yang S, Xue J, Zhao C, Tammela T, Li Z, Zhang Y, Na F, Wang M, Liu Y, Chen C. A blood-brain barrier-like vascular gate limits immunotherapy efficacy in neuroendocrine cancers. Cell. 2026 May 7:S0092-8674(26)00452-6. doi: 10.1016/j.cell.2026.04.017. Epub ahead of print. PMID: 42102817.
声明:本文仅用于分享,不代表平台立场,如涉及版权等问题,请尽快联系我们,我们第一时间更正,谢谢!
往期热文:
Nature Genetics | PCOS演化悖论:遗传学视角解析“平衡多效性”对女性生育窗口的代偿延长
Cell | 谁说mRNA出核必须走核孔?揭开内源性巨型RNA跨核膜的“出芽”之谜
Nature Methods | 荧光成像新纪元:抗原稳定型多色纳米抗体的全光谱突围
Nature | 还在用对侧脑区做“健康对照”?研究揭示星形胶质细胞跨半球网络带来的实验新雷区
Nature Medicine | 帕金森病的肠道倒计时:隐藏在微生态中的患病轨迹
Nature | 打破多倍体对称性!单体型基因组揭示“染色体融合”触发二倍体化
Cell | 从“帮凶”到“刺客”:研究揭示亲环素家族蛋白PPID阻断HIV核输入的全新机制
Nature Medicine | 从表观遗传到宏观流行病学:最新研究实锤农药暴露与肠癌年轻化的深层关联
Nature Methods | 被颠覆的序列之谜:让AI重新理解RNA语言的生命密码
Nature Genetics | 从遗传发掘到临床预警,高度近视高危人群筛选迎来新标尺
2026-05-09
·生物谷
研究揭示了蛋白质回收功能受损是T细胞耗竭的出人意料的罪魁祸首。
T细胞耗竭(T cell exhaustion)是慢性感染和肿瘤中常见的T细胞功能障碍状态。耗竭的T细胞增殖能力下降、效应分子(如IFN-γ、TNF-α)分泌减少,并高表达抑制性受体(如PD-1、CTLA-4、TIM-3)。虽然免疫检查点抑制剂(如抗PD-1抗体)可以部分逆转耗竭,但许多患者仍无法恢复完全的T细胞功能。蛋白质稳态(proteostasis)是指细胞内蛋白质合成、折叠、运输和降解的平衡状态。其中,“泛素-蛋白酶体系统”负责标记和降解受损或错误折叠的蛋白质,而E3泛素连接酶是这一系统中的“标签工”——它们特异性地识别靶蛋白并为其添加泛素链,从而引导蛋白被降解。
T细胞是我们免疫系统的关键组成部分,是对抗感染和疾病的重要保护者。但它们的防御能力是有限的。T细胞并非取之不尽的保护者。通常,当T细胞参与杀伤肿瘤细胞后,它们会“燃尽”自己,陷入一种称为T细胞耗竭的功能障碍状态。从耗竭中恢复——或者完全避免耗竭——将极大地有助于它们战胜疾病。如今,将T细胞从“燃尽”中拯救出来并使它们恢复抗癌状态,是癌症免疫治疗领域的一个主要焦点。
加州大学圣地亚哥分校生物科学学院Ananda Goldrath教授实验室在《Cell》期刊上发表的新研究,在我们目前对T细胞耗竭的理解中发现了一个意想不到的因素。
蛋白质稳态是一个由细胞过程组成的网络,负责协调细胞内蛋白质的正确构建、运动和破坏。该网络的一个组成部分是细胞内的一种回收功能。健康细胞会持续拆解老旧和受损的蛋白质,以保存能量并重复利用构件来制造新蛋白质。
这项由Goldrath实验室的博士后研究员Nicole Scharping领导,并包括拉霍亚免疫学研究所和阿肯色大学医学科学家的研究(该研究还被选为《Cell》封面文章),揭示了蛋白质回收功能受损是T细胞耗竭的出人意料的罪魁祸首。
Scharping说:“我们发现,耗竭T细胞的回收程序正在崩溃,导致受损和错误折叠的蛋白质堆积起来,无处可去。”
但该研究还发现了一种通过修复破损的回收功能并恢复正常蛋白质稳态来逆转错误折叠蛋白积累的方法。Scharping说,这个问题可以通过“标记和分拣”修复来解决。一个称为E3连接酶的酶家族在回收设施中充当“贴标签者”,给老旧蛋白贴上标签,以便细胞知道要分解它们。
Scharping说:“在耗竭的T细胞中,许多这些酶被关闭,回收工作停滞。当我们恢复特定的E3连接酶时,堆积物被清除,T细胞恢复了功能,在清除肿瘤方面表现更好。”
研究人员指出,蛋白质加工受损并非功能失调的T细胞所独有。这一发现提示,蛋白质回收受损可能将原本不相关的疾病的生物学过程联系在一起。
Goldrath(分子生物学系教授)说:“我们认为这种蛋白质稳态的丧失类似于帕金森病和阿尔茨海默病等蛋白质聚集疾病中神经元发生的情况。将这些细胞从耗竭中拯救出来,可以改善T细胞对慢性感染和肿瘤的反应能力。”
这一发现得益于质谱技术的应用——一种鉴定和测量分子质量的先进分析方法,由加州大学圣地亚哥分校(细胞与发育生物学系)Eric Bennett教授实验室和拉霍亚免疫学研究所全球自身免疫研究所助理教授Samuel Myers实验室领导。质谱技术提供了同时检查数千种蛋白质的能力,并提示NEURL3、RNF149和WSB1是负责恢复T细胞回收功能的E3连接酶。
当这些连接酶被恢复、蛋白质积累的阻滞被清除后,研究人员发现他们可以提高细胞疗法对抗癌症的效力。虽然这项新研究是在小鼠中进行的,但研究人员指出,类似的策略可应用于人类癌症的免疫治疗。
Goldrath说:“我们揭示了T细胞失去抗癌能力的一种机制,现在我们有了一个靶点,可以用来设计干预措施,改善对免疫治疗的反应,并预防慢性感染。”(生物谷Bioon.com)
参考文献:
Nicole E. Scharping et al, Proteostasis sustains T cell differentiation potential and tumor-infiltrating lymphocyte function, Cell (2026). DOI: 10.1016/j.cell.2026.02.019.
免疫疗法蛋白降解靶向嵌合体
2026-04-24
·知乎专栏
结直肠癌治疗失败,常因癌细胞变得‘刀枪不入’——无论药物外排、修复损伤,还是与周围环境‘结盟’,多重机制共同编织了一张致密的耐药网络。摘要结直肠癌是全球范围内高发病率和高死亡率的恶性肿瘤,化疗是其综合治疗的核心手段,但化疗耐药性的产生是导致治疗失败和患者预后不良的关键因素。目前,尽管对耐药机制的研究不断深入,涉及药物外排、DNA 修复、凋亡逃逸、肿瘤微环境、表观遗传、肿瘤干细胞、自噬、代谢重编程及信号通路异常等多个层面,但如何系统整合这些知识并转化为有效的临床干预仍是重大挑战。本文旨在系统综述结直肠癌化疗耐药的主要分子机制,并深入探讨当前及未来潜在的临床应对策略,包括新型靶向药物、联合治疗、纳米递送系统、免疫治疗联合、耐药逆转剂、液体活检监测、个体化治疗及多学科管理,以期为克服耐药、提高疗效提供理论依据与实践方向。关键词:结直肠癌;化疗耐药;分子机制;多药耐药;靶向治疗;联合策略;肿瘤微环境;个体化医疗前言结直肠癌是全球范围内发病率和死亡率均位居前列的恶性肿瘤,其治疗严重依赖于以 5-氟尿嘧啶(5-FU)、奥沙利铂和伊立替康为基础的化疗方案。然而,无论是原发性还是获得性化疗耐药,都已成为制约临床疗效、导致治疗失败和患者预后不良的核心挑战。化疗耐药的形成并非单一因素所致,而是一个涉及肿瘤细胞自身特性、肿瘤微环境(TME)以及二者之间复杂交互作用的动态、多步骤过程。近年来,随着基因组学、蛋白质组学、代谢组学以及单细胞测序等技术的飞速发展,我们对结直肠癌化疗耐药机制的理解正不断深化,从分子到系统层面揭示了其复杂性,这为开发新型的逆转耐药策略奠定了坚实的理论基础。在肿瘤细胞自身层面,多种内在机制共同驱动了耐药性的产生。癌症干细胞(CSCs)被认为是肿瘤复发、转移和耐药的关键驱动细胞亚群。研究表明,具有干细胞特性的细胞,如表达 LGR5 或 CLU 的细胞,能够通过维持静息状态或表型可塑性来逃避化疗药物的杀伤,并在治疗后重新填充肿瘤。例如,ZEB2 阳性的静息/慢周期细胞群体在化疗前就已存在,并具有预定的间充质样耐药表型,化疗后该群体扩增,导致治疗无应答。此外,上皮-间充质转化(EMT)过程与干细胞特性及耐药性紧密关联。转录因子如 ZEB2 和 SNAI1 不仅能诱导 EMT,还能通过激活核苷酸切除修复(NER)通路(如上调 ERCC1)或调控鞘氨醇激酶 2(SPHK2)信号,分别增强对奥沙利铂和 5-FU 的抵抗。代谢重编程是另一个关键耐药机制。肿瘤细胞通过改变能量获取方式,例如增强糖酵解(由 SIAH2/WNK1 轴驱动)或脂肪酸β-氧化(由 FOXA2/ALDOB 轴介导),来适应化疗压力并维持生存。过氧化物酶体膜蛋白 PMP70 则通过维持脂质代谢稳态和抵抗脂质过氧化,在奥沙利铂耐药中发挥保护作用。同时,异常的细胞信号通路,如持续活化的 AKT 信号(受 HSP47/PHLPP1 轴调控)、Wnt/β-catenin 通路以及 CD45-Wnt 信号轴,均被证实能促进干细胞特性并抑制化疗诱导的细胞凋亡,从而赋予耐药性。肿瘤微环境在塑造化疗耐药性方面扮演着同等重要的角色。癌症相关成纤维细胞(CAFs)是 TME 中最主要的间质细胞成分,它们通过分泌多种因子,特别是外泌体,与肿瘤细胞进行广泛对话。CAF 来源的外泌体携带蛋白质、脂质、microRNAs(miRNAs)、长链非编码 RNAs(lncRNAs)等货物,能够将耐药表型传递给肿瘤细胞,通过改变 DNA 损伤反应、铁死亡、凋亡和氧化应激等途径,重塑肿瘤细胞对 5-FU、奥沙利铂和伊立替康等药物的反应。肿瘤相关巨噬细胞(TAMs)的极化状态也影响耐药,例如,BRD4 通过上调 PAI-1 表达促进 M2 样 TAM 极化,进而导致奥沙利铂耐药。此外,肠道微生物群的失调,如具核梭杆菌的富集,也被发现可通过激活 Wnt/β-catenin 等信号通路,促进结直肠癌的发生和化疗耐药。在分子水平上,非编码 RNA(ncRNA)构成了一个精细而复杂的调控网络。MicroRNAs(如 miR-488、miR-33a-3p、miR-224-5p)可通过靶向 PFKFB3、EphA2、Bcl-2 等基因,分别影响糖酵解、细胞信号转导和自噬,从而调节化疗敏感性。长链非编码 RNAs(如 LncRNA-HMG、SNHG15)和环状 RNAs(circRNAs)则通过充当分子海绵、调控转录或影响蛋白稳定性等方式参与耐药。例如,LncRNA-HMG 通过结合并促进 p53 降解,抑制铁死亡,进而导致化疗耐药。表观遗传学改变,包括 DNA 甲基化、组蛋白修饰等,也在不改变 DNA 序列的情况下持续影响基因表达,驱动耐药表型的产生和维持。面对多维度、多层次的耐药机制,开发有效的应对策略至关重要。传统的药物再利用策略展现出潜力,例如抗蠕虫药阿苯达唑通过靶向泛素连接酶 RNF20,能够抑制结直肠癌进展并使耐药细胞对 5-FU 和奥沙利铂重新敏感。针对特定耐药通路的靶向干预是另一主要方向,包括使用 Aurora B 激酶抑制剂(AZD1152)增强 5-FU 疗效、使用 Rac1 抑制剂(1A-116)或 SPHK2 抑制剂(ABC294640)逆转 5-FU 耐药、以及通过抑制 GCLC 或靶向 PMP70 来克服奥沙利铂耐药。针对肿瘤干细胞和 EMT 过程的策略,如抑制 PIK3C3/VPS34(使用 36-077)或靶向 Claudin-1/EPHA2 相互作用,也被证明能有效增敏化疗。此外,调节肿瘤微环境,例如改变 CAF 功能或 TAM 极化,以及利用纳米技术实现药物的靶向递送和控释,为提高药物在肿瘤部位的积累、克服耐药提供了新思路。随着精准医学的发展,基于患者肿瘤的分子特征(如 CMS 分型、RAS/BRAF 突变状态、左右半结肠起源)和新型生物标志物(如 ANXA1、ENY2、MSLN)进行个体化治疗选择,正成为优化临床决策、逆转或避免耐药的关键。尽管抗 EGFR 疗法在右半结肠癌中通常疗效有限,但个别案例显示,在严格筛选的 RAS 野生型患者中,后期线使用仍可能获益,提示需要更精细的生物标志物来指导患者分层。综上所述,结直肠癌的化疗耐药是一个由肿瘤细胞内在变异与外部微环境信号共同编织的复杂网络。全面解析这些机制,并在此基础上发展多靶点、多层次、个体化的综合治疗策略,是未来克服耐药、改善结直肠癌患者临床预后的必由之路。1. 药物转运蛋白介导的多药耐药机制1.1 ATP 结合盒转运蛋白超家族的作用ATP 结合盒转运蛋白超家族是介导结直肠癌多药耐药的核心机制之一,其成员通过水解 ATP 提供的能量,主动将多种化疗药物泵出细胞外,导致细胞内药物浓度降低,从而引发治疗失败。其中,ABCB1(P-糖蛋白)的过表达是经典的多药耐药机制。研究表明,ABCB1 能主动外排包括伊立替康活性代谢产物 SN-38 在内的多种化疗药物。其表达受多种因素调控,例如,胰岛素样生长因子 2 mRNA 结合蛋白 3 作为 m6A 阅读器,可直接结合 ABCB1 mRNA 的 m6A 修饰区域,从而促进其 mRNA 的稳定性与表达,进而赋予结直肠癌细胞耐药性。此外,Hedgehog-GLI 信号通路可通过转录因子 GLI1 直接结合 ABCB1 的启动子区域,上调其表达,促进对 5-氟尿嘧啶和奥沙利铂的耐药。针对 ABCB1 的抑制剂,如 EGFR 抑制剂 sapitinib,可通过与 ABCB1 的流出位点相互作用,增加细胞内紫杉醇的积累,从而在体外逆转耐药。除 ABCB1 外,ABCC 家族(如 MRP1)和 ABCG2(BCRP)也广泛参与结直肠癌的化疗耐药。这些转运蛋白能介导对 5-氟尿嘧啶、奥沙利铂、伊立替康等多种药物的外排。例如,在 5-氟尿嘧啶耐药的结直肠癌细胞中,MRP1 和 ABCG2 的表达常显著上调。其表达调控网络复杂,涉及多种信号通路和转录因子。研究发现,内质网应激感应器 IRE1α可通过剪接 XBP1 mRNA 生成活性转录因子 XBP1s,进而诱导 ABCB1、ABCC1 和 ABCG2 的表达,而抑制 IRE1α的 RNase 活性则可下调这些转运蛋白并增敏耐药细胞。此外,PI3K/Akt/mTOR 信号通路的激活也被证实可上调 ABCG2 和 ABCC5 的表达,从而限制 5-氟尿嘧啶的疗效。非编码 RNA,如某些 miRNAs,也被发现可通过靶向调控这些转运蛋白的表达来影响耐药性。最新研究进一步揭示了转运蛋白的翻译后修饰及其与细胞微环境的相互作用在调节其活性和稳定性中的关键作用。例如,蛋白精氨酸甲基转移酶 3 可通过精氨酸甲基化修饰 Y-box 结合蛋白 1,影响其相分离与核转位,进而转录调控 ABC 转运蛋白的表达,导致直肠癌放化疗抵抗。此外,转运蛋白的活性也受到其与细胞膜脂筏相互作用的调节。虽然具体机制在结直肠癌中的研究尚在深入,但已知 ABC 转运蛋白的功能依赖于其正确的膜定位和构象变化。一些天然化合物,如蛇毒来源的 L-氨基酸氧化酶,被证实具有抑制 P-糖蛋白的潜力,提示其可能通过影响转运蛋白的活性或膜环境来克服耐药。同样,植物黄酮醇异鼠李素也能显著降低结直肠癌细胞中 P-gp、MRP1 和 MRP5 的 mRNA 和蛋白水平,其分子对接显示与这些蛋白有高结合亲和力,可能干扰其功能。这些发现为开发针对转运蛋白功能调控而非单纯表达抑制的新型逆转耐药策略提供了新视角。1.2 调控药物内流与分布的机制药物摄入转运体功能的下调是结直肠癌细胞产生化疗耐药的重要机制之一。以 5-氟尿嘧啶(5-FU)为例,其前体药物需要依赖特定的膜转运蛋白进入细胞内才能发挥细胞毒性作用。研究表明,人平衡型核苷转运体 1(hENT1)是介导 5-FU 类似物吉西他滨等核苷类化疗药物进入细胞的关键蛋白。当这些转运体的表达或功能下调时,会直接导致细胞内药物浓度不足,从而引发耐药。例如,在结直肠癌中,ATP 结合盒(ABC)转运蛋白家族的过度表达是导致多药耐药(MDR)的经典途径,这些蛋白能将多种化疗药物泵出细胞外,降低其胞内有效浓度。除了经典的 ABC 转运蛋白,其他类型的转运体也参与其中。例如,有研究指出,脂质转运蛋白如 ABCA 亚家族成员的表达失调与结直肠癌进展和耐药相关,它们通过改变细胞膜脂质组成或影响药物在细胞内的微环境来间接影响药物敏感性。因此,靶向这些药物摄入转运体,恢复其功能或抑制外排泵活性,是克服此类耐药策略的重要研究方向。除了药物摄入,药物在细胞内的分布和靶向定位异常也是导致耐药的关键环节。以奥沙利铂为例,其作用机制是与 DNA 形成铂-加合物,从而诱导细胞凋亡。然而,药物能否有效到达细胞核内的 DNA 靶点,受到细胞内转运和分布过程的严格调控。研究发现,铜转运蛋白(如 CTR1)在奥沙利铂的摄取和细胞内分布中扮演重要角色。这些转运蛋白的异常表达或功能丧失,会直接影响奥沙利铂进入细胞并转运至作用位点的效率。此外,细胞内药物分布的改变还可能涉及细胞器水平的隔离。例如,过氧化物酶体膜蛋白 PMP70 被发现在奥沙利铂耐药的结直肠癌细胞中高表达,并可能通过调节脂质过氧化等代谢途径,间接影响细胞对药物的反应和存活。这种细胞器功能的改变,使得药物无法在正确的位置达到有效的杀伤浓度。同时,细胞内药物解毒系统的激活,如谷胱甘肽 S-转移酶(GST)等酶的表达上调,可以加速药物代谢失活,也是影响药物最终到达 DNA 靶点效率的重要因素。因此,深入理解并干预影响化疗药物细胞内分布和定位的分子机制,对于破解奥沙利铂等药物的耐药性具有重要临床意义。2. DNA 损伤修复系统的异常激活2.1 错配修复与核苷酸切除修复通路错配修复系统缺陷是结直肠癌发生发展及治疗反应异质性的关键分子机制之一。该系统主要由 MLH1、MSH2、MSH6 和 PMS2 等基因编码的蛋白构成,其功能失活会导致 DNA 复制过程中产生的碱基错配无法被有效纠正,进而引发微卫星不稳定性。MSI-H/dMMR 表型在结直肠癌中约占 15%,且与特定的临床病理特征相关,例如更常见于右半结肠、黏液腺癌或低分化癌。尽管 dMMR 肿瘤因其高突变负荷和丰富的肿瘤新抗原而对免疫检查点抑制剂表现出极高的敏感性,在晚期和局部晚期疾病中均能诱导显著的病理完全缓解,但其对传统化疗,特别是以氟尿嘧啶为基础的方案,可能存在原发性耐药。研究显示,接受新辅助化疗的 dMMR 直肠癌患者中,有相当比例(如 29%)出现疾病进展,而 pMMR 患者则无此现象,这提示 MMR 状态是预测化疗反应的重要生物标志物。这种耐药性可能与 dMMR 肿瘤细胞因基因组不稳定而累积的获得性突变有关,这些突变可能激活其他促生存或药物外排通路,从而抵消特定化疗药物的细胞毒性作用。因此,在临床实践中,对所有结直肠癌进行普遍的 MMR 状态检测至关重要,这不仅有助于识别林奇综合征患者,指导遗传咨询和家系管理,更是制定个体化治疗方案(如选择免疫治疗而非传统化疗)的基础。核苷酸切除修复通路是细胞应对庞大体积 DNA 加合物损伤的核心修复机制,其关键因子 ERCC1 的表达水平与以奥沙利铂为代表的铂类药物的疗效密切相关。奥沙利铂通过形成 DNA 链内和链间铂加合物,干扰 DNA 复制与转录,最终诱导细胞凋亡。NER 通路,特别是其中的 ERCC1-XPF 核酸内切酶复合物,能够识别并切除这些铂加合物,完成损伤 DNA 的修复。大量研究表明,肿瘤细胞中 ERCC1 的过表达会增强其对 DNA 铂加合物的修复能力,从而降低奥沙利铂的细胞毒性,导致临床耐药。因此,ERCC1 已被广泛认为是预测结直肠癌患者对奥沙利铂为基础化疗方案敏感性的负向生物标志物。在错配修复功能正常的 pMMR 结直肠癌中,NER 通路的活性状态对化疗结局的影响尤为显著。除了 ERCC1,NER 通路其他组分如 XPD 的多态性也可能影响修复效率,进而与肿瘤易感性和治疗反应差异相关联。值得注意的是,DNA 修复通路并非孤立运作,NER 与 MMR 等其他修复途径之间存在交叉对话,共同维持基因组稳定性。例如,在应对某些类型的 DNA 损伤时,这些通路可能协同或互补,而通路的缺陷或功能亢进则共同构成了肿瘤细胞复杂耐药网络的重要组成部分。深入解析 ERCC1 等 NER 关键因子的调控机制,并探索其抑制剂,是克服铂类耐药、改善 pMMR 结直肠癌患者预后的重要研究方向。2.2 同源重组修复与跨损伤合成同源重组修复是细胞应对 DNA 双链断裂的关键机制,其功能完整性对维持基因组稳定至关重要。在结直肠癌中,同源重组修复相关基因的突变或功能改变,特别是 BRCA1/2 等基因的缺陷,会严重影响细胞对化疗药物(如铂类和拓扑异构酶抑制剂)造成的 DNA 损伤的应答能力。这种缺陷不仅与 PARP 抑制剂的敏感性相关,也深刻影响传统化疗的效果。例如,研究发现携带同源重组修复基因突变的结直肠癌患者在接受化疗后,其肿瘤转移的可能性更低,且总生存期显著延长,这表明同源重组修复缺陷可能是一个预测化疗疗效的有利生物标志物。从机制上看,同源重组修复功能的丧失会导致 DNA 损伤修复能力下降,迫使肿瘤细胞更依赖于其他易错的修复途径,从而增加其对 DNA 损伤类化疗药物的敏感性。此外,同源重组修复通路中的关键蛋白如 RAD51 的功能也受到精细调控。例如,细胞骨架蛋白 smoothelin 被发现在 DNA 损伤后迅速募集至损伤位点,通过稳定 RAD51 蛋白、防止其被 E3 泛素连接酶 HUWE1 降解,从而促进同源重组修复的进行。反之,抑制 smoothelin 则会加剧 DNA 损伤并增强 CRC 细胞对伊立替康等药物的敏感性。这些研究揭示了同源重组修复通路在结直肠癌化疗耐药中的核心地位,靶向该通路的关键组分(如 RAD51)或利用其缺陷状态(如同源重组修复突变)已成为克服耐药的新策略。跨损伤合成是细胞在 DNA 复制过程中绕过化疗药物造成的损伤位点的一种易错修复机制,由专门的跨损伤合成聚合酶(如 Pol ζ、Pol κ等)执行。在结直肠癌中,跨损伤合成的异常激活是驱动化疗耐药和肿瘤进化的重要因素。研究表明,跨损伤合成关键组分 REV7(Pol ζ的组成部分)的表达在 5-氟尿嘧啶和奥沙利铂耐药的 CRC 细胞中显著上调。这种上调增强了细胞的跨损伤合成能力,允许 DNA 复制机器绕过药物诱导的损伤,从而促进细胞存活并导致突变积累,最终产生耐药克隆。实验证实,利用 CRISPR/Cas9 技术敲除 REV7 能显著逆转 CRC 细胞对 5-氟尿嘧啶和奥沙利铂的耐药性,并在小鼠移植瘤模型中显示出强大的协同抗肿瘤效应。此外,其他跨损伤合成聚合酶也参与其中。例如,长链非编码 RNA H19 在缺氧或奥沙利铂处理的 CRC 细胞中表达增强,并通过作为 miR-675-3p 的竞争性内源 RNA 来调节上皮-间质转化,从而促进奥沙利铂耐药。另一个例子是,RIP140 转录因子被发现能正向调节编码跨损伤合成聚合酶κ的 POLK 基因表达。在 CRC 细胞中,RIP140 的缺失会降低 POLK 水平,增加细胞对烷化剂(如甲基磺酸甲酯)的敏感性,这表明跨损伤合成聚合酶κ在应对特定 DNA 损伤中发挥作用。这些机制共同表明,跨损伤合成通路为 CRC 细胞在化疗压力下提供了生存优势,并成为基因组不稳定的来源。因此,靶向跨损伤合成通路的关键分子,如 REV7 或调控其表达的因子,为逆转结直肠癌的化疗耐药提供了极具前景的新方向[PCSJTJJWX]。3. 细胞凋亡与自噬途径的失调3.1 凋亡通路关键分子的改变在结直肠癌化疗耐药的形成过程中,凋亡通路关键分子的改变扮演了核心角色。抗凋亡蛋白的过表达与促凋亡蛋白的功能抑制共同作用,显著提高了肿瘤细胞的凋亡阈值,使其能够在化疗药物的攻击下存活。例如,Bcl-2 家族蛋白的失衡是导致凋亡抵抗的关键机制之一。研究表明,在缺氧环境下,结直肠癌细胞中 Bcl-2 的表达显著上调,这直接导致了细胞对化疗药物的抵抗。α-hederin 能够通过抑制 AKT/Bcl2 信号通路,下调 Bcl-2 的表达,从而逆转缺氧介导的结直肠癌耐药。此外,其他抗凋亡蛋白如 Bcl-xL 和 Mcl-1 也参与了耐药过程。去泛素化酶 USP49 通过 C-MYC-USP49-BAG2 轴稳定 BAG2 蛋白,而 BAG2 是已知的抗凋亡蛋白,能够拮抗细胞凋亡,从而促进结直肠癌细胞的存活和化疗抵抗。另一方面,促凋亡蛋白如 Bax 的功能受到抑制。研究发现,分泌型黏蛋白 MUC5AC 通过 MUC5AC-CD44 轴激活 Src 信号,并导致 p53 及其靶基因 p21 的下调,同时上调β-catenin 及其靶基因 CD44 和 Lgr5,这一系列变化不仅增强了肿瘤发生,还通过下调 p53 通路削弱了促凋亡信号,从而赋予细胞对 5-氟尿嘧啶和奥沙利铂的耐药性。p53 肿瘤抑制基因的突变或功能丧失在超过 50% 的结直肠癌中存在,是核心的耐药机制之一。p53 的失活导致细胞周期检查点失效和凋亡诱导能力急剧下降。例如,长链非编码 RNA SNHG15 在结直肠癌中表达上调,其过表达能够促进细胞增殖、5-氟尿嘧啶耐药和糖酵解,并上调 BCL2 的表达,从而抑制凋亡。同样,miR-17-5p 通过 METTL14 依赖的成熟过程被上调,进而靶向下调 Mitofusin 2 (MFN2),破坏线粒体稳态,最终导致 5-氟尿嘧啶化疗抵抗,这一过程也涉及凋亡通路的抑制。这些凋亡通路关键分子的系统性改变,共同构建了一个有利于肿瘤细胞在化疗压力下存活的微环境,深刻影响了结直肠癌的治疗效果。3.2 自噬的双重角色与调控自噬在结直肠癌化疗耐药中扮演着复杂且具有高度情境依赖性的双重角色。一方面,化疗药物如奥沙利铂和 5-氟尿嘧啶(5-FU)能够诱导保护性自噬,帮助肿瘤细胞应对药物应激,从而促进耐药性的产生。这种保护性自噬通过清除受损的细胞器,为细胞提供能量和底物,维持其生存。例如,研究发现,在奥沙利铂耐药的结直肠癌细胞中,miR-135b-5p 表达上调,通过靶向 MUL1/ULK1 通路诱导保护性自噬,进而促进耐药。类似地,肿瘤相关成纤维细胞(CAFs)来源的条件培养基能够上调结直肠癌细胞的自噬水平,从而降低其对 5-FU 的敏感性,这种效应可被自噬抑制剂 3-甲基腺嘌呤(3-MA)所逆转。基于此,使用自噬抑制剂如氯喹(CQ)或 3-MA 在临床前模型中显示出显著的增敏效果。例如,在微卫星高度不稳定(MSI-H)的结直肠癌细胞中,联合使用 5-FU 和酪蛋白激酶 1-α(CK1α)抑制剂 D4476 能够抑制自噬流,从而增强 5-FU 的细胞毒性。此外,中药活性成分如藤黄酸、重楼皂苷 A 等也被证实可通过调控 AMPK/mTOR 等信号通路诱导自噬,进而抑制结直肠癌细胞增殖或促进其凋亡。这些研究共同表明,靶向抑制保护性自噬是克服化疗耐药的一个潜在策略。然而,自噬在结直肠癌中的作用并非总是保护性的,其具体角色高度依赖于肿瘤类型、发展阶段、药物背景以及肿瘤微环境等多种因素。在某些情况下,过度或持续的自噬可能超过细胞的承受能力,导致 II 型程序性细胞死亡,即自噬性细胞死亡。这种促死亡的自噬作用机制复杂,且与保护性自噬的调控网络存在交叉。例如,研究发现,细胞周期蛋白依赖性激酶 9(CDK9)的表达与结直肠癌组织中自噬水平呈正相关,且高表达 CDK9 提示预后不良。这表明自噬的激活可能与肿瘤的恶性进展相关。此外,肠道菌群如具核梭杆菌(Fusobacterium nucleatum)可通过调节 Hippo 通路等方式抑制化疗药物诱导的细胞焦亡,从而介导耐药,这一过程也可能涉及自噬通路的改变。另一方面,长链非编码 RNA H19 在缺氧条件下表达上调,通过作为 miR-675-3p 的竞争性内源 RNA 调控上皮-间质转化(EMT),从而促进奥沙利铂耐药,而抑制 H19 表达则可改善缺氧诱导的耐药性。这些研究揭示了自噬调控网络的复杂性,其最终效应是促进生存还是导致死亡,取决于多种信号通路的整合与平衡。因此,深入理解自噬在特定肿瘤背景下的精确调控机制,对于制定有效的治疗策略至关重要。4. 肿瘤微环境在化疗耐药中的关键作用4.1 癌症相关成纤维细胞的相互作用癌症相关成纤维细胞(CAFs)是结直肠癌(CRC)肿瘤微环境(TME)中最关键的基质细胞之一,其通过多种机制与肿瘤细胞相互作用,共同驱动肿瘤进展并介导化疗耐药。CAFs 通过分泌多种细胞因子、外泌体及重塑细胞外基质(ECM),构建了一个物理和生化屏障,严重阻碍了化疗药物的渗透并传递促生存信号。例如,CAFs 能够分泌肝细胞生长因子(HGF),通过旁分泌作用激活肿瘤细胞上的 MET 受体,从而促进 CRC 细胞的迁移和侵袭。此外,CAFs 来源的白细胞介素-6(IL-6)在介导奥沙利铂耐药中扮演了核心角色。研究表明,CAFs 与 CRC 细胞共培养或经奥沙利铂刺激后,其分泌的 IL-6 水平显著升高,而 CAFs 来源的条件培养基能够增强 CRC 细胞在奥沙利铂处理下的存活率,这一效应可被 IL-6 中和抗体所逆转。除了细胞因子,CAFs 衍生的外泌体也是传递耐药信号的重要载体。例如,CAFs 来源的外泌体 miR-181b-3p 能够被 CRC 细胞摄取,通过下调分选连接蛋白 2(SNX2)的表达,从而促进 CRC 细胞的增殖、迁移并抑制其凋亡。同时,CAFs 通过分泌和重塑 ECM,形成致密的纤维化间质,这不仅为肿瘤细胞提供了物理屏障,其本身也直接参与耐药。研究发现,由 CAFs 生成的 ECM 能够赋予 CRC 细胞对奥沙利铂和西妥昔单抗的抵抗能力,而 SNAI1 在 CAFs 中的表达是调控这一过程的关键。这种致密的 ECM 结构在单细胞测序分析中被证实与 CRC 肝转移灶中 ECM 重塑/促血管生成型 CAF 亚群密切相关,这些 CAF 亚群与强烈的促结缔组织增生反应和 Wnt 信号通路激活区域共存。CAFs 诱导的关键信号通路激活,特别是 Wnt/β-catenin 通路,是微环境介导耐药的核心机制,直接增强了肿瘤细胞的干性并抑制其凋亡。在 CRC 中,CAFs 高表达 Wnt5a,其状态与肿瘤大小、浸润深度、淋巴结转移和复发显著相关。Wnt5a 来源于 CAFs,能够显著刺激 CRC 细胞的增殖和迁移,是 CRC 进展的关键驱动因子。除了经典的 Wnt 配体,CAFs 来源的外泌体也能激活 Wnt/β-catenin 信号。例如,外泌体携带的 miR-92a-3p 被证实可以激活 CRC 细胞中的 Wnt/β-catenin 通路,从而维持肿瘤干性。这种对干性通路的调控是 CAFs 促进肿瘤恶性进展和耐药的重要方式。此外,CAFs 还能通过其他信号轴影响肿瘤细胞。例如,CAFs 中脂质代谢的重编程导致脂肪酸合成酶(FASN)表达上调,其分泌的脂质代谢物被 CRC 细胞通过 CD36 受体摄取,从而增强 CRC 细胞的迁移能力,这为肿瘤转移和可能的生存优势提供了新的代谢视角。CAFs 的异质性进一步复杂化了其功能,已鉴定出包括肌成纤维细胞型 CAF(myCAF)、炎症型 CAF(iCAF)、抗原呈递型 CAF(apCAF)以及新近发现的 T 细胞抑制型 CAF(TinCAF)等多种亚型。其中,iCAF 亚型以分泌大量 IL-6、CXCL1 等炎症因子为特征,而梭杆菌感染可以诱导 CAFs 表现出显著的 iCAF 表型,进而与癌细胞相互作用增强其侵袭能力。TinCAF 则通过表达 NECTIN2 等免疫抑制分子,直接抑制效应 T 细胞的功能,促进免疫逃逸。综上所述,CAFs 通过构建物理屏障、分泌可溶性因子和外泌体、以及激活关键促生存和干性通路,形成了一个多维度的耐药网络。靶向 CAFs 或其分泌的关键因子(如 IL-6、Wnt5a、外泌体生物发生)或破坏其介导的免疫抑制微环境,已成为克服 CRC 化疗耐药极具前景的策略。4.2 免疫细胞与细胞因子的影响肿瘤微环境中的免疫细胞与细胞因子在结直肠癌化疗耐药的形成中扮演着复杂而关键的角色。肿瘤相关巨噬细胞,尤其是 M2 型巨噬细胞,是构成免疫抑制微环境的核心力量。这些细胞通过分泌多种免疫抑制因子,如转化生长因子-β和白细胞介素-10,直接抑制效应 T 细胞的功能,从而削弱机体对肿瘤的免疫监视和攻击能力。这种免疫抑制状态不仅促进了肿瘤的免疫逃逸,还间接导致了化疗耐药,因为有效的抗肿瘤免疫反应对于清除化疗后残留的肿瘤细胞至关重要。此外,M2 型巨噬细胞还能分泌血管内皮生长因子等促血管生成因子,促进肿瘤血管新生,为肿瘤细胞提供营养并可能降低化疗药物的递送效率,进一步加剧耐药。在结直肠癌中,肿瘤相关巨噬细胞的浸润与患者的不良预后密切相关,其通过重塑肿瘤微环境,为肿瘤细胞创造了有利于生存和抵抗治疗的庇护所。同时,其他免疫抑制细胞,如调节性 T 细胞和髓系来源的抑制性细胞,也通过类似的机制协同作用,共同营造一个高度免疫抑制的局部环境,使得化疗药物难以发挥最大疗效。化疗本身作为一种强烈的细胞应激,可能触发一系列复杂的生物学反应,其中损伤相关分子模式的释放是一个重要环节。化疗药物导致肿瘤细胞死亡时,会释放出包括高迁移率族蛋白 B1、ATP 等在内的损伤相关分子模式,这些分子作为危险信号,可以激活模式识别受体,如 Toll 样受体,从而触发强烈的炎症反应。然而,这种由治疗引发的炎症反应具有双刃剑效应。一方面,它可能招募并激活免疫细胞,试图清除肿瘤;但另一方面,在结直肠癌的特定微环境中,持续的炎症反应反而可能促进肿瘤的再生和转移。例如,化疗后释放的炎症因子,如白细胞介素-6 和白细胞介素-8,可以激活下游的 STAT3 和 NF-κB 等信号通路,这些通路与肿瘤细胞的增殖、存活、上皮-间质转化以及干细胞特性的维持密切相关,从而助长了“化疗后复发”的恶性循环。研究表明,在结直肠癌患者中,外周血中某些促炎细胞因子水平的升高与疾病进展和不良预后相关。更值得注意的是,这种治疗诱导的炎症微环境可能进一步招募和极化为 M2 型的肿瘤相关巨噬细胞,形成一个自我强化的促肿瘤、促耐药的正反馈环路。因此,深入理解化疗如何通过影响免疫细胞和细胞因子来重塑肿瘤微环境,对于打破耐药循环、开发新的联合治疗策略具有至关重要的意义。5. 表观遗传学调控与耐药5.1 DNA 甲基化与组蛋白修饰DNA 甲基化是结直肠癌(CRC)中研究最为深入的表观遗传机制之一,其异常模式在化疗耐药的形成中扮演着关键角色。具体而言,启动子区域的异常高甲基化是导致关键抑癌基因和药物代谢基因转录沉默的重要途径。例如,在 CRC 中,APC、p16 等经典抑癌基因的启动子常因高甲基化而失活,破坏了正常的细胞周期调控和凋亡通路,从而赋予癌细胞生存优势。此外,药物代谢相关基因的沉默也会直接影响化疗药物的细胞内浓度和效力。一项大规模甲基化组学分析发现,在从正常黏膜到腺瘤再到腺癌的演进序列中,细胞外基质(ECM)组织相关基因(如 ITGA4、FBN1、COL4A1、COL4A2)的启动子区发生了显著的高甲基化,并与其表达水平呈负相关。ECM 功能的破坏与肿瘤侵袭、免疫逃逸和耐药性密切相关,这提示 DNA 甲基化介导的基因沉默是驱动 CRC 进展和耐药的一个核心表观遗传程序。另一方面,基因组整体的低甲基化状态同样不容忽视,它可能导致原癌基因的异常激活和转座子元件的去抑制,从而加剧基因组的不稳定性,为癌细胞的进化与适应化疗压力提供了遗传基础。这种高甲基化与低甲基化并存的“甲基化失衡”现象,共同构成了 CRC 耐药性的复杂表观遗传景观。组蛋白修饰是另一类至关重要的表观遗传调控方式,通过改变染色质状态全局性影响基因表达谱,在 CRC 耐药中作用显著。组蛋白修饰酶,如组蛋白甲基转移酶 EZH2 和组蛋白去乙酰化酶(HDACs)的异常表达,是重塑耐药相关染色质环境的核心驱动因素。EZH2 作为多梳抑制复合物 2(PRC2)的催化亚基,负责催化组蛋白 H3 第 27 位赖氨酸的三甲基化(H3K27me3),这一修饰通常与基因转录抑制相关。在 CRC 中,EZH2 的过表达可通过抑制一系列抑癌基因(包括与上皮-间质转化 EMT 和药物敏感性相关的基因)来促进肿瘤进展和耐药。转录因子 ZEB1 在驱动 CRC 转移和耐药时,就协调了组蛋白去乙酰化、组蛋白甲基化和 DNA 甲基化等多种表观遗传重编程事件,以沉默上皮表型相关基因。与此同时,HDACs 通过去除组蛋白尾部的乙酰基团,使染色质结构变得致密,从而抑制转录。HDACs 的异常活性与多种癌症的耐药性有关,这使得 HDAC 抑制剂(HDACi)成为逆转耐药的研究热点。研究表明,抑制 HDAC1/2 的催化活性可以促进 CRC 细胞分化并抑制肿瘤生长,其机制涉及关键组蛋白乙酰化标记(如 H3K27ac 和 H3K9ac)的全基因组重编程,这些修饰在 HDAC1/2 结合区域富集,并与分化基因的上调相关。此外,组蛋白修饰与 DNA 甲基化通路之间存在紧密的交互对话。例如,在胃癌中,DNA 甲基转移酶 1(DNMT1)、EZH2 和 HDAC1 被发现形成复合物,协同作用以更有效地沉默靶基因,这揭示了表观遗传调控网络的复杂性。因此,靶向这些组蛋白修饰酶,尤其是开发 HDAC 抑制剂,或联合使用 DNA 去甲基化药物(如 5-氮杂胞苷),已成为克服 CRC 化疗耐药极具前景的表观遗传治疗策略。5.2 非编码 RNA 的调控网络在结直肠癌(CRC)化疗耐药的形成中,非编码 RNA(ncRNA)构成了一个复杂而精密的调控网络,其中 microRNAs(miRNAs)扮演着关键角色。作为一类短链非编码 RNA,miRNAs 通过靶向信使 RNA(mRNA)的 3‘非翻译区(3’-UTR),在转录后水平调控基因表达,从而深刻影响肿瘤细胞的生物学行为。研究发现,多种 miRNAs 的表达失调与 CRC 的化疗耐药密切相关。例如,miR-21 和 miR-155 作为典型的致癌 miRNA,其过表达通过靶向 PTEN、PDCD4 等关键的抑癌基因,促进了 CRC 细胞的存活、增殖和耐药性。具体而言,miR-155 的过表达与多种化疗药物的耐药性相关,它通过调控广泛的 mRNA 转录本,影响细胞凋亡、细胞周期和 DNA 修复等通路,从而促进肿瘤细胞存活和上皮-间质转化(EMT)过程。相反,一些具有抑癌功能的 miRNAs,如 let-7 家族和 miR-34a,在耐药细胞中常常表达下调,导致其靶向的促癌基因(如 CCND1)失去调控。例如,miR-1247-3p 在 CRC 细胞中表达显著降低,而其靶基因细胞周期蛋白 D1(CCND1)的表达则相应升高;恢复 miR-1247-3p 的表达可以通过负调控 CCND1 来有效降低 CRC 细胞的化疗耐药性并促进细胞凋亡。此外,miR-100-5p 在奥沙利铂耐药的 CRC 细胞中表达上调,它通过靶向 C 末端结构域小磷酸酶样蛋白(CTDSPL),进而调控 pRB/E2F1 信号轴,增强了细胞周期介导的化疗耐药。这些研究共同揭示了 miRNAs 在 CRC 化疗耐药中的双向调控作用,它们既可作为驱动耐药的“帮凶”,也可作为逆转耐药的潜在“钥匙”,构成了耐药调控网络的核心环节。长链非编码 RNA(lncRNA)和环状 RNA(circRNA)作为另一大类重要的调控分子,通过更为复杂的机制参与 CRC 化疗耐药。它们能够作为竞争性内源 RNA(ceRNA),像“分子海绵”一样吸附 miRNA,解除 miRNA 对其靶 mRNA 的抑制作用,从而精细调控下游信号通路和耐药表型。例如,lncRNA BCAR4 在 CRC 组织和化疗耐药患者中表达升高,它通过吸附 miR-483-3p,促进其靶基因 RAB5C 的表达,从而增强了 CRC 细胞的干性、活力并抑制细胞凋亡,最终导致奥沙利铂耐药。另一个重要的 lncRNA H19 在缺氧或奥沙利铂处理的 CRC 细胞中表达增强,并通过充当 miR-675-3p 的 ceRNA 来调控上皮-间质转化(EMT),从而介导缺氧诱导的奥沙利铂耐药。此外,lncRNA PVT1-214 在奥沙利铂耐药的 CRC 细胞中异常高表达,并通过抑制 miR-128 的功能来促进耐药。环状 RNA 同样在此网络中发挥重要作用。研究显示,环状 RNA ciRS-122(hsa_circ_0005963)在奥沙利铂耐药的 CRC 细胞中高表达,它可以通过海绵吸附 miR-122,解除其对丙酮酸激酶 M2(PKM2)的抑制,从而促进糖酵解和 ATP 产生,最终诱导化疗耐药。这些 lncRNA 和 circRNA 通过构建复杂的 ceRNA 网络,不仅调控了包括 EMT、细胞干性、代谢重编程在内的多个关键生物学过程,还参与了耐药信号在细胞间的传递。例如,耐药细胞来源的外泌体可以将 ciRS-122 传递给敏感细胞,传播耐药表型。同样,外泌体中的 lncRNA,如 NEAT1、OIP5-AS1 和 MALAT1,也在奥沙利铂耐药中高表达,可能参与了细胞间通讯和耐药传播。综上所述,lncRNA 和 circRNA 通过 ceRNA 网络等机制,构成了一个多层次、动态的调控体系,精细地塑造了 CRC 的化疗耐药表型,为开发新的诊断标志物和逆转耐药的策略提供了极具潜力的靶点。6. 肿瘤干细胞与细胞可塑性6.1 干细胞特性维持与耐药结直肠癌干细胞(CSCs)是肿瘤内一小群具有自我更新、多向分化潜能和启动肿瘤形成能力的特殊细胞亚群,其存在是导致化疗耐药、肿瘤复发和转移的关键因素。这些细胞可通过特定的表面标志物进行鉴定和富集,其中 CD44、CD133 和 Lgr5 是研究最为广泛的标志物。研究表明,高表达 CD133 的结直肠癌细胞对 5-氟尿嘧啶(5-FU)和奥沙利铂等化疗药物表现出更强的抵抗性。此外,CD133 高表达与 CD24 低表达的组合被证明与结直肠癌患者更差的疾病特异性生存期相关,提示这种标志物组合可能具有预后价值。Lgr5 作为肠道干细胞的标志物,在结直肠癌干细胞中也高表达,其表达水平与肿瘤位置相关,在近端结直肠癌中表达更高。这些干细胞标志物的高表达不仅用于识别 CSCs,更与其内在的耐药特性紧密相连,例如,CD133+细胞中 ABC 转运蛋白家族成员如 ABCG2 的表达上调,能主动泵出化疗药物,降低细胞内药物浓度,从而介导多药耐药。结直肠癌干细胞的耐药性源于其一系列独特的生物学特性。首先,CSCs 具有强大的 DNA 损伤修复能力。研究发现,BMP 拮抗剂 CHRDL2 通过抑制 BMP 信号、增强 Wnt 信号,不仅能提升干细胞表型,还能激活 DNA 损伤应答通路,从而增强结直肠癌类器官和细胞在化疗和放疗下的存活能力。其次,CSCs 常处于细胞周期停滞或静息状态,这种休眠特性使其能逃避针对快速增殖细胞的周期特异性化疗药物的杀伤。例如,表达簇集蛋白(CLU)的“复苏干细胞”(RevSC)在稳态下处于静息状态,但在损伤(如化疗)后能够存活并再生其他干细胞,这与化疗耐药和患者不良生存相关。同样,在未经治疗的结直肠癌中存在的 ZEB2+静息/慢周期细胞群体,具有预定的间充质样化疗耐药表型,在化疗后成为优势群体,直接导致治疗无应答。最后,CSCs 高表达 ABC 转运蛋白(如 ABCG2、ABCB1/P-gp),能有效外排化疗药物。研究发现,核糖体生物合成因子 RPF2 可通过 CARM1-MYCN 轴上调 ABCB1 的表达,从而促进结直肠癌细胞的化疗耐药。这些特性共同构成了 CSCs 对化疗的天然防御屏障。维持结直肠癌干细胞干性和耐药的核心在于一系列关键信号通路的持续异常激活。Wnt/β-catenin 通路是最核心的调控通路之一。FOXD1 通过直接与β-catenin 结合,促进其核转位,激活下游 LGR5、SOX2 等干细胞基因的表达,从而增强细胞干性和对奥沙利铂的耐药性。同样,双皮质素样激酶 1(DCLK1)通过磷酸化并稳定 CCAR1,进而正调控β-catenin 信号通路,维持癌症干性,是导致 5-FU 耐药的重要机制。靶向此通路,例如使用β-catenin 抑制剂 XAV-939 或表观遗传药物丙戊酸(VPA)促进β-catenin 的泛素化降解,可以有效抑制干细胞特性并增敏化疗。Notch 信号通路是另一个关键通路。m6A 阅读器蛋白 YTHDF1 能结合 m6A 修饰的 NOTCH1 mRNA 并促进其翻译,从而激活 Notch 信号,驱动干细胞特性和对奥沙利铂及 5-FU 的耐药。Hedgehog 通路也与干性和耐药相关。研究发现,具核梭杆菌(Fusobacterium nucleatum)能促进结直肠癌干细胞特性并诱导化疗耐药,而二甲双胍可通过调控 MYC/miR-361-5p 级联反应抑制 Hedgehog 信号,从而逆转具核梭杆菌诱导的干性和耐药。此外,去泛素化酶 USP4 通过稳定β-catenin 和 Twist1 蛋白来促进干细胞特性,其小分子抑制剂 U4-I05 能降解这两种蛋白,抑制干性并增强对奥沙利铂和 5-FU 的敏感性。这些核心通路的交互作用构成了一个复杂的调控网络,共同维持着结直肠癌干细胞的生存、自我更新和耐药表型,使其成为治疗中难以根除的“种子”细胞。6.2 上皮-间质转化与耐药上皮-间质转化(EMT)是结直肠癌(CRC)获得化疗耐药和转移潜能的关键生物学过程。在 EMT 过程中,肿瘤细胞失去上皮表型(如 E-钙黏蛋白表达下调),获得间质表型(如波形蛋白、N-钙黏蛋白表达上调),从而获得迁移、侵袭和干细胞样特性。这种表型转化伴随着细胞周期减慢和凋亡抵抗,直接导致对化疗药物的敏感性降低。例如,在瑞戈非尼长期处理的 CRC 细胞中,观察到细胞呈现衰老样特征,并最终通过诱导 EMT 获得更侵袭性的表型和耐药性。同样,对奥沙利铂耐药的 CRC 细胞株也表现出明显的 EMT 特征,其增殖、迁移和侵袭能力增强,而凋亡能力减弱。研究表明,EMT 不仅与肿瘤的侵袭转移密切相关,更是 CRC 化疗失败的重要原因之一。部分 EMT(p-EMT)作为一种混合状态,同样与 CRC 细胞的迁移、转移和化疗耐药相关,尤其是在营养限制等压力条件下,p-EMT 特征基因签名与不良预后显著相关。因此,深入解析 EMT 过程及其调控网络,对于理解 CRC 耐药机制和开发新的治疗策略至关重要。EMT 转录因子(EMT-TFs),如 Snail、ZEB1 和 Twist,是驱动 CRC 细胞发生表型转化的核心调控者。这些转录因子不仅通过抑制上皮标志物(如 E-钙黏蛋白)和激活间质标志物(如波形蛋白)来诱导 EMT,还直接调控多种耐药相关基因的表达,从而形成耐药与转移的协同机制。例如,在 CRC 中,转录因子 SOX2 通过激活 ABCC2(一种多药耐药相关蛋白)的表达,并协同β-连环蛋白和自噬信号,共同维持化疗耐药、干细胞特性和 EMT 表型。同样,酪氨酸激酶受体 TYRO3 通过下调 ENO1 来抑制 EMT 过程,从而促进肿瘤发生和耐药。长链非编码 RNA NEF 则通过表观遗传学方式促进 DOK1 的表达,进而抑制 MEK/ERK 信号通路,从而抑制 EMT 和奥沙利铂耐药。此外,EMT-TFs 如 ZEB1 还能被其他分子反向调控,例如肌球蛋白重链 11(MYH11)通过抑制 ZEB1 的表达来抑制 EMT 和 CRC 进展。这些研究揭示了 EMT 转录因子处于一个复杂的调控网络中心,它们整合来自肿瘤微环境(如缺氧、细胞因子)和细胞内信号通路(如 Wnt/β-连环蛋白、STAT3)的信号,共同塑造 CRC 细胞的耐药和转移表型。靶向这些关键的 EMT 转录因子或其上游调控通路,有望成为逆转 CRC 化疗耐药的有效策略。7. 代谢重编程与化疗抵抗7.1 能量代谢途径的适应性改变结直肠癌(CRC)细胞在化疗压力下展现出显著的代谢重编程能力,这是其获得耐药性的关键机制之一。其中,有氧糖酵解(Warburg 效应)的增强是耐药细胞的典型特征,即使在氧气充足条件下,癌细胞也优先利用糖酵解途径产生能量和生物合成前体,这不仅满足了快速增殖的需求,还通过维持还原型烟酰胺腺嘌呤二核苷酸磷酸(NADPH)的产生来支持抗氧化防御系统,从而帮助细胞在药物诱导的氧化应激下存活。这种代谢转换并非孤立存在,耐药细胞还表现出高度的能量依赖性,能够在葡萄糖和谷氨酰胺代谢之间进行适应性转换,以应对营养匮乏的肿瘤微环境和治疗压力。例如,研究发现靶向这种双重能量依赖性的纳米系统(VSi-BP@HA)能够有效抑制 CRC 细胞的能量代谢,并逆转其化疗耐药性。此外,脂质代谢的重编程在耐药中也扮演着核心角色。耐药细胞通过增强脂肪酸氧化(FAO)和从头脂质合成(DNL)来维持能量和氧化还原平衡。特别是,脂质滴(LDs)的形成被证明是 CRC 药物耐受性持续细胞(DTPs)的关键生存策略,其中脂质合成关键酶 Lipin1(LPIN1)的表达上调,通过将游离脂肪酸隔离到脂质滴中,防止脂质过氧化和铁死亡,从而赋予细胞耐药性。同时,过氧化物酶体膜蛋白 PMP70 通过维持过氧化物酶体功能,保护耐药细胞免受过度脂质过氧化损伤,其缺失会破坏脂质稳态,导致细胞周期停滞和代谢功能受损。这些代谢途径的适应性改变共同构成了一个复杂的网络,使 CRC 细胞能够灵活应对化疗药物的攻击,维持其生存优势。7.2 氧化应激防御系统的增强化疗药物产生的活性氧(ROS)是杀伤肿瘤细胞的重要机制之一,然而耐药细胞通过上调其抗氧化防御系统,有效清除 ROS,从而削弱药物的效力。研究表明,结直肠癌(CRC)细胞能够通过多种途径增强其抗氧化能力以应对化疗诱导的氧化应激。例如,过氧化物酶体膜蛋白 PMP70 在奥沙利铂(LOHP)耐药的 CRC 细胞中高表达,其作为脂质过氧化的潜在保护者,通过维持过氧化物酶体稳定性和抵抗选择性自噬(pexophagy),帮助细胞清除过量的 ROS,从而赋予化疗耐药性。此外,核因子 E2 相关因子 2(NRF2)作为关键的抗氧化转录因子,其信号通路在 CRC 中被广泛研究。一篇系统性综述指出,氧化应激和炎症在 CRC 中紧密相连,其中 NRF2 是驱动结直肠癌发生的关键氧化还原敏感通路之一,其激活有助于促进基因组不稳定、免疫逃逸和化疗耐药。在临床样本分析中也发现,CRC 患者肿瘤周围脂肪组织中 NRF2/血红素加氧酶-1(HO-1)抗氧化级联的表达下调与疾病进展阶段相关,提示抗氧化防御系统的失调与不良预后有关。除了经典的 NRF2 通路,其他分子也参与调控氧化还原平衡。例如,亲环素 A(CypA)被鉴定为一种氧化还原敏感的效应物,在化疗耐药 CRC 样本中上调,它通过与过氧化物还原酶 2(PRDX2)相互作用,形成非经典的抗氧化防御装置,降低细胞内 ROS 水平,增加细胞在化疗药物攻击下的存活率。这些发现共同揭示了 CRC 细胞通过多层次、多靶点增强其抗氧化防御网络,是导致化疗耐药的关键机制。线粒体作为细胞的能量工厂和 ROS 的主要来源,其功能的适应性改变对于维持细胞在化疗压力下的存活至关重要。耐药细胞通过调整线粒体的融合/分裂动态平衡,有助于维持线粒体健康,避免凋亡信号的触发。研究表明,线粒体功能的改变与 CRC 的化疗耐药密切相关。例如,槐耳清膏处理 CRC 细胞可诱导线粒体损伤,表现为线粒体膜电位下降和线粒体活性氧(mtROS)升高,从而激活线粒体途径的细胞凋亡。相反,耐药细胞则可能通过增强线粒体质量控制来避免此类损伤。线粒体热休克蛋白 90 家族成员 TRAP1 在氧化应激下对结肠癌细胞凋亡具有调节作用。抑制 TRAP1 会加速氧化应激诱导的 DNA 损伤反应和细胞死亡,提示 TRAP1 在保护线粒体功能、帮助细胞适应氧化应激环境中扮演重要角色。此外,线粒体代谢的重编程也是适应性改变的一部分。有研究发现,耐药的 CRC 细胞会改变其代谢谱,例如 PMP70 缺失的细胞表现出中性脂质减少、脂滴减少和细胞周期停滞,这表明过氧化物酶体功能与线粒体代谢相互关联,共同维持细胞的稳态和耐药表型。线粒体动力学(融合与分裂)的平衡直接影响其功能、ROS 产生和细胞命运。虽然当前提供的参考文献中未直接阐述线粒体融合/分裂蛋白在 CRC 耐药中的作用,但上述研究均指向线粒体健康和功能完整性是抵抗化疗诱导凋亡的核心。因此,靶向线粒体质量控制通路或干扰其代谢适应性,可能成为克服 CRC 化疗耐药的新策略。8. 关键信号通路的异常激活8.1 PI3K/Akt/mTOR 通路PI3K/Akt/mTOR 信号通路是调控细胞生长、增殖、代谢和存活的核心通路,其在结直肠癌中的组成性激活是导致化疗耐药的关键机制之一。该通路的异常活化通过促进蛋白质合成、加速细胞周期进程并抑制细胞自噬和凋亡,从而成为多种化疗药物耐药的共同下游枢纽。例如,研究发现粘蛋白 3A(MUC3A)在结直肠癌中过表达,并通过激活 PI3K/Akt/mTOR 通路促进肿瘤细胞的增殖、侵袭以及对 5-氟尿嘧啶(5-FU)的化疗抵抗。使用该通路的抑制剂雷帕霉素处理,可以消除 MUC3A 敲除细胞与对照细胞在增殖、侵袭和化疗耐药性方面的差异。此外,该通路的激活与不良预后密切相关,其上游调控因子的异常,如 PTEN 的缺失或 PIK3CA 的突变,是导致通路持续活化的常见原因。这些遗传改变使得肿瘤细胞即使在化疗压力下也能维持生存信号,导致治疗失败。针对 PI3K/Akt/mTOR 通路的抑制剂已成为克服结直肠癌化疗耐药的研究热点,多项临床前及临床试验正在探索其与化疗联合应用的潜力。例如,在奥沙利铂耐药的结直肠癌细胞中,转录因子 KLF5 被鉴定为化疗耐药的潜在驱动因子,其通过激活 PI3K/AKT 通路赋予细胞耐药性。通过高通量药物筛选发现,PI3K/AKT 抑制剂 GDC-0941 能够与奥沙利铂协同作用,在临床前模型中增强疗效并克服耐药。另一项研究显示,岩藻多糖 FPS1M 作为 TLR4 的刺激剂,通过激活 TLR4 介导的 PI3K/AKT/mTOR 信号轴,促进巨噬细胞向 M1 表型分化,从而改善免疫抑制的肿瘤微环境,增强肿瘤对卡培他滨的化疗敏感性。这些研究表明,靶向该通路不仅能直接抑制肿瘤细胞存活,还能重塑肿瘤微环境,为逆转耐药提供了多角度的策略。目前,已有多种 PI3K、Akt 或 mTOR 的特异性抑制剂进入临床试验阶段,旨在与 5-FU、奥沙利铂等标准化疗方案联合,以期改善耐药患者的预后。8.2 MAPK/ERK 与 Wnt/β-catenin 通路KRAS、BRAF 等基因的突变是驱动结直肠癌(CRC)发生发展及化疗耐药的关键分子事件,尤其与右半结肠癌的临床特征和不良预后密切相关。这些突变导致 MAPK/ERK 信号通路持续激活,进而驱动肿瘤细胞异常增殖并抑制其凋亡,构成了对表皮生长因子受体(EGFR)靶向治疗及多种化疗药物产生耐药的重要基础。例如,携带 KRAS 突变的 CRC 细胞对西妥昔单抗等抗 EGFR 治疗天然耐药,而 BRAF V600E 突变则与更差的预后和对标准化疗及靶向治疗的抵抗相关。研究证实,在 KRAS 或 BRAF 突变的 CRC 细胞中,MAPK 通路抑制剂(如曲美替尼)虽能抑制下游 ERK 的磷酸化,但单药治疗效果有限,常伴随适应性耐药的出现。这种耐药机制部分源于 MEK/ERK 通路抑制后引发的代偿性反馈激活,例如抗凋亡蛋白 Bcl-xL 的上调,或通过 PI3K/AKT/mTOR 等其他通路的激活来维持细胞存活。因此,针对 MAPK 通路耐药,联合治疗策略显得尤为重要。例如,联合使用 MEK 抑制剂和 BCL-XL 抑制剂 ABT-263,在 KRAS 突变 CRC 的临床前模型中显示出协同的抗肿瘤效应。此外,转录因子 Yes 相关蛋白(YAP)的表达上调与 BRAF V600E 突变 CRC 细胞对 MAPK 通路抑制剂的原发性耐药有关,提示联合靶向 YAP 和 MAPK 通路可能是一种有前景的治疗策略。这些发现系统解析了 MAPK 通路异常在 CRC 耐药中的核心作用,并为克服此类耐药提供了潜在的联合用药方向。Wnt/β-catenin 信号通路的异常激活不仅是结直肠癌发生的早期驱动力,也在维持肿瘤干细胞特性及化疗耐药表型中扮演着核心角色。该通路的持续活化,通常由 APC、CTNNB1 等基因突变导致,能够上调多种多药耐药相关蛋白(如 ABC 转运蛋白)和干细胞相关基因的表达,从而赋予肿瘤细胞强大的生存和耐药能力。例如,在 CRC 细胞中,环状 RNA circ-ABCC1 通过外泌体从 CD133+肿瘤干细胞传递至其他细胞,激活 Wnt/β-catenin 通路,进而增强细胞的干性、转移能力和耐药性。类似地,circ-PRKDC 通过吸附 miR-375 上调 FOXM1,进而激活 Wnt/β-catenin 通路,导致对 5-氟尿嘧啶(5-FU)的耐药。成纤维细胞生长因子 9(FGF9)则通过下调 APC 表达来激活 Wnt 通路,介导了对顺铂的耐药。此外,Wnt 通路与 MAPK 通路之间存在复杂的交互作用(crosstalk),共同调控耐药表型。研究发现,在 KRAS/BRAF 突变的 CRC 中,Wnt 信号的激活与抗凋亡 BCL-2 家族蛋白(如 MCL1)的表达上调相关,遗传或药理学抑制 Wnt 信号可以增强 BCL-XL 抑制剂 ABT-263 的疗效。淋巴增强因子 1(LEF1)的长亚型高表达于 CRC 细胞,其下调能通过影响 Wnt 通路反应,增加细胞对奥沙利铂、5-FU 和伊立替康的敏感性。这些证据表明,靶向 Wnt/β-catenin 通路或其上下游调控分子,是逆转 CRC 干细胞介导的化疗耐药、改善治疗效果的潜在策略。例如,中药来源的活性成分如 3β-[(α-l-阿拉伯吡喃糖基)氧基]-urs-12,18(19)-二烯-28-酸β-d-葡萄糖吡喃酯(AGE)可通过介导 Wnt 信号通路诱导细胞周期阻滞,从而抑制 5-FU 敏感及耐药 CRC 细胞的增殖。9. 应对化疗耐药的综合策略与前沿进展9.1 基于机制的新型靶向药物联合针对特定耐药机制开发新型靶向药物,并与传统化疗联用,是克服结直肠癌(CRC)化疗耐药的重要策略。耐药机制复杂多样,涉及药物外排、DNA 损伤修复、细胞存活信号通路异常激活等多个层面。例如,ABC 转运蛋白家族的过表达是导致多药耐药(MDR)的经典机制之一,第三代抑制剂如 Tariquidar 旨在阻断这些外排泵的功能,从而恢复肿瘤细胞内化疗药物的有效浓度。然而,耐药性常由多种机制共同驱动,单一靶点抑制往往效果有限。因此,开发针对其他关键耐药节点的药物至关重要。聚腺苷二磷酸核糖聚合酶(PARP)抑制剂如奥拉帕利,通过抑制 DNA 损伤修复,对存在同源重组修复缺陷的肿瘤细胞具有合成致死效应。研究表明,在 CRC 中,某些亚群或经化疗诱导后可能表现出对 PARP 抑制剂的敏感性,将其与造成 DNA 损伤的化疗药物(如奥沙利铂)联用,有望克服由增强的 DNA 修复能力介导的耐药。同样,Akt 作为 PI3K/AKT/mTOR 信号通路的关键节点,其异常激活与细胞存活、增殖和化疗抵抗密切相关。Akt 抑制剂如 Ipatasertib,能够阻断该促生存信号,当与化疗联合时,可诱导更强的凋亡反应。研究证实,靶向抑制 Akt 通路能够逆转由 Claudin-1 与 EPHA2 相互作用所促进的癌症干细胞特性和化疗耐药。此外,针对肿瘤代谢重编程的干预也是重要方向。例如,抑制脂质从头合成(DNL)关键酶 Lipin1,能够破坏耐药持久细胞(DTPs)的脂质稳态,诱导铁死亡,从而克服其对常规化疗的抵抗。这些基于特定机制的新型靶向药物,通过与标准化疗方案(如 FOLFOX 或 FOLFIRI)进行合理组合,旨在从多个层面协同攻击耐药肿瘤细胞,为改善晚期 CRC 患者的治疗结局提供了新的希望。采用多靶点联合抑制策略,例如同时抑制 PI3K 和 MEK,旨在克服信号通路的代偿性激活和交叉对话,这是应对 CRC 靶向治疗耐药的前沿方向。肿瘤细胞具有强大的适应性,当一条关键信号通路被药物阻断时,常常会通过激活平行或下游的替代通路来维持生存和增殖,导致单药治疗失败。PI3K/AKT/mTOR 和 RAS/RAF/MEK/ERK 是 CRC 中两条频繁异常且相互串扰的核心信号通路。临床前和临床证据表明,单独抑制 MEK(MAPK/ERK 激酶)可能因解除对上游 RTK(如 EGFR)的反馈抑制而反弹性激活 PI3K/AKT 通路;反之,单独抑制 PI3K 也可能导致 MAPK 通路的代偿性激活。因此,同时靶向这两条通路被认为是一种有前景的策略,能够更全面地阻断致癌信号网络,防止代偿逃逸,从而更有效地抑制肿瘤生长并延缓耐药发生。例如,在携带 RAS 或 BRAF 突变的 CRC 中,MAPK 通路持续激活,而 PI3K 通路也常伴随异常,采用 MEK 抑制剂与 PI3K 或 AKT 抑制剂的联合方案已在临床前模型中显示出协同抗肿瘤活性。这种联合策略不仅适用于对 EGFR 单抗(如西妥昔单抗)耐药的 RAS 突变型 CRC,也可能为其他难治性亚群(如 BRAF V600E 突变型)提供治疗选择。除了上述通路,针对其他耐药相关靶点的联合也在探索中。例如,为了克服由癌症干细胞(CSCs)介导的耐药,联合靶向 Wnt/β-catenin 和 Notch 等干细胞相关通路的策略正在研究中。此外,将靶向药物与免疫检查点抑制剂联用,通过调节肿瘤微环境来增强抗肿瘤免疫,也是克服耐药的新兴组合模式。然而,多靶点联合也带来了挑战,如毒性叠加和最佳给药方案的确定。未来需要通过生物标志物指导的患者分层和合理的临床试验设计,来优化这些联合策略,实现疗效与安全性的平衡,最终为 CRC 患者提供更有效的个性化治疗方案。9.2 纳米技术与药物递送系统纳米技术在克服结直肠癌化疗耐药方面展现出巨大潜力,其核心在于设计智能纳米载体以实现药物的靶向递送和控释。这些载体,如脂质体、聚合物纳米粒等,能够利用肿瘤组织特有的增强渗透和滞留效应被动靶向至肿瘤部位,增加药物蓄积。为了进一步提升靶向特异性,纳米载体表面可修饰特定的靶向分子,例如叶酸、透明质酸或 RGD 肽,这些配体能够识别并结合在结直肠癌细胞上过表达的相应受体,从而实现主动靶向,将药物精准递送至肿瘤细胞内部,减少对正常组织的损伤。这种主动靶向策略已被证明能显著增强细胞对纳米药物的摄取,降低半数抑制浓度,并提高肿瘤选择性细胞毒性。更重要的是,智能纳米载体能够通过响应肿瘤微环境中的特定刺激(如 pH、酶或氧化还原状态)实现药物的可控释放,确保药物在病灶部位以有效浓度持续作用。此外,纳米载体还能有效包封并保护化疗药物,避免其被肿瘤细胞中过度表达的 ATP 结合盒转运蛋白外排,这是克服多药耐药的关键机制之一。通过整合被动靶向、主动识别和刺激响应释放,纳米递送系统为逆转结直肠癌化疗耐药提供了强有力的工具。开发共递送系统是纳米技术应对结直肠癌耐药的另一项前沿策略,该系统能够同时装载化疗药物和耐药逆转剂,在时空上实现协同作用。例如,纳米载体可以共递送传统化疗药物(如 5-氟尿嘧啶或奥沙利铂)与能够敲低耐药基因的小干扰 RNA,从而在抑制肿瘤生长的同时,从根源上削弱癌细胞的耐药能力。研究表明,将化疗药物与靶向特定耐药相关蛋白(如 Claudin-1、ZEB2 或癌症干细胞标记物)的 siRNA 共同装载,能够有效逆转由这些分子介导的化疗抵抗。除了基因治疗剂,纳米共递送系统还可以整合小分子耐药逆转剂。例如,将化疗药物与能够抑制脂滴形成或诱导铁死亡的药物(如他汀类药物)共同递送,可以靶向治疗耐受性持久细胞,通过破坏其脂质代谢稳态来克服耐药。这种协同策略不仅提高了治疗效果,还能减少因分别给药可能产生的毒副作用和药代动力学不匹配问题。此外,基于壳聚糖、脂质体或聚合物的纳米平台因其良好的生物相容性和可修饰性,成为构建此类多功能共递送系统的理想选择。通过将不同作用机制的 therapeutics 整合于单一纳米载体中,共递送系统为实现对耐药性结直肠癌的高效、精准联合治疗开辟了新途径。9.3 免疫治疗与化疗的协同化疗药物与免疫检查点抑制剂的联合应用是克服结直肠癌(CRC)免疫治疗耐药、提升疗效的重要策略。其协同效应的核心机制在于化疗能够重塑肿瘤免疫微环境(TME),为免疫治疗创造有利条件。一方面,以奥沙利铂、5-氟尿嘧啶(5-FU)等为基础的 FOLFOX 等化疗方案能够诱导免疫原性细胞死亡(ICD),释放损伤相关分子模式(DAMPs),从而增强肿瘤细胞的免疫原性,促进树突状细胞成熟和抗原呈递,为 T 细胞活化提供信号。另一方面,化疗药物能够选择性消耗或重编程免疫抑制性细胞。例如,雷帕霉素(Rapamycin)被证实可以重塑免疫抑制性 TME,有效消耗α-SMA+的癌症相关成纤维细胞(CAFs)并降解肿瘤组织中的胶原,从而增加 T 淋巴细胞向肿瘤组织的浸润,为抗 PD-1 治疗创造有利的免疫环境。此外,化疗还可能通过影响 PD-L1 等免疫检查点分子的表达来增强免疫治疗敏感性。研究表明,PD-L1 在肿瘤细胞中的低表达或缺失可能与化疗耐药相关,而通过 JNK 抑制剂等策略可以克服这种由 PD-L1 介导的耐药,从而恢复化疗药物的疗效。这些机制共同作用,使得化疗能够将“冷”肿瘤转化为“热”肿瘤,提高免疫检查点抑制剂的响应率。临床前研究也证实,将 FOLFOX 化疗方案与抗 PD-1 抗体或 PD-L1 siRNA 联合,通过协同诱导 ICD 和阻断 PD-1/PD-L1 通路,在结直肠癌小鼠模型中显示出显著的抗肿瘤效果。因此,深入理解化疗与免疫治疗之间的相互作用机制,对于设计有效的联合治疗方案至关重要。针对特定分子亚型的结直肠癌,尤其是错配修复缺陷/高度微卫星不稳定(dMMR/MSI-H)亚型,免疫治疗已成为一线标准治疗。然而,如何优化治疗策略以克服或延缓耐药是当前的研究重点。对于 dMMR/MSI-H 型 CRC,免疫检查点抑制剂(如抗 PD-1/PD-L1 单抗)单药或联合抗 CTLA-4 单抗已显示出卓越疗效。但部分患者仍会出现原发性或继发性耐药,因此探索免疫治疗与化疗的序贯或联合应用成为重要方向。联合策略旨在利用化疗的免疫调节作用,进一步增强免疫治疗的深度和持久性。例如,在 MSI-H 型 CRC 中,化疗可能通过增加肿瘤突变负荷或改变肿瘤抗原谱,与免疫治疗产生协同,从而延缓耐药发生。对于更为常见的错配修复完整/微卫星稳定(pMMR/MSS)型 CRC,其本身对单药免疫治疗反应不佳,联合化疗则成为激活其免疫应答的关键手段。研究表明,化疗药物如伊立替康(Irinotecan)的治疗效果可能受到其下游信号通路(如 NF-κB、PD-L1)激活的限制,而这些通路同时介导了免疫抑制,因此靶向这些通路(如联合 PD-L1 抑制剂)可能成为克服化疗和免疫治疗双重耐药的有效策略。此外,通过调节肠道微生物群来增强免疫治疗反应也是一个新兴领域,例如尿石素 B(Urolithin B)等微生物代谢产物能够重塑肠道菌群和肿瘤免疫微环境,降低 PD-L1 表达,当与一线化疗药物或抗 PD-1 抗体联用时,可改善治疗效果。这些研究为针对不同亚型 CRC 制定个体化的化疗-免疫联合或序贯方案提供了理论依据和新的思路。9.4 耐药性动态监测与个体化治疗利用液体活检技术,特别是循环肿瘤 DNA(ctDNA)和循环肿瘤细胞(CTC),为结直肠癌化疗耐药的实时动态监测提供了革命性的工具。传统的组织活检具有侵入性、无法重复取样且难以反映肿瘤时空异质性的局限,而液体活检通过检测外周血中肿瘤来源的生物标志物,能够无创、高频次地追踪治疗过程中的基因变异动态。例如,在结直肠癌患者中,通过 ctDNA 测序可以早期发现与奥沙利铂或 5-氟尿嘧啶耐药相关的基因突变,如 KRAS、BRAF 或 PIK3CA 的获得性突变,从而在影像学进展之前预警耐药克隆的出现。此外,CTC 不仅可用于计数评估预后,其分子特征分析还能揭示上皮-间质转化(EMT)等与侵袭和耐药相关的表型变化。基于这些动态信息,临床医生能够及时调整治疗方案,例如在检测到特定耐药突变后,从标准化疗方案切换为针对该通路的靶向药物或新型联合疗法,实现治疗策略的主动干预,避免无效治疗并改善患者预后。这种“实时反馈-动态调整”的模式,标志着结直肠癌治疗从静态、经验性向动态、精准化的重大转变。基于患者肿瘤组织的多组学分析是实现“一人一策”个体化治疗方案的基石。通过对肿瘤样本进行基因组、转录组、蛋白组等多层次的整合分析,可以全面解析驱动肿瘤发生、进展及耐药的核心分子网络。例如,基因组测序能够鉴定出如 RAS、RAF、错配修复基因等关键驱动突变和耐药突变;转录组分析可以揭示与癌症干细胞特性、EMT 或特定信号通路(如 Wnt/β-catenin、PI3K/AKT)异常激活相关的基因表达谱,这些特征常与化疗耐药密切相关;而蛋白组学则能反映信号通路的实际活化状态及药物靶点的蛋白表达水平。利用这些海量的多组学数据,可以构建复杂的预测模型,用于评估患者对特定化疗或靶向药物(如奥沙利铂、5-氟尿嘧啶、西妥昔单抗等)的初始敏感性及耐药风险。例如,有研究通过分析患者来源的类器官(PDOs)模型,结合转录组学,成功揭示了环状 RNA hsa_circ_0020095 在介导 5-FU 和奥沙利铂耐药中的作用,并验证了其作为治疗靶点的潜力。这种深度分子剖析使得治疗方案能够超越传统的病理分型,根据患者独特的肿瘤生物学特征进行定制,例如为具有特定基因突变或通路激活的患者匹配相应的靶向药物或通路抑制剂,或针对高表达多药耐药蛋白(如 P-gp)的患者设计包含逆转剂的联合方案,从而最大化疗效并克服耐药。结论结直肠癌化疗耐药的形成与演变,是一个由肿瘤细胞内在特性与肿瘤微环境外部因素共同驱动、多机制交织互作的动态过程。从[专家视角]审视,这一领域的认知已从早期聚焦于单一基因突变或通路异常,深化为对复杂调控网络的系统性理解。例如,药物外排泵的激活、DNA 损伤修复能力的增强、凋亡逃逸以及表观遗传重塑等内在机制,与肿瘤相关成纤维细胞、免疫细胞等构成的微环境压力之间,存在着广泛的“交叉对话”和代偿性激活。这意味着,任何针对单一环节的干预,都可能迅速被网络中的其他节点所补偿,从而导致治疗失败。因此,未来的研究必须超越“线性思维”,采用系统生物学的方法,解析这些交互网络的核心枢纽与关键节点。面对这一挑战,平衡不同研究的观点意味着需承认耐药机制的异质性——不同患者、甚至同一患者不同病灶或不同治疗阶段,主导的耐药机制可能截然不同。这要求我们的应对策略从“一刀切”转向“组合拳”和“动态调整”。联合疗法,如将传统化疗与针对特定信号通路(如 EGFR、VEGF)的靶向药物、旨在解除免疫抑制的免疫检查点抑制剂、以及能物理性破坏肿瘤结构的消融治疗等相结合,已成为克服或延缓耐药的重要方向。而纳米递药系统等前沿技术,为实现多种作用机制药物的协同递送、提高靶向性和降低全身毒性提供了强大工具。然而,将实验室发现转化为临床效益,始终是最大的瓶颈。加强转化研究至关重要,需要充分利用患者来源类器官、人源肿瘤异种移植模型等能够较好模拟肿瘤体内特征和异质性的平台,来高通量筛选和验证潜在的耐药逆转策略。未来的突破性进展可能依赖于以下几方面的协同推进:一是开发能够同时干预多个关键耐药节点的新型小分子或生物制剂,并确保其安全性;二是建立基于循环肿瘤 DNA、循环肿瘤细胞等液体活检技术的实时、动态耐药监测体系,实现治疗过程中的“早发现、早干预”;三是构建整合临床病理特征、医学影像组学与多组学分子信息的智能诊疗系统,通过人工智能算法预测耐药风险、甄别主导机制并推荐最优的个体化治疗组合。唯有通过这种多维度、一体化的精准医疗模式,方有望最终破解结直肠癌的化疗耐药困局,显著延长患者的生存期并提高生活质量。参考文献[1] Sabeti Aghabozorgi A, Moradi Sarabi M, Jafarzadeh-Esfehani R, et al. Molecular determinants of response to 5-fluorouracil-based chemotherapy in colorectal cancer: The undisputable role of micro-ribonucleic acids. World J Gastrointest Oncol. 2020;12(9):942-956. doi:10.4251/wjgo.v12.i9.942 [2] Anselmino LE, Malizia F, Avila A, et al. Overcoming Therapy Resistance in Colorectal Cancer: Targeting the Rac1 Signaling Pathway as a Potential Therapeutic Approach. Cells. 2024;13(21). Published 2024 Oct 26. doi:10.3390/cells13211776 [3] Sreekumar R, Al-Saihati H, Emaduddin M, et al. The ZEB2-dependent EMT transcriptional programme drives therapy resistance by activating nucleotide excision repair genes ERCC1 and ERCC4 in colorectal cancer. Mol Oncol. 2021;15(8):2065-2083. doi:10.1002/1878-0261.12965 [4] Kumar B, Ahmad R, Sharma S, et al. PIK3C3 Inhibition Promotes Sensitivity to Colon Cancer Therapy by Inhibiting Cancer Stem Cells. Cancers (Basel). 2021;13(9). Published 2021 Apr 30. doi:10.3390/cancers13092168 [5] Hjazi A, Saleh RO, Al-Khafaji ZA, et al. Exosomal Dialogue: The Emerging Role of Cancer-Associated Fibroblast-Derived Exosomes in Colorectal Cancer Progression and Therapy Resistance. J Biochem Mol Toxicol. 2026;40(4):e70780. doi:10.1002/jbt.70780 [6] Huang P, Guo K, Tu J, et al. Single-Cell RNA Sequencing Reveals LEF1 as a Prognostic Biomarker for Poor Outcomes in Oxaliplatin-Resistant Colorectal Cancer. Hum Mutat. 2025:6705599. Published 2025 None. doi:10.1155/humu/6705599 [7] Hao W, Wang Z, Ma H. Characterization of stem cell landscape and identification of stemness-related gene ENY2 in colorectal cancer by intergrated transcriptomic analysis. Glob Med Genet. 2025;12(4):100067. Published 2025 Dec. doi:10.1016/j.gmg.2025.100067 [8] Jiang JK, Lin CH, Lin CC, et al. Interleukin-17 receptor A drives cancer stem-like properties in colorectal cancer through STAT3 activation. J Cancer. 2026;17(1):74-85. Published 2026 None. doi:10.7150/jca.121654 [9] Hlavca S, Chan WH, Engel RM, Abud HE. Clusterin: a marker and mediator of chemoresistance in colorectal cancer. Cancer Metastasis Rev. 2024;43(1):379-391. doi:10.1007/s10555-024-10173-y [10] Francescangeli F, Contavalli P, De Angelis ML, et al. A pre-existing population of ZEB2+ quiescent cells with stemness and mesenchymal features dictate chemoresistance in colorectal cancer. J Exp Clin Cancer Res. 2020;39(1):2. Published 2020 Jan 8. doi:10.1186/s13046-019-1505-4 [11] Janakiraman H, Gao Z, Zhu Y, et al. Targeting SNAI1-Mediated Colorectal Cancer Chemoresistance and Stemness by Sphingosine Kinase 2 Inhibition. World J Oncol. 2024;15(5):744-757. doi:10.14740/wjon1890 [12] Kiu KT, Chu CY, Cheng YC, et al. SIAH2-WNK1 Signaling Drives Glycolytic Metabolism and Therapeutic Resistance in Colorectal Cancer. Int J Mol Sci. 2026;27(2). Published 2026 Jan 21. doi:10.3390/ijms27021065 [13] Jin Y, Hu C, Pang D. FOXA2/ALDOB axis modulation of fatty acid beta-oxidation influences irinotecan resistance in colorectal cancer. Biochim Biophys Acta Mol Cell Res. 2026;1873(3):120108. doi:10.1016/j.bbamcr.2026.120108 [14] Yin J, Shao Y, Huang F, et al. Peroxisomal membrane protein PMP70 confers drug resistance in colorectal cancer. Cell Death Dis. 2025;16(1):293. Published 2025 Apr 14. doi:10.1038/s41419-025-07572-6 [15] Chern Y, Zhang P, Ju H, Tai IT. Heat shock protein 47 promotes tumor survival and therapy resistance by modulating AKT signaling via PHLPP1 in colorectal cancer. Cancer Biol Med. 2020;17(2):343-356. doi:10.20892/j.issn.2095-3941.2019.0261 [16] Park SY, Kim JY, Jang GB, et al. Aberrant activation of the CD45-Wnt signaling axis promotes stemness and therapy resistance in colorectal cancer cells. Theranostics. 2021;11(18):8755-8770. Published 2021 None. doi:10.7150/thno.63446 [17] Pan D, Hu J, Li G, et al. BRD4 regulates PAI-1 expression in tumor-associated macrophages to drive chemoresistance in colorectal cancer. Oncogene. 2025;44(31):2778-2791. doi:10.1038/s41388-025-03453-6 [18] Pandey K, Umar S. Microbiome in drug resistance to colon cancer. Curr Opin Physiol. 23. doi:10.1016/j.cophys.2021.100472 [19] Deng X, Li D, Ke X, et al. Mir-488 alleviates chemoresistance and glycolysis of colorectal cancer by targeting PFKFB3. J Clin Lab Anal. 2021;35(1):e23578. doi:10.1002/jcla.23578 [20] Xin Z, Hu C, Zhang C, et al. LncRNA-HMG incites colorectal cancer cells to chemoresistance via repressing p53-mediated ferroptosis. Redox Biol. 77:103362. doi:10.1016/j.redox.2024.103362 [21] Hussen BM, Abdullah SR, Mohammed AA, et al. Advanced strategies of targeting circular RNAs as therapeutic approaches in colorectal cancer drug resistance. Pathol Res Pract. 260:155402. doi:10.1016/j.prp.2024.155402 [22] Luo M, Yang X, Chen HN, Nice EC, Huang C. Drug resistance in colorectal cancer: An epigenetic overview. Biochim Biophys Acta Rev Cancer. 2021;1876(2):188623. doi:10.1016/j.bbcan.2021.188623 [23] Fatima I, Ahmad R, Barman S, et al. Albendazole inhibits colon cancer progression and therapy resistance by targeting ubiquitin ligase RNF20. Br J Cancer. 2024;130(6):1046-1058. doi:10.1038/s41416-023-02570-x [24] Shah ET, Molloy C, Gough M, et al. Inhibition of Aurora B kinase (AURKB) enhances the effectiveness of 5-fluorouracil chemotherapy against colorectal cancer cells. Br J Cancer. 2024;130(7):1196-1205. doi:10.1038/s41416-024-02584-z [25] Laoukili J, Constantinides A, Wassenaar ECE, et al. Peritoneal metastases from colorectal cancer belong to Consensus Molecular Subtype 4 and are sensitised to oxaliplatin by inhibiting reducing capacity. Br J Cancer. 2022;126(12):1824-1833. doi:10.1038/s41416-022-01742-5 [26] Primeaux M, Liu X, Gowrikumar S, et al. Claudin-1 interacts with EPHA2 to promote cancer stemness and chemoresistance in colorectal cancer. Cancer Lett. 579:216479. doi:10.1016/j.canlet.2023.216479 [27] Pan Q, Fan X, Xie L, et al. Nano-enabled colorectal cancer therapy. J Control Release. 362:548-564. doi:10.1016/j.jconrel.2023.09.014 [28] Wang X, Xu Z, Wang J, et al. A Mitochondria-Targeted Biomimetic Nanomedicine Capable of Reversing Drug Resistance in Colorectal Cancer Through Mitochondrial Dysfunction. Adv Sci (Weinh). 2025;12(13):e2410630. doi:10.1002/advs.202410630 [29] Pricope DL, Grigoraș A, Dimofte GM, Amalinei C. The Multifaceted Role of Annexin A1 in Colorectal Cancer: From Molecular Mechanisms to Predictive and Prognostic Implications. Med Sci (Basel). 2025;13(4). Published 2025 Nov 10. doi:10.3390/medsci13040263 [30] Nagata K, Shinto E, Shiraishi T, et al. Mesothelin Expression is Correlated with Chemoresistance in Stage IV Colorectal Cancer. Ann Surg Oncol. 2021;28(13):8579-8586. doi:10.1245/s10434-021-10507-y [31] Xiao A, Fakih M. Response to anti-EGFR therapy in chemo-refractory right-sided RAS wild-type metastatic colorectal cancer: a case report and literature review. J Gastrointest Oncol. 2025;16(1):292-300. doi:10.21037/jgo-24-458 [32] Dean M, Moitra K, Allikmets R. The human ATP-binding cassette (ABC) transporter superfamily. Hum Mutat. 2022;43(9):1162-1182. doi:10.1002/humu.24418 [33] Nikolova B, Semkova S, Tsoneva I, et al. Redox-related Molecular Mechanism of Sensitizing Colon Cancer Cells to Camptothecin Analog SN38. Anticancer Res. 2020;40(9):5159-5170. doi:10.21873/anticanres.14519 [34] Yang Z, Zhao F, Gu X, et al. Binding of RNA m6A by IGF2BP3 triggers chemoresistance of HCT8 cells via upregulation of ABCB1. Am J Cancer Res. 2021;11(4):1428-1445. Published 2021 None. [35] Po A, Citarella A, Catanzaro G, et al. Hedgehog-GLI signalling promotes chemoresistance through the regulation of ABC transporters in colorectal cancer cells. Sci Rep. 2020;10(1):13988. Published 2020 Aug 19. doi:10.1038/s41598-020-70871-9 [36] Gao HL, Gupta P, Cui Q, et al. Sapitinib Reverses Anticancer Drug Resistance in Colon Cancer Cells Overexpressing the ABCB1 Transporter. Front Oncol. 10:574861. Published 2020 None. doi:10.3389/fonc.2020.574861 [37] Liu C, Xing W, Yu H, Zhang W, Si T. ABCB1 and ABCG2 restricts the efficacy of gedatolisib (PF-05212384), a PI3K inhibitor in colorectal cancer cells. Cancer Cell Int. 2021;21(1):108. Published 2021 Feb 16. doi:10.1186/s12935-021-01800-7 [38] 乐音子, 顾云慧, 王雅慧, 等. 人结肠癌 5-氟尿嘧啶耐药细胞模型的构建及筛选[J]. 中国老年学杂志, 2024,44(18):4559-4564. [39] Gao Q, Li XX, Xu YM, et al. IRE1α-targeting downregulates ABC transporters and overcomes drug resistance of colon cancer cells. Cancer Lett. 476:67-74. doi:10.1016/j.canlet.2020.02.007 [40] El-Daly SM, Abo-Elfadl MT, Hussein J, Abo-Zeid MAM. Enhancement of the antitumor effect of 5-fluorouracil with modulation in drug transporters expression using PI3K inhibitors in colorectal cancer cells. Life Sci. 315:121320. doi:10.1016/j.lfs.2022.121320 [41] Vaghari-Tabari M, Majidinia M, Moein S, et al. MicroRNAs and colorectal cancer chemoresistance: New solution for old problem. Life Sci. 259:118255. doi:10.1016/j.lfs.2020.118255 [42] Shi Y, Wan T, Liu S, et al. Dynamic Arginine Methylation of YBX1 Relay Controls Its Phase Separation and Chemoradiotherapy Resistance in Rectal Cancer. Adv Sci (Weinh). 2025;12(46):e02786. doi:10.1002/advs.202502786 [43] Nikodijević DD, Jovankić JV, Cvetković DM, Anđelković MZ, Nikezić AG, Milutinović MG. L-amino acid oxidase from snake venom: Biotransformation and induction of apoptosis in human colon cancer cells. Eur J Pharmacol. 910:174466. doi:10.1016/j.ejphar.2021.174466 [44] Radenković N, Milenković D, Nikodijević D, Jovanović Stojanov S, Podolski Renić A, Milutinović M. Isorhamnetin Modulates Drug-Resistance-Related Biomarkers in Colon Cancer Cells. Int J Mol Sci. 2025;26(13). Published 2025 Jun 27. doi:10.3390/ijms26136208 [45] Ren SN, Zhang ZY, Guo RJ, et al. Application of nanotechnology in reversing therapeutic resistance and controlling metastasis of colorectal cancer. World J Gastroenterol. 2023;29(13):1911-1941. doi:10.3748/wjg.v29.i13.1911 [46] Alketbi L, Al-Ali A, Talaat IM, Hamid Q, Bajbouj K. The Role of ATP-Binding Cassette Subfamily A in Colorectal Cancer Progression and Resistance. Int J Mol Sci. 2023;24(2). Published 2023 Jan 10. doi:10.3390/ijms24021344 [47] Vlahović I, Rajc J, Švagelj I, Šolić K, Švagelj D. Potential predictors for CDX2 expression loss and mismatch repair deficiency in colorectal cancer. Pathol Oncol Res. 29:1610908. Published 2023 None. doi:10.3389/pore.2023.1610908 [48] Rico SD, Höflmayer D, Büscheck F, et al. Elevated MUC5AC expression is associated with mismatch repair deficiency and proximal tumor location but not with cancer progression in colon cancer. Med Mol Morphol. 2021;54(2):156-165. doi:10.1007/s00795-020-00274-2 [49] 常东, 王帆, 韩冰, 等. DNA 损伤修复与大肠癌关系的研究进展[J]. 癌变·畸变·突变, 2008,20(5):415-417. [50] Cercek A, Dos Santos Fernandes G, Roxburgh CS, et al. Mismatch Repair-Deficient Rectal Cancer and Resistance to Neoadjuvant Chemotherapy. Clin Cancer Res. 2020;26(13):3271-3279. doi:10.1158/1078-0432.CCR-19-3728 [51] Farchoukh LF, Celebrezze J, Medich D, et al. DNA Mismatch Repair-deficient Rectal Cancer Is Frequently Associated With Lynch Syndrome and With Poor Response to Neoadjuvant Therapy. Am J Surg Pathol. 2022;46(9):1260-1268. doi:10.1097/PAS.0000000000001918 [52] 赵欣楠, 魏敏杰, 赵琳. DNA 损伤修复调控因子与肿瘤耐药的研究进展[J]. 现代肿瘤医学, 2013,21(11):2608-2612. [53] Grigorie TR, Potlog G, Alexandrescu ST. Lynch Syndrome-Impact of the Type of Deficient Mismatch Repair Gene Mutation on Diagnosis, Clinical Presentation, Surveillance and Therapeutic Approaches. Medicina (Kaunas). 2025;61(1). Published 2025 Jan 14. doi:10.3390/medicina61010120 [54] Durhuus JA, Galanakis M, Maltesen T, et al. A registry-based study on universal screening for defective mismatch repair in colorectal cancer in Denmark highlights disparities in screening uptake and counselling referrals. Transl Oncol. 46:102013. doi:10.1016/j.tranon.2024.102013 [55] 夏莹, 周东波, 胡成平. DNA 修复与顺铂耐药[J]. 国际病理科学与临床杂志, 2006,26(4):336-339. [56] 刘博雅, 杨鑫, 任梦梦, 等. DNA 损伤修复机制——解读 2015 年诺贝尔化学奖[J]. 中国生物化学与分子生物学报, 2015,31(12): [57] 唐春兰. DNA 损伤修复与肺癌顺铂耐药机制的研究进展[J]. 中国肺癌杂志, 2011,14(12):960-964. [58] 王秋明, 黄婉兰, 林英城. 非小细胞肺癌分子预测标志物[J]. 国际肿瘤学杂志, 2011,38(4): [59] 邝仕成. XPD 基因单核苷酸多态与肿瘤遗传易感性的研究进展[J]. 医学信息, 2015(32):399-399. [60] 朱守民, 夏昭林. DNA 损伤修复基本方式的研究进展[J]. 国外医学(分子生物学分册), 2003,25(5):270-272. [61] 董怡萍, 张丹, 韩苏夏. DNA 损伤修复机制的研究进展[J]. 中华放射肿瘤学杂志, 2017,26(9):1103-1108. [62] Li J, Steffen P, Tse BCY, et al. Deep Sequencing of Early T Stage Colorectal Cancers Reveals Disruption of Homologous Recombination Repair in Microsatellite Stable Tumours with High Mutational Burdens. Cancers (Basel). 2022;14(12). Published 2022 Jun 14. doi:10.3390/cancers14122933 [63] Lin Y, Liao X, Zhang Y, et al. Homologous Recombination Pathway Alternation Predicts Prognosis of Colorectal Cancer With Chemotherapy. Front Pharmacol. 13:920939. Published 2022 None. doi:10.3389/fphar.2022.920939 [64] Xu W, Shen M, Ma J, et al. The cytoskeletal protein smoothelin maintains homologous recombination repair by stabilizing RAD51 in an HUWE1-dependent manner in colorectal cancer. Acta Pharm Sin B. 2025;15(12):6444-6460. doi:10.1016/j.apsb.2025.09.042 [65] Tu JL, Wu BH, Wu HB, et al. Design, synthesis and evaluation of N3-substituted quinazolinone derivatives as potential Bloom’s Syndrome protein (BLM) helicase inhibitor for sensitization treatment of colorectal cancer. Eur J Med Chem. 246:114944. doi:10.1016/j.ejmech.2022.114944 [66] Sun X, Hou W, Liu X, Chai J, Guo H, Yu J. Targeting REV7 effectively reverses 5-FU and oxaliplatin resistance in colorectal cancer. Cancer Cell Int. 2020;20(1):580. Published 2020 Dec 3. doi:10.1186/s12935-020-01668-z [67] Weng X, Ma T, Chen Q, et al. Decreased expression of H19/miR-675 ameliorates hypoxia-induced oxaliplatin resistance in colorectal cancer. Heliyon. 2024;10(5):e27027. Published 2024 Mar 15. doi:10.1016/j.heliyon.2024.e27027 [68] Palassin P, Lapierre M, Bonnet S, et al. RIP140 regulates POLK gene expression and the response to alkylating drugs in colon cancer cells. Cancer Drug Resist. 2022;5(2):401-414. Published 2022 None. doi:10.20517/cdr.2021.133 [69] Chen J, Xu J, Yang J, et al. α‑hederin overcomes hypoxia‑mediated drug resistance in colorectal cancer by inhibiting the AKT/Bcl2 pathway. Int J Oncol. 2023;62(3). doi:10.3892/ijo.2023.5481 [70] Tu R, Kang W, Kang Y, et al. C-MYC-USP49-BAG2 axis promotes proliferation and chemoresistance of colorectal cancer cells in vitro. Biochem Biophys Res Commun. 607:117-123. doi:10.1016/j.bbrc.2022.03.138 [71] Pothuraju R, Rachagani S, Krishn SR, et al. Molecular implications of MUC5AC-CD44 axis in colorectal cancer progression and chemoresistance. Mol Cancer. 2020;19(1):37. Published 2020 Feb 25. doi:10.1186/s12943-020-01156-y [72] Li M, Sun S, Bian Z, et al. SNHG15 promotes chemoresistance and glycolysis in colorectal cancer. Pathol Res Pract. 246:154480. doi:10.1016/j.prp.2023.154480 [73] Sun K, Chen L, Li Y, et al. METTL14-dependent maturation of pri-miR-17 regulates mitochondrial homeostasis and induces chemoresistance in colorectal cancer. Cell Death Dis. 2023;14(2):148. Published 2023 Feb 21. doi:10.1038/s41419-023-05670-x [74] Wang H, Wang X, Zhang H, et al. The HSF1/miR-135b-5p axis induces protective autophagy to promote oxaliplatin resistance through the MUL1/ULK1 pathway in colorectal cancer. Oncogene. 2021;40(28):4695-4708. doi:10.1038/s41388-021-01898-z [75] 王松, 李飞峰, 刘树林, 等. 线粒体自噬对结直肠癌的影响[J]. 国际遗传学杂志, 2017,40(2):98-103. [76] 张容圣, 李庚, 张小超, 等. 肿瘤相关成纤维细胞上调自噬促进结直肠癌耐药[J]. 中华实验外科杂志, 2018,35(10):1878-1880. [77] Siri M, Behrouj H, Dastghaib S, et al. Casein Kinase-1-Alpha Inhibitor (D4476) Sensitizes Microsatellite Instable Colorectal Cancer Cells to 5-Fluorouracil via Authophagy Flux Inhibition. Arch Immunol Ther Exp (Warsz). 2021;69(1):26. Published 2021 Sep 18. doi:10.1007/s00005-021-00629-2 [78] 戴维, 赵燕, 赵成进. 藤黄酸诱导结直肠癌 Caco2 细胞自噬和凋亡[J]. 山西医科大学学报, 2019,50(11):1510-1514. [79] 彭鹏, 杨书胜, 向雨晨, 等. 重楼皂苷 A 直接靶向活化 AMPK 诱导结直肠癌细胞自噬[J]. 江苏大学学报(医学版), 2021,31(4):290-295. [80] 杨成香, 李艳. 自噬在结直肠癌中的双重作用和研究现状[J]. 肿瘤, 2024,44(3):252-261. [81] 郭桂芳, 夏良平. 自噬及其与结直肠癌的发生和治疗[J]. 国际肿瘤学杂志, 2010,37(10): [82] Zheng L, Lu J, Kong DL. Expression of cyclin-dependent kinase 9 is positively correlated with the autophagy level in colon cancer. World J Gastrointest Oncol. 2024;16(2):314-330. doi:10.4251/wjgo.v16.i2.314 [83] Wang N, Zhang L, Leng XX, et al. Fusobacterium nucleatum induces chemoresistance in colorectal cancer by inhibiting pyroptosis via the Hippo pathway. Gut Microbes. 2024 Jan-Dec;16(1):2333790. doi:10.1080/19490976.2024.2333790 [84] 谢春岐, 牛犇, 费禹翔, 等. 癌症相关成纤维细胞在结直肠癌中的研究进展[J]. 药学进展, 2025,49(9):733-739. [85] Yang T, Zhiheng H, Zhanhuai W, et al. Increased RAB31 Expression in Cancer-Associated Fibroblasts Promotes Colon Cancer Progression Through HGF-MET Signaling. Front Oncol. 10:1747. Published 2020 None. doi:10.3389/fonc.2020.01747 [86] Wang J, Lu Y, Wang Z, Wei P. Cancer-Associated Fibroblasts Enhance Oxaliplatin Resistance in Colorectal Cancer Cells via Paracrine IL-6: An In Vitro Study. Iran J Pharm Res. 2025 Jan-Dec;24(1):e160886. Published 2025 Jan-Dec None. doi:10.5812/ijpr-160886 [87] Jiang Y, Qiu Q, Jing X, et al. Cancer-associated fibroblast-derived exosome miR-181b-3p promotes the occurrence and development of colorectal cancer by regulating SNX2 expression. Biochem Biophys Res Commun. 641:177-185. doi:10.1016/j.bbrc.2022.12.026 [88] Galindo-Pumariño C, Collado M, Castillo ME, et al. SNAI1-expressing fibroblasts and derived-extracellular matrix as mediators of drug resistance in colorectal cancer patients. Toxicol Appl Pharmacol. 450:116171. doi:10.1016/j.taap.2022.116171 [89] Giguelay A, Turtoi E, Khelaf L, et al. The landscape of cancer-associated fibroblasts in colorectal cancer liver metastases. Theranostics. 2022;12(17):7624-7639. Published 2022 None. doi:10.7150/thno.72853 [90] Hirashima T, Karasawa H, Aizawa T, et al. Wnt5a in cancer-associated fibroblasts promotes colorectal cancer progression. Biochem Biophys Res Commun. 568:37-42. doi:10.1016/j.bbrc.2021.06.062 [91] Yang L, Akbarabadi P, Babashah S. Cancer-associated fibroblasts-derived exosomes in colorectal cancer progression: Mechanism and therapeutic opportunities. Biochim Biophys Acta Rev Cancer. 2026;1881(2):189545. doi:10.1016/j.bbcan.2026.189545 [92] Gong J, Lin Y, Zhang H, et al. Reprogramming of lipid metabolism in cancer-associated fibroblasts potentiates migration of colorectal cancer cells. Cell Death Dis. 2020;11(4):267. Published 2020 Apr 23. doi:10.1038/s41419-020-2434-z [93] Chen Y, Liang Z, Lai M. Targeting the devil: Strategies against cancer-associated fibroblasts in colorectal cancer. Transl Res. 270:81-93. doi:10.1016/j.trsl.2024.04.003 [94] Karta J, Meyers M, Rodriguez F, et al. Fusobacterium nucleatum interacts with cancer-associated fibroblasts to promote colorectal cancer. EMBO J. 2025;44(19):5375-5393. doi:10.1038/s44318-025-00542-w [95] Agorku DJ, Bosio A, Alves F, Ströbel P, Hardt O. Colorectal cancer-associated fibroblasts inhibit effector T cells via NECTIN2 signaling. Cancer Lett. 595:216985. doi:10.1016/j.canlet.2024.216985 [96] 那迪娜·帕尔哈提, 黄陈. 肿瘤相关成纤维细胞在结直肠癌发生与发展及化疗耐药中的作用研究进展[J]. 中华结直肠疾病电子杂志, 2023,12(3):241-247. [97] 李佳鑫, 孙燕. 结直肠癌免疫微环境中肿瘤相关巨噬细胞的作用[J]. 中国肿瘤临床, 2020,47(17):890-896. [98] 解广东, 王本军, 白克运, 等. 结直肠癌细胞免疫逃逸研究概况[J]. 中国中西医结合外科杂志, 2019,25(2):221-224. [99] Bhat AA, Nisar S, Singh M, et al. Cytokine- and chemokine-induced inflammatory colorectal tumor microenvironment: Emerging avenue for targeted therapy. Cancer Commun (Lond). 2022;42(8):689-715. doi:10.1002/cac2.12295 [100] Liu W, Kuang T, Liu L, Deng W. The role of innate immune cells in the colorectal cancer tumor microenvironment and advances in anti-tumor therapy research. Front Immunol. 15:1407449. Published 2024 None. doi:10.3389/fimmu.2024.1407449 [101] 党利敏, 马虎林. 调节性 T 细胞在结直肠癌免疫治疗中的意义[J]. 河北医药, 2024,46(2):279-283,288. [102] Maryam S, Krukiewicz K, Haq IU, Khan AA, Yahya G, Cavalu S. Interleukins (Cytokines) as Biomarkers in Colorectal Cancer: Progression, Detection, and Monitoring. J Clin Med. 2023;12(9). Published 2023 Apr 25. doi:10.3390/jcm12093127 [103] Gulubova M, Chonov D, Aleksandrova E, Ivanova K, Ignatova MM, Vlaykova T. Interleukin-6-Positive Immune Cells as a Possible New Immunologic Marker Associated With the Colorectal Cancer Prognosis. Appl Immunohistochem Mol Morphol. 2024 May-Jun 01;32(5):233-243. doi:10.1097/PAI.0000000000001198 [104] Li W, Chen F, Gao H, et al. Cytokine concentration in peripheral blood of patients with colorectal cancer. Front Immunol. 14:1175513. Published 2023 None. doi:10.3389/fimmu.2023.1175513 [105] 周云帆, 刘妙登, 刘欢, 等. 免疫细胞促进结直肠癌转移的研究现状与展望[J]. 中华实验外科杂志, 2025,42(3):398-404. [106] 余升红. 肿瘤 DNA 异常甲基化与组蛋白修饰[J]. 医学分子生物学杂志, 2004,1(5):288-291. [107] Norollahi SE, Foumani MG, Pishkhan MK, et al. DNA Methylation Profiling of MYC, SMAD2/3 and DNMT3A in Colorectal Cancer. Oman Med J. 2021;36(6):e315. Published 2021 Nov. doi:10.5001/omj.2020.93 [108] Menyhart O, Müller D, Győrffy B. DNA methylation-mediated extracellular matrix gene silencing in colorectal cancer. Semin Oncol. 2026;53(2):152469. doi:10.1016/j.seminoncol.2026.152469 [109] 张翠娟, 周庚寅. DNA 甲基化和组蛋白修饰在肿瘤中的作用[J]. 中国现代普通外科进展, 2004,7(6):333-334,336. [110] Kamal T, Biswas AUH, Bhuiyan AR, et al. Implication of Epigenetic Alterations of ZEB1 in Colorectal Cancer (CRC) Pathogenesis and Therapy Development. Curr Issues Mol Biol. 2026;48(3). Published 2026 Mar 4. doi:10.3390/cimb48030276 [111] Likasitwatanakul P, Li Z, Doan P, et al. Chemical Perturbations Impacting Histone Acetylation Govern Colorectal Cancer Differentiation. Gastroenterology. 2026;170(1):70-88. doi:10.1053/j.gastro.2025.07.003 [112] 高福利, 吕瑛, 朱银鑫, 等. DNA 甲基化和组蛋白修饰的关键酶在胃癌中存在相互作用[J]. 中华消化杂志, 2013,33(2):106-110. [113] Tahghighi A, Seyedhashemi E, Mohammadi J, et al. Epigenetic marvels: exploring the landscape of colorectal cancer treatment through cutting-edge epigenetic-based drug strategies. Clin Epigenetics. 2025;17(1):34. Published 2025 Feb 22. doi:10.1186/s13148-025-01844-w [114] Hussen BM, Sulaiman SHA, Abdullah SR, et al. MiRNA-155: A double-edged sword in colorectal cancer progression and drug resistance mechanisms. Int J Biol Macromol. 299:140134. doi:10.1016/j.ijbiomac.2025.140134 [115] Wang D, Wang J, Yao F, et al. miR-1247-3p regulation of CCND1 affects chemoresistance in colorectal cancer. PLoS One. 2024;19(12):e0309979. Published 2024 None. doi:10.1371/journal.pone.0309979 [116] Chen YP, Baskaran R, Devi HS, et al. miR-100-5p Enhances Cell Cycle-Mediated Chemoresistance by Modulating the CTDSPL/pRB/E2F1 Signaling Pathway in Oxaliplatin-Resistant Colorectal Cancer Cells. Oncol Res. 2026;34(4):17. Published 2026 None. doi:10.32604/or.2026.073080 [117] Li X, Chen X, Fu C, Xie M, Ouyang S. Long Non-Coding RNA BCAR4 Promotes Oxaliplatin Resistance in Colorectal Cancer by Modulating miR-484-3p/RAB5C Expression. Chemotherapy. 2023;68(3):119-130. doi:10.1159/000529134 [118] Tong D, Zou E, Bai L, et al. IRF-1 mediated long non-coding RNA PVT1-214 promotes oxaliplatin resistance of colorectal cancer via miR-128 inhibition. J BUON. 2020 Sep-Oct;25(5):2205-2214. [119] Wang X, Zhang H, Yang H, et al. Exosome-delivered circRNA promotes glycolysis to induce chemoresistance through the miR-122-PKM2 axis in colorectal cancer. Mol Oncol. 2020;14(3):539-555. doi:10.1002/1878-0261.12629 [120] Sahebnasagh R, Deli H, Shadboorestan A, et al. Identification of key lncRNAs associated with oxaliplatin resistance in colorectal cancer cells and isolated exosomes: From In-Silico prediction to In-Vitro validation. PLoS One. 2024;19(10):e0311680. Published 2024 None. doi:10.1371/journal.pone.0311680 [121] 王丹丹, 王文鑫, 张晓岚. 长链非编码 RNA 作为 ceRNA 在结直肠癌中的作用[J]. 胃肠病学和肝病学杂志, 2021,30(4):365-368. [122] Lei X, He Q, Li Z, et al. Cancer stem cells in colorectal cancer and the association with chemotherapy resistance. Med Oncol. 2021;38(4):43. Published 2021 Mar 18. doi:10.1007/s12032-021-01488-9 [123] 兰莉莉, 黎军, 孙琳, 等. 结直肠癌肿瘤干细胞标记物研究进展[J]. 成都医学院学报, 2021,16(1):127-130. [124] Liu C, Wang JL, Wu DZ, Yuan YW, Xin L. Methionine restriction enhances the chemotherapeutic sensitivity of colorectal cancer stem cells by miR-320d/C-Myc axis. Mol Cell Biochem. 2022;477(7):2001-2013. doi:10.1007/s11010-022-04416-1 [125] Huang JL, Oshi M, Endo I, Takabe K. Clinical relevance of stem cell surface markers CD133, CD24, and CD44 in colorectal cancer. Am J Cancer Res. 2021;11(10):5141-5154. Published 2021 None. [126] Rey I, Effendi-Ys R. Association between colorectal cancer stem cells’ expression of LGR5 and the location of colorectal cancer. Gastroenterol Hepatol Bed Bench. 2025;18(4):483-489. doi:10.22037/ghfbb.v18i4.3200 [127] 张欢乐, 张凤春, 王萍, 等. 结直肠癌干细胞微球体的培养及耐药机制[J]. 肿瘤, 2016,36(2): [128] Clarkson E, Lewis A. BMP antagonist CHRDL2 enhances the cancer stem-cell phenotype and increases chemotherapy resistance in colorectal cancer. Mol Oncol. 2025;19(11):3135-3155. doi:10.1002/1878-0261.70064 [129] Lu M, Hu X, Cheng C, et al. RPF2 mediates the CARM1‑MYCN axis to promote chemotherapy resistance in colorectal cancer cells. Oncol Rep. 2024;51(1). doi:10.3892/or.2023.8670 [130] Feng WQ, Zhang YC, Gao H, et al. FOXD1 promotes chemotherapy resistance by enhancing cell stemness in colorectal cancer through β‑catenin nuclear localization. Oncol Rep. 2023;50(1). doi:10.3892/or.2023.8571 [131] Wang L, Zhao L, Lin Z, et al. Targeting DCLK1 overcomes 5-fluorouracil resistance in colorectal cancer through inhibiting CCAR1/β-catenin pathway-mediated cancer stemness. Clin Transl Med. 2022;12(5):e743. doi:10.1002/ctm2.743 [132] Roca MS, Lombardi R, Testa C, et al. Valproic acid improves the efficacy of oxaliplatin/fluoropyrimidine-based chemotherapy by targeting cancer stem cell via β-Catenin modulation in colorectal cancer. Cell Death Dis. 2025;16(1):583. Published 2025 Aug 1. doi:10.1038/s41419-025-07902-8 [133] Cheung H, Chen H, Chen D, et al. YTHDF1 targets the chemotherapy response by suppressing NOTCH1-induced stemness in colorectal cancer. Signal Transduct Target Ther. 2025;10(1):409. Published 2025 Dec 22. doi:10.1038/s41392-025-02507-1 [134] Hong XL, Yu TC, Huang XW, et al. Metformin abrogates Fusobacterium nucleatum-induced chemoresistance in colorectal cancer by inhibiting miR-361-5p/sonic hedgehog signaling-regulated stemness. Br J Cancer. 2023;128(2):363-374. doi:10.1038/s41416-022-02044-6 [135] Li F, Zhou Y, Lin X, et al. A novel USP4 inhibitor that suppresses colorectal cancer stemness by promoting β-catenin and Twist1 degradation. J Transl Med. 2025;23(1):114. Published 2025 Jan 24. doi:10.1186/s12967-024-06001-0 [136] Ni Q, Li M, Yu S. Research Progress of Epithelial-mesenchymal Transition Treatment and Drug Resistance in Colorectal Cancer. Technol Cancer Res Treat. 21:15330338221081219. doi:10.1177/15330338221081219 [137] Kehagias P, Kindt N, Krayem M, et al. Regorafenib Induces Senescence and Epithelial-Mesenchymal Transition in Colorectal Cancer to Promote Drug Resistance. Cells. 2022;11(22). Published 2022 Nov 18. doi:10.3390/cells11223663 [138] Zhang C, Zeng M, Xu Y, et al. S100A6 mediated epithelial-mesenchymal transition affects chemosensitivity of colorectal cancer to oxaliplatin. Gene. 914:148406. doi:10.1016/j.gene.2024.148406 [139] Zhang Z, Zheng S, Lin Y, et al. Genomics and prognosis analysis of epithelial-mesenchymal transition in colorectal cancer patients. BMC Cancer. 2020;20(1):1135. Published 2020 Nov 23. doi:10.1186/s12885-020-07615-5 [140] Pastorino GA, Sheraj I, Huebner K, et al. A partial epithelial-mesenchymal transition signature for highly aggressive colorectal cancer cells that survive under nutrient restriction. J Pathol. 2024;262(3):347-361. doi:10.1002/path.6240 [141] Li R, Xu H, Gao X. The ceRNA network regulates epithelial-mesenchymal transition in colorectal cancer. Heliyon. 2023;9(3):e14143. Published 2023 Mar. doi:10.1016/j.heliyon.2023.e14143 [142] Zhu Y, Huang S, Chen S, et al. SOX2 promotes chemoresistance, cancer stem cells properties, and epithelial-mesenchymal transition by β-catenin and Beclin1/autophagy signaling in colorectal cancer. Cell Death Dis. 2021;12(5):449. Published 2021 May 5. doi:10.1038/s41419-021-03733-5 [143] Shao X, Sun Y, Zhong K, et al. TYRO3 promotes tumorigenesis and drug resistance in colorectal cancer by enhancing the epithelial-mesenchymal transition process. Aging (Albany NY). 2023;15(8):3035-3051. doi:10.18632/aging.204656 [144] Shi CJ, Xue ZH, Zeng WQ, et al. LncRNA-NEF suppressed oxaliplatin resistance and epithelial-mesenchymal transition in colorectal cancer through epigenetically inactivating MEK/ERK signaling. Cancer Gene Ther. 2023;30(6):855-865. doi:10.1038/s41417-023-00595-1 [145] Jiang Y, Xu Y, Zhu Q, et al. MYH11 Suppresses Colorectal Cancer Progression by Inhibiting Epithelial-Mesenchymal Transition via ZEB1 Regulation. Oncol Res. 2025;33(9):2379-2398. Published 2025 None. doi:10.32604/or.2025.063501 [146] Sun L, Xing J, Zhou X, Song X, Gao S. Wnt/β-catenin signalling, epithelial-mesenchymal transition and crosslink signalling in colorectal cancer cells. Biomed Pharmacother. 175:116685. doi:10.1016/j.biopha.2024.116685 [147] Li Y, Wang M, Jiang L, et al. SIPA1 promotes epithelial-mesenchymal transition in colorectal cancer through STAT3 activation. Heliyon. 2024;10(14):e34527. Published 2024 Jul 30. doi:10.1016/j.heliyon.2024.e34527 [148] Nenkov M, Ma Y, Gaßler N, Chen Y. Metabolic Reprogramming of Colorectal Cancer Cells and the Microenvironment: Implication for Therapy. Int J Mol Sci. 2021;22(12). Published 2021 Jun 10. doi:10.3390/ijms22126262 [149] 章丹宁, 杜利莉, 卞勇. 能量代谢重编程与结直肠癌的研究进展[J]. 中华肿瘤防治杂志, 2025,32(16):1015-1020. [150] Cheng Q, Chen Y, Zou D, et al. Targeting Metabolic Adaptation of Colorectal Cancer with Vanadium-Doped Nanosystem to Enhance Chemotherapy and Immunotherapy. Adv Sci (Weinh). 2025;12(7):e2409329. doi:10.1002/advs.202409329 [151] Jog E, Jainarayanan AK, La Ferlita A, et al. Inhibiting de novo lipogenesis identifies a therapeutic vulnerability in therapy-resistant colorectal cancer. Redox Biol. 79:103458. doi:10.1016/j.redox.2024.103458 [152] Chen D, Zhou X, Yan P, et al. Lipid metabolism reprogramming in colorectal cancer. J Cell Biochem. 2023;124(1):3-16. doi:10.1002/jcb.30347 [153] Marinescu R, Marinescu D, Preda D, et al. Oxidative Stress and Inflammation in Colorectal Cancer-Redox-Immune Crosstalk, Biomarkers, and Translational Implications: A Qualitative Systematic Review. Life (Basel). 2026;16(3). Published 2026 Mar 5. doi:10.3390/life16030424 [154] Ruangwannasak S, Khamsai S, Pakdeechote P, et al. Differential Oxidative Stress Profiles in Circulating and Peritumoral Adipose Tissue Across Stages of Colorectal Cancer. Int J Mol Sci. 2026;27(2). Published 2026 Jan 10. doi:10.3390/ijms27020707 [155] Peng L, Jiang J, Chen HN, et al. Redox-sensitive cyclophilin A elicits chemoresistance through realigning cellular oxidative status in colorectal cancer. Cell Rep. 2021;37(9):110069. doi:10.1016/j.celrep.2021.110069 [156] 盛宋昊然, 李玉斌, 郭家成, 等. 槐耳清膏通过氧化应激诱导结直肠癌细胞发生线粒体途径凋亡[J]. 中国中药杂志, 2024,49(12):3320-3329. [157] Bhasin N, Dabral P, Senavirathna L, Pan S, Chen R. Inhibition of TRAP1 Accelerates the DNA Damage Response, Activation of the Heat Shock Response and Metabolic Reprogramming in Colon Cancer Cells. Front Biosci (Landmark Ed). 2023;28(9):227. doi:10.31083/j.fbl2809227 [158] Leiphrakpam PD, Are C. PI3K/Akt/mTOR Signaling Pathway as a Target for Colorectal Cancer Treatment. Int J Mol Sci. 2024;25(6). Published 2024 Mar 9. doi:10.3390/ijms25063178 [159] Zhong J, Ding S, Zhang X, et al. To Investigate the Occurrence and Development of Colorectal Cancer Based on the PI3K/AKT/mTOR Signaling Pathway. Front Biosci (Landmark Ed). 2023;28(2):37. doi:10.31083/j.fbl2802037 [160] Su W, Feng B, Hu L, Guo X, Yu M. MUC3A promotes the progression of colorectal cancer through the PI3K/Akt/mTOR pathway. BMC Cancer. 2022;22(1):602. Published 2022 Jun 2. doi:10.1186/s12885-022-09709-8 [161] Gao M, Qian J, Xia P, et al. Targeting the KLF5/PI3K/AKT axis as a therapeutic strategy to overcome neoadjuvant chemoresistance in colorectal cancer. Front Immunol. 16:1593639. Published 2025 None. doi:10.3389/fimmu.2025.1593639 [162] Deng Z, Wu N, Suo Q, et al. Fucoidan, as an immunostimulator promotes M1 macrophage differentiation and enhances the chemotherapeutic sensitivity of capecitabine in colon cancer. Int J Biol Macromol. 2022;222(Pt A):562-572. doi:10.1016/j.ijbiomac.2022.09.201 [163] Su M, Zhan L, Zhang Y, Zhang J. Yes-activated protein promotes primary resistance of BRAF V600E mutant metastatic colorectal cancer cells to mitogen-activated protein kinase pathway inhibitors. J Gastrointest Oncol. 2021;12(3):953-963. doi:10.21037/jgo-21-258 [164] Koyama M, Kitazawa M, Nakamura S, et al. Low‑dose trametinib and Bcl‑xL antagonist have a specific antitumor effect in KRAS‑mutated colorectal cancer cells. Int J Oncol. 2020;57(5):1179-1191. doi:10.3892/ijo.2020.5117 [165] Morris VK, Bekaii-Saab T. Improvements in Clinical Outcomes for BRAFV600E -Mutant Metastatic Colorectal Cancer. Clin Cancer Res. 2020;26(17):4435-4441. doi:10.1158/1078-0432.CCR-19-3809 [166] Hirashita Y, Tsukamoto Y, Kudo Y, et al. Early response in phosphorylation of ribosomal protein S6 is associated with sensitivity to trametinib in colorectal cancer cells. Lab Invest. 2021;101(8):1036-1047. doi:10.1038/s41374-021-00590-w [167] Thakuri PS, Gupta M, Joshi R, Singh S, Tavana H. Synergistic Inhibition of Kinase Pathways Overcomes Resistance of Colorectal Cancer Spheroids to Cyclic Targeted Therapies. ACS Pharmacol Transl Sci. 2019;2(4):275-284. Published 2019 Aug 9. doi:10.1021/acsptsci.9b00042 [168] Koni M, Pinnarò V, Brizzi MF. The Wnt Signalling Pathway: A Tailored Target in Cancer. Int J Mol Sci. 2020;21(20). Published 2020 Oct 18. doi:10.3390/ijms21207697 [169] 孙纲, 韩大跃, 关宏伟, 等. Wnt/β-catenin 及 p38MAPK 信号通路在人胃癌组织中的表达情况与化学治疗预后及多药耐药相关性研究[J]. 成都医学院学报, 2017,12(1):28-32. [170] Zhao H, Chen S, Fu Q. Exosomes from CD133+ cells carrying circ-ABCC1 mediate cell stemness and metastasis in colorectal cancer. J Cell Biochem. 2020;121(5-6):3286-3297. doi:10.1002/jcb.29600 [171] Chen H, Pei L, Xie P, Guo G. Circ-PRKDC Contributes to 5-Fluorouracil Resistance of Colorectal Cancer Cells by Regulating miR-375/FOXM1 Axis and Wnt/β-Catenin Pathway. Onco Targets Ther. 13:5939-5953. Published 2020 None. doi:10.2147/OTT.S253468 [172] Zhang Z, Zhang Y, Qin X, Wang Y, Fu J. FGF9 promotes cisplatin resistance in colorectal cancer via regulation of Wnt/β-catenin signaling pathway. Exp Ther Med. 2020;19(3):1711-1718. doi:10.3892/etm.2019.8399 [173] 杨春勇, 梁庆模. Wnt 与 MAPK 信号通路在肿瘤发生中的串话[J]. 医学分子生物学杂志, 2010,7(5):441-447. [174] Jung HR, Oh Y, Na D, et al. CRISPR screens identify a novel combination treatment targeting BCL-XL and WNT signaling for KRAS/BRAF-mutated colorectal cancers. Oncogene. 2021;40(18):3287-3302. doi:10.1038/s41388-021-01777-7 [175] Fakhr E, Zare F, Azadmanesh K, Teimoori-Toolabi L. LEF1 silencing sensitizes colorectal cancer cells to oxaliplatin, 5-FU, and irinotecan. Biomed Pharmacother. 143:112091. doi:10.1016/j.biopha.2021.112091 [176] Zhang W, Peng C, Shen X, et al. A Bioactive Compound from Sanguisorba officinalis L. Inhibits Cell Proliferation and Induces Cell Death in 5-Fluorouracil-Sensitive/Resistant Colorectal Cancer Cells. Molecules. 2021;26(13). Published 2021 Jun 24. doi:10.3390/molecules26133843 [177] Ali Ibrahim Mze A, Abdul Rahman A, Ibahim MJ, Wan Mohamad Zain WNI. The role of glycoproteins in autophagy and apoptosis in colorectal cancer: A scoping review. Med Oncol. 2026;43(2):110. Published 2026 Jan 12. doi:10.1007/s12032-025-03197-z [178] 刘明顺, 方明治, 李敏. 结直肠癌靶向药物耐药机制研究进展[J]. 现代肿瘤医学, 2021,29(15):2730-2735. [179] Duta-Ion SG, Juganaru IR, Hotinceanu IA, et al. Redefining Therapeutic Approaches in Colorectal Cancer: Targeting Molecular Pathways and Overcoming Resistance. Int J Mol Sci. 2024;25(23). Published 2024 Nov 21. doi:10.3390/ijms252312507 [180] Favier S, Xiong W, Chalabi-Dchar M, et al. Exploring and bypassing resistance to targeted therapies in colorectal cancer. Biomed Pharmacother. 193:118741. doi:10.1016/j.biopha.2025.118741 [181] Valdenizia R. Silva, Luciano de S. Santos, Rosane B. Dias, 等. 靶向信号通路杀伤结直肠癌干细胞的新型药物[J]. 癌症, 2023,42(4):169-212. [182] Singh M, Morris VK, Bandey IN, Hong DS, Kopetz S. Advancements in combining targeted therapy and immunotherapy for colorectal cancer. Trends Cancer. 2024;10(7):598-609. doi:10.1016/j.trecan.2024.05.001 [183] 周黎明. 结直肠癌的靶向治疗药物研究进展[J]. 延安大学学报(医学科学版), 2022,20(1):1-9. [184] Al Bitar S, El-Sabban M, Doughan S, Abou-Kheir W. Molecular mechanisms targeting drug-resistance and metastasis in colorectal cancer: Updates and beyond. World J Gastroenterol. 2023;29(9):1395-1426. doi:10.3748/wjg.v29.i9.1395 [185] Wang K, Shen R, Meng T, Hu F, Yuan H. Nano-Drug Delivery Systems Based on Different Targeting Mechanisms in the Targeted Therapy of Colorectal Cancer. Molecules. 2022;27(9). Published 2022 May 6. doi:10.3390/molecules27092981 [186] 周立强, 辛林. 主动靶向纳米粒子治疗结直肠癌的研究进展[J]. 广东医学, 2019,40(8):1035-1039. [187] Piotrowska U, Szatko J, Nowakowska A, Klimaszewska E, Ogorzałek M, Sobczak M. Chitosan-Based Drug Delivery Systems for Targeted Chemotherapy in Colorectal Cancer: A Scoping Review. Mar Drugs. 2025;23(12). Published 2025 Dec 6. doi:10.3390/md23120467 [188] Leonard TE, Liko AF, Gustiananda M, Putra ABN, Juanssilfero AB, Hartrianti P. Thiolated pectin-chitosan composites: Potential mucoadhesive drug delivery system with selective cytotoxicity towards colorectal cancer. Int J Biol Macromol. 225:1-12. doi:10.1016/j.ijbiomac.2022.12.012 [189] Fatfat Z, Fatfat M, Gali-Muhtasib H. Micelles as potential drug delivery systems for colorectal cancer treatment. World J Gastroenterol. 2022;28(25):2867-2880. doi:10.3748/wjg.v28.i25.2867 [190] Li L, Chen M, Reis RL, Kundu SC, Xiao B, Shi X. Advancements of nanoscale drug formulations for combination treatment of colorectal cancer. Int J Pharm. 674:125508. doi:10.1016/j.ijpharm.2025.125508 [191] 王凯苑, 甘雅玲, 周岩, 等. 结直肠癌治疗药物的纳米化及临床转化研究进展[J]. 药学进展, 2017,41(11):824-829. [192] Zhang J, Ali K, Wang J. Research Advances of Lipid Nanoparticles in the Treatment of Colorectal Cancer. Int J Nanomedicine. 19:6693-6715. Published 2024 None. doi:10.2147/IJN.S466490 [193] Mao X, Li X, Liu S, et al. Global research trends on nanotechnology and colorectal cancer: a two-decade analysis. Nanomedicine (Lond). 2025;20(4):343-354. doi:10.1080/17435889.2025.2452833 [194] Cao W, Zhang X, Li R, et al. Lipid core-shell nanoparticles co-deliver FOLFOX regimen and siPD-L1 for synergistic targeted cancer treatment. J Control Release. 368:52-65. doi:10.1016/j.jconrel.2024.02.025 [195] Jia M, Yuan Z, Yu H, et al. Rapamycin circumvents anti PD-1 therapy resistance in colorectal cancer by reducing PD-L1 expression and optimizing the tumor microenvironment. Biomed Pharmacother. 176:116883. doi:10.1016/j.biopha.2024.116883 [196] Sun L, Patai ÁV, Hogenson TL, Fernandez-Zapico ME, Qin B, Sinicrope FA. Irreversible JNK blockade overcomes PD-L1-mediated resistance to chemotherapy in colorectal cancer. Oncogene. 2021;40(32):5105-5115. doi:10.1038/s41388-021-01910-6 [197] Rahimi A, Baghernejadan Z, Hazrati A, et al. Combination therapy with immune checkpoint inhibitors in colorectal cancer: Challenges, resistance mechanisms, and the role of microbiota. Biomed Pharmacother. 186:118014. doi:10.1016/j.biopha.2025.118014 [198] Shan J, Han D, Shen C, Lei Q, Zhang Y. Mechanism and strategies of immunotherapy resistance in colorectal cancer. Front Immunol. 13:1016646. Published 2022 None. doi:10.3389/fimmu.2022.1016646 [199] Saurav S, Karfa S, Vu T, et al. Overcoming Irinotecan Resistance by Targeting Its Downstream Signaling Pathways in Colon Cancer. Cancers (Basel). 2024;16(20). Published 2024 Oct 15. doi:10.3390/cancers16203491 [200] Wang L, Chen J, Chen Q, et al. The Gut Microbiota Metabolite Urolithin B Prevents Colorectal Carcinogenesis by Remodeling Microbiota and PD-L1/HLA-B. Oxid Med Cell Longev. 2023:6480848. Published 2023 None. doi:10.1155/2023/6480848 [201] Xiong L, Wei Y, Jia Q, et al. The application of extracellular vesicles in colorectal cancer metastasis and drug resistance: recent advances and trends. J Nanobiotechnology. 2023;21(1):143. Published 2023 Apr 29. doi:10.1186/s12951-023-01888-1 [202] Haynes J, Manogaran P. Mechanisms and Strategies to Overcome Drug Resistance in Colorectal Cancer. Int J Mol Sci. 2025;26(5). Published 2025 Feb 25. doi:10.3390/ijms26051988 [203] Wu C, Sun G, Wang F, et al. DYRK2 downregulation in colorectal cancer leads to epithelial-mesenchymal transition induction and chemoresistance. Sci Rep. 2022;12(1):22496. Published 2022 Dec 28. doi:10.1038/s41598-022-25053-0 [204] Wu CWK, Reid M, Leedham S, Lui RN. The emerging era of personalized medicine in advanced colorectal cancer. J Gastroenterol Hepatol. 2022;37(8):1411-1425. doi:10.1111/jgh.15937 [205] Yang Y, Jiang H, Li W, et al. FOXM1/DVL2/Snail axis drives metastasis and chemoresistance of colorectal cancer. Aging (Albany NY). 2020;12(23):24424-24440. doi:10.18632/aging.202300 [206] 赵岩, 周长春, 孙燕来. 类器官模型在结直肠癌耐药中的研究进展[J]. 中华肿瘤防治杂志, 2024,31(22):1406-1412. [207] Li X, Li T, Zhao Y, et al. Hsa_circ_0020095 modulates chemoresistance of CRC in a PDO model. Front Med (Lausanne). 12:1556611. Published 2025 None. doi:10.3389/fmed.2025.1556611 [208] Patsalias A, Kozovska Z. Personalized medicine: Stem cells in colorectal cancer treatment. Biomed Pharmacother. 141:111821. doi:10.1016/j.biopha.2021.111821 [209] 王熙, 吴川清. 结直肠癌多药耐药逆转的研究进展[J]. 国际肿瘤学杂志, 2023,50(1):42-46.
100 项与 抗PD-1抗体(Chulalongkorn University) 相关的药物交易
登录后查看更多信息
研发状态
10 条进展最快的记录, 后查看更多信息
登录
| 适应症 | 最高研发状态 | 国家/地区 | 公司 | 日期 |
|---|---|---|---|---|
| 肿瘤 | 临床前 | 泰国 | 2022-10-22 |
登录后查看更多信息
临床结果
临床结果
适应症
分期
评价
查看全部结果
| 研究 | 分期 | 人群特征 | 评价人数 | 分组 | 结果 | 评价 | 发布日期 |
|---|
No Data | |||||||
登录后查看更多信息
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
核心专利
使用我们的核心专利数据促进您的研究。
登录
或
临床分析
紧跟全球注册中心的最新临床试验。
登录
或
批准
利用最新的监管批准信息加速您的研究。
登录
或
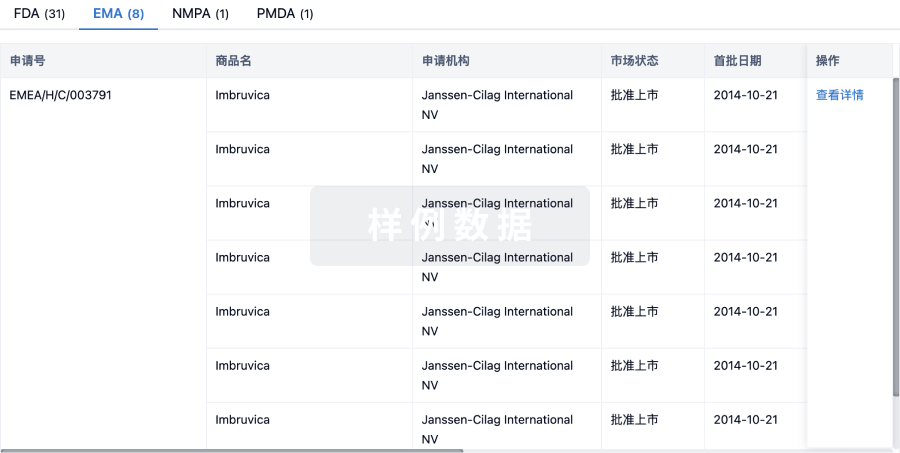
特殊审评
只需点击几下即可了解关键药物信息。
登录
或
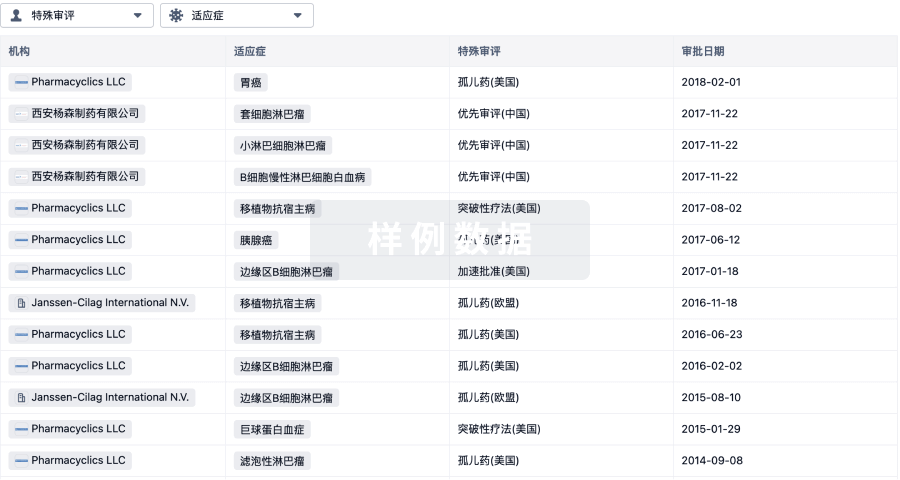
生物医药百科问答
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用