预约演示
更新于:2026-04-04
PF-06767832
更新于:2026-04-04
概要
基本信息
药物类型 小分子化药 |
别名 PF 06767832、PF-06767832、PF06767832 |
作用方式 调节剂 |
作用机制 M1 receptor调节剂(毒蕈碱型乙酰胆碱受体M1调节剂) |
治疗领域- |
在研适应症- |
非在研适应症- |
原研机构 |
在研机构- |
非在研机构 |
权益机构- |
最高研发阶段无进展药物发现 |
首次获批日期- |
最高研发阶段(中国)- |
特殊审评- |

结构/序列
分子式C22H23N3O3S |
InChIKeyVVZZHFMLRHJXTO-RXVVDRJESA-N |
CAS号1859081-58-5 |
关联
100 项与 PF-06767832 相关的临床结果
登录后查看更多信息
100 项与 PF-06767832 相关的转化医学
登录后查看更多信息
100 项与 PF-06767832 相关的专利(医药)
登录后查看更多信息
5
项与 PF-06767832 相关的文献(医药)2022-05-01·FASEB JOURNAL
Investigating Drivers for M
1
Muscarinic Acetylcholine Receptor‐Mediated Adverse Events by M
1
Positive Allosteric Modulators
Article
作者: van der Westhuizen, Emma T. ; Khajehali, Elham ; Christopoulos, Arthur ; Nguyen, Huong T. M. ; Valant, Celine
Introduction:
Targeting the allosteric site of the M
1
muscarinic acetylcholine receptor (M1 mAChR) is a promising strategy to develop selective drugs to treat cognitive disorders. By binding to the allosteric site, positive allosteric modulators of the M
1
mAChRs (M
1
PAMs) can enhance the binding (α) and/or the efficacy (β) of the endogenous ligand acetylcholine (ACh) (Fig 1), therefore improving impaired cholinergic transmission that is evident in Alzheimer’s disease and Schizophrenia. In addition, M
1
PAMs may also display direct allosteric agonism (t
B
) (Fig 1). To date, many highly selective M
1
PAMs with diverse structures have been developed, unfortunately, the majority of them still caused M
1‐
mediated cholinergic adverse effects (AEs), including gastrointestinal (GI) disturbances, salivation, increased heart rate and convulsions. This results from the ability of the M
1
mAChR to interact with multiple intracellular partners, leading to different functional outcomes, both the beneficial (i.e, improved cognition), and the detrimental (i.e, GI AEs) effects. Excitingly, M
1
PAMs can bias the M1 mAChR towards specific signalling outcomes versus others, a phenomenon called biased modulation. However, to date, it is unknown which intracellular partner is the most predominant in M
1
PAM activity, and how this may drive the preclinically observed AEs.
Aim:
To determine the role(s) Gα protein(s) and β‐arrestins exert on the allosteric activity of 5 structurally distinct M
1
PAMs and link these to their reported AEs.
Methods:
The M
1
PAMs including BQCA, MK7622, PF06767832, MIPS1780 which showed AEs in
in vivo
studies, and VU0486846 without observed AEs, were assessed against ACh, in parental or CRISPR HEK293A cells with deletion of specific G proteins or β‐arrestins (ΔGα
q/11,
ΔGα
12/13
or Δβ‐arrestin 1/2) expressing the human M
1
mAChR. [
3
H]‐NMS binding assays were performed to determine the binding affinity of ACh (K
A
), and of the 5 M
1
PAMs (K
B
) and their binding cooperativity (α) with ACh. IP
1
accumulation and b‐arrestin 2 recruitment were performed to quantify the efficacy (τ
B
) of the M
1
PAMs and their functional cooperativity (αβ) with ACh.
Results:
ΔGα
q/11
, ΔGα
12/13
or Δb‐arrestin 1/2 significantly reduced the binding affinity of ACh, and for 3 M
1
‐PAMs, MK7622, PF06767832 and MIPS1780. Interestingly, all these deletions significantly increased the binding cooperativity of these M
1
‐PAMs with ACh. Unsurprisingly, ΔGα
q/11
completely abolished IP1 responses from both orthosteric and allosteric ligands. Notably, ΔGα
12/13
or Δβ‐arrestin 1/2 both increased ACh efficacy (τ
A
). While ΔG
12/13
caused no change in efficacy of tested PAMs but increased functional cooperativity of PF06767832 and MIPS1780 with ACh, Δβ‐arrestin 1/2 affected the functional properties (τ
B
and αβ) of MK7622 in IP1 pathway. In β‐arrestin 2 pathway, ΔGα
q/11
increased MIPS1780 efficacy and ΔGα
12/13
increased functional cooperativity of BQCA.
Conclusion:
Gα
q/11
, Gα
12/13
and b‐arrestin 1/2 influence binding affinity and functional properties for both ACh and M
1
PAMs. These transductors affect M
1
PAMs with observed AEs more than the M
1
PAM without AEs, VU0486846, and thus playing a potential role in driving AEs.
2021-07-01·British Journal of Pharmacology
Functional brain imaging in larval zebrafish for characterising the effects of seizurogenic compounds acting via a range of pharmacological mechanisms
Article
作者: Andrew D. Randall ; Anna Tochwin ; Will S. Redfern ; Karen Tse ; Joseph Pinion ; Aya Takesono ; Piotr Grabowski ; Malcolm J. Hetheridge ; Jeremy Metz ; Maciej Trznadel ; Marc Goodfellow ; Jonathan S. Ball ; Matthew J. Winter ; Charles R. Tyler
Background and Purpose:
Functional brain imaging using genetically encoded Ca2+ sensors in larval zebrafish is being developed for studying seizures and epilepsy as a more ethical alternative to rodent models. Despite this, few data have been generated on pharmacological mechanisms of action other than GABAA antagonism. Assessing larval responsiveness across multiple mechanisms is vital to test the translational power of this approach, as well as assessing its validity for detecting unwanted drug‐induced seizures and testing antiepileptic drug efficacy.
Experimental Approach:
Using light‐sheet imaging, we systematically analysed the responsiveness of 4 days post fertilisation (dpf; which are not considered protected under European animal experiment legislation) transgenic larval zebrafish to treatment with 57 compounds spanning more than 12 drug classes with a link to seizure generation in mammals, alongside eight compounds with no such link.
Key Results:
We show 4dpf zebrafish are responsive to a wide range of mechanisms implicated in seizure generation, with cerebellar circuitry activated regardless of the initiating pharmacology. Analysis of functional connectivity revealed compounds targeting cholinergic and monoaminergic reuptake, in particular, showed phenotypic consistency broadly mapping onto what is known about neurotransmitter‐specific circuitry in the larval zebrafish brain. Many seizure‐associated compounds also exhibited altered whole brain functional connectivity compared with controls.
Conclusions and Implications:
This work represents a significant step forward in understanding the translational power of 4dpf larval zebrafish for use in neuropharmacological studies and for studying the events driving transition from small‐scale pharmacological activation of local circuits, to the large network‐wide abnormal synchronous activity associated with seizures.
2020-12-16·ACS chemical neuroscience3区 · 医学
Restoring Agonist Function at a Chemogenetically Modified M1Muscarinic Acetylcholine Receptor
3区 · 医学
Article
作者: Khajehali, Elham ; Lindsley, Craig W. ; van der Westhuizen, Emma T. ; Christopoulos, Arthur ; Tobin, Andrew B. ; Finlayson, Lisa ; Sexton, Patrick M. ; Valant, Celine ; Molloy, Colin ; Bradley, Sophie
Designer receptors exclusively activated by designer drugs (DREADDs) have been successfully employed to activate signaling pathways associated with specific muscarinic acetylcholine receptor (mAChR) subtypes. The M1 DREADD mAChR displays minimal responsiveness to the endogenous agonist acetylcholine (ACh) but responds to clozapine-N-oxide (CNO), an otherwise pharmacologically inert ligand. We have previously shown that benzyl quinolone carboxylic acid (BQCA), an M1 mAChR positive allosteric modulator (PAM), can rescue ACh responsiveness at these receptors. However, whether this effect is chemotype specific or applies to next-generation M1 PAMs with distinct scaffolds is unknown. Here, we reveal that new M1 PAMs restore ACh function at the M1 DREADD while modulating ACh binding at the M1 wild-type mAChR. Importantly, we demonstrate that the modulation of ACh function by M1 PAMs is translated in vivo using transgenic M1 DREADD mice. Our data provide important insights into mechanisms that define allosteric ligand modulation of agonist affinity vs efficacy and how these effects play out in the regulation of in vivo responses.
3
项与 PF-06767832 相关的新闻(医药)2026-03-24
胆碱酯酶抑制剂是目前针对阿尔茨海默病的一线对症治疗药物,但存在疗效低和不良反应的问题。M1毒蕈碱乙酰胆碱受体代表了一个潜在的替代治疗靶点;然而,针对这一G蛋白偶联受体的药物研发项目此前均告失败,主要原因在于胆碱能不良反应。利用新型化学遗传学和磷酸化缺陷型G蛋白偏向性小鼠模型,并结合一系列探针分子工具,研究人员发现了以往未被重视的、可通过药物干预的M1毒蕈碱乙酰胆碱受体神经过程,包括焦虑样行为和过度活动。通过绘制调控这些反应的上游信号通路,研究人员确定了受体磷酸化依赖性信号传导在驱动临床相关结果以及控制包括“癫痫样”惊厥在内的不良反应中的重要性。研究人员得出结论:能促进受体磷酸化依赖性信号传导的M1毒蕈碱乙酰胆碱受体配体,除了能驱动学习记忆和抗焦虑行为等对治疗阿尔茨海默病有益的反应外,还能预防胆碱能不良反应。
阿尔茨海默病是最常见的痴呆形式,在英国影响约85万人,在全球范围内影响5000万人。与许多形式的痴呆一样,阿尔茨海默病伴随一系列症状,不仅包括记忆丧失,还包括焦虑和激越等行为障碍。目前尚未出现能够同时应对阿尔茨海默病多种症状的单一治疗策略。相反,阿尔茨海默病症状治疗的基础策略主要集中于通过抑制乙酰胆碱酯酶(负责分解乙酰胆碱的酶)来恢复胆碱能传递,从而解决认知缺陷。然而,这种治疗的疗效受到严重剂量相关胆碱能不良反应的限制,这主要是由于全身(包括中枢和外周)胆碱能系统的整体上调所致。
一个被广泛考虑的替代策略是靶向M1毒蕈碱乙酰胆碱受体,这是响应乙酰胆碱的G蛋白偶联受体家族的五个成员之一,在临床前动物研究中显示出促认知效果。然而,尽管在临床试验中出现了一些积极的疗效迹象,偏向M1/M4的正位激动剂呫诺美林和双位激动剂GSK1034702仍因胆碱能不良反应而失败。另一种旨在通过增加对M1毒蕈碱乙酰胆碱受体的选择性来减少不良反应的策略,是靶向非保守的别构位点以正向调节受体活性,即所谓的正向别构调节剂。尽管已发现多种结构不同的M1毒蕈碱乙酰胆碱受体选择性正向别构调节剂,其中一些分子同样遇到了不良反应的问题,包括颞叶癫痫。这些研究突显了人们对M1选择性正向别构调节剂和正位/双位配体在决定临床有效反应与不良反应方面的微妙药理和结构特性严重缺乏认识。
此外,利用偏向信号传导的概念来规避曾阻碍药物开发的毒蕈碱不良反应的潜力,也同样缺乏深入的研究。为了充分发挥M1毒蕈碱乙酰胆碱受体偏向性配体的潜力,有必要解析介导临床相关M1毒蕈碱乙酰胆碱受体神经过程的体内信号通路,并将其与导致不良反应的通路区分开来。研究人员通过构建G蛋白偏向性M1毒蕈碱乙酰胆碱受体来应对这一挑战,具体方法是去除受体磷酸化位点,从而使受体与磷酸化/抑制蛋白依赖性信号传导脱钩。通过将该G蛋白偏向性受体敲入野生型M1毒蕈碱乙酰胆碱受体的基因位点,研究人员能够将神经和外周反应归因于G蛋白依赖性信号传导或受体磷酸化/抑制蛋白依赖性信号传导。
研究人员将这一方法与设计师药物专一性激活设计师受体技术的独特应用相结合,该技术在M1毒蕈碱乙酰胆碱受体的跨膜结构域3和5引入突变,产生了一种对天然配体乙酰胆碱无响应、但能被原本惰性的化学配体氯氮平-N-氧化物激活的受体突变体。尽管许多其他研究使用毒蕈碱型设计师受体来探究G蛋白信号传导在神经过程中的作用,但在此,通过将M1-设计师受体的编码序列敲入M1毒蕈碱乙酰胆碱受体基因位点,研究人员在一项旨在明确M1毒蕈碱乙酰胆碱受体作用及其“可成药性”的研究中,用M1-设计师受体替代野生型M1毒蕈碱乙酰胆碱受体进行表达。研究人员预测,这种突变小鼠的表型将模拟M1受体敲除小鼠,因为M1-设计师受体不对乙酰胆碱产生响应。然而,与M1受体敲除小鼠不同的是,在M1-设计师受体小鼠中,由M1毒蕈碱乙酰胆碱受体活性缺失导致的缺陷可通过给予氯氮平-N-氧化物得到纠正。通过这种方式,研究人员认为可以定义那些可能通过药物激活M1毒蕈碱乙酰胆碱受体来实现靶向的生理反应。利用这一方法,结合M1-PD小鼠以及表达磷酸化缺陷型M1-设计师受体的小鼠品系,研究人员在此阐述了,对M1毒蕈碱乙酰胆碱受体的药物靶向不仅影响学习和记忆,还能纠正焦虑相关行为和过度活跃的紊乱,这表明M1毒蕈碱乙酰胆碱受体配体有望治疗与阿尔茨海默病相关的广泛症状。通过绘制M1毒蕈碱乙酰胆碱受体神经反应和不良反应背后的双模信号通路,研究人员进一步得出结论:偏向于M1毒蕈碱乙酰胆碱受体磷酸化/抑制蛋白依赖性信号传导的配体,将在临床相关反应中发挥疗效,同时最大限度地减少包括“癫痫样”惊厥在内的不良反应。
一、结果
01.M1-设计师受体小鼠的构建
与先前研究一致,在中国仓鼠卵巢细胞中表达的人源野生型M1毒蕈碱乙酰胆碱受体,在肌醇磷酸积累实验中可被乙酰胆碱强效激活,而对氯氮平-N-氧化物的响应很弱(附表1)。表达M1-设计师受体的中国仓鼠卵巢细胞则表现出乙酰胆碱效价显著降低(5,000倍),同时对氯氮平-N-氧化物呈现纳摩尔级别的响应(附表1)。研究人员通过将M1毒蕈碱乙酰胆碱受体基因座进行靶向改造,经同源重组后用M1-设计师受体的编码序列替换M1毒蕈碱乙酰胆碱受体的编码序列(图1a和附图1a-c),由此获得的小鼠在海马和皮质中转录的M1-设计师受体信使RNA水平与野生型动物(指表达C端带有HA表位标签的野生型M1毒蕈碱乙酰胆碱受体的动物)中M1毒蕈碱乙酰胆碱受体的水平无显著差异(图1b)。此外,对照小鼠皮质中M1-设计师受体蛋白的表达水平与野生型受体相当,但在海马中的表达水平略高(图1c、d和附图2)。M1-设计师受体小鼠表现出正常的繁殖能力和健康指标。
图1 M1-设计师受体敲入小鼠的构建
a, M1受体的蛇形图,显示了为构建M1-设计师受体而引入的突变位点。
b, 逆转录定量聚合酶链式反应显示野生型或M1-设计师受体小鼠海马或皮质中M1毒蕈碱乙酰胆碱受体RNA的转录水平。数据表示为与β-肌动蛋白RNA转录的比值(\(n = 3\)只小鼠)。
c, d, 使用HA标签抗体,通过蛋白质印迹分析检测野生型M1-HA、M1-设计师受体和M1敲除小鼠海马(c)或皮质(d)制备的膜溶物中M1毒蕈碱乙酰胆碱受体的表达水平。所示数据为每种基因型各两只小鼠的结果。至少在另外两次独立实验中获得了类似数据(附图2)。Na⁺/K⁺-ATP酶的表达作为上样对照。
e, f, 在卡巴胆碱或氯氮平-N-氧化物刺激下,野生型(e)或M1-设计师受体(f)小鼠皮质膜制备物中[³⁵S]-GTPγS结合的刺激情况。数据表示为均值 ± 标准误,来自三到四次独立实验,每次实验设重复,并以野生型受体的最大响应进行归一化。
为了确定工程化小鼠中表达的M1-设计师受体对毒蕈碱配体的敏感性,研究人员在从皮质制备的膜中评估了M1-设计师受体与异源三聚体G蛋白的偶联情况。在这些实验中,氯氮平-N-氧化物在野生型动物的皮质膜中没有显示出显著活性,而毒蕈碱激动剂卡巴胆碱则使[³⁵S]GTPγS结合出现强劲增加(图1e和附表2)。相比之下,在M1-设计师受体小鼠的皮质膜中,氯氮平-N-氧化物引起Gq/11偶联的显著增强,而卡巴胆碱则未能刺激出反应(图1f和附表2)。
深圳市富临神通科技有限公司(原“东莞市富临塑胶原料有限公司”)是 Intan Technologies中国代理商,我们为客户提供Headstages、芯片、记录系统、刺激/记录系统、SPI线等电生理产品。
因此,在工程化小鼠中,M1-设计师受体的表达水平与野生型小鼠中的M1毒蕈碱乙酰胆碱受体接近相当,该受体不被天然配体乙酰胆碱激活,但能被氯氮平-N-氧化物激活。
02.氯氮平-N-氧化物降低M1-设计师受体小鼠的过度焦虑水平
在高架十字迷宫测试中,M1敲除动物表现出焦虑样表型增强,表现为与对照小鼠相比,进入开放臂的次数显著减少(图2a–g)。与对照相比,M1-设计师受体小鼠同样表现出进入开放臂的次数(占总进入次数的比例)减少,表明M1-设计师受体动物模拟了M1敲除小鼠的表型(图2a–g)。M1-设计师受体小鼠表现出的过度焦虑表型,通过在行为测试前30分钟给予氯氮平-N-氧化物(0.3 mg kg⁻¹)得以恢复到正常水平(图2a–g)。研究人员实验室先前的药代动力学研究确定,给予0.3 mg kg⁻¹氯氮平-N-氧化物(腹腔注射)可达到约50 nM的血浆暴露浓度,该浓度足以在体内完全激活病毒表达的毒蕈碱型设计师受体。重要的是,氯氮平-N-氧化物(0.3 mg kg⁻¹,腹腔注射)对野生型小鼠和M1敲除小鼠在高架十字迷宫测试中的表现均无影响(图2a–g)。这些数据表明,M1毒蕈碱乙酰胆碱受体功能缺失导致焦虑样表型,而该表型可通过选择性配体直接激活M1毒蕈碱乙酰胆碱受体来恢复。
图2 M1-设计师受体小鼠的表型改变可通过氯氮平-N-氧化物得到纠正
a–f, 热图显示野生型(a、d)、M1敲除(b、e)和M1-设计师受体(c、f)小鼠在高架十字迷宫测试中的活动分布,分别在接受溶剂对照(a–c)或0.3 mg kg⁻¹氯氮平-N-氧化物(d–f)(腹腔注射,测试前30分钟)给药后的情况。热图表示每治疗组2–6只小鼠的活动停留时间(蓝色,0秒停留;红色,5秒停留)。
g, 野生型、M1敲除或M1-设计师受体小鼠在高架十字迷宫测试前接受溶剂或氯氮平-N-氧化物处理后的平均焦虑水平(以开臂进入次数/闭臂进入次数与总进入次数的比值计算)。数据来自16–26只小鼠,采用双因素方差分析及Sidak多重比较检验进行统计学分析(*P < 0.05,**P < 0.01,***P < 0.001)。
h–j, 野生型(h)、M1敲除(i)或M1-设计师受体(j)小鼠在旷场测试中的代表性运动轨迹图。
k, 野生型、M1敲除或M1-设计师受体小鼠在旷场测试中10分钟内的平均运动距离。插图:每种基因型小鼠在10分钟内的总运动距离。数据表示为3–5只小鼠的均值。
l, 野生型、M1敲除或M1-设计师受体小鼠在Y迷宫测试前30分钟给予溶剂或0.3 mg kg⁻¹氯氮平-N-氧化物处理后,8分钟内的总运动距离。数据表示为均值 ± 标准误,来自4–13只小鼠,采用双因素方差分析及Sidak多重比较检验进行统计学分析(**P < 0.01;***P < 0.001)。
03.M1-设计师受体小鼠的过度活跃可通过氯氮平-N-氧化物恢复
与先前研究一致,研究人员证实M1敲除小鼠在旷场测试中表现出过度活跃(图2h–k;另见后续图4e、g–k)。由于M1-设计师受体对天然配体乙酰胆碱无响应,表达M1-设计师受体的小鼠同样表现出过度活动表型(图2h–k)。在Y迷宫范式测试中,M1敲除和M1-设计师受体动物也均表现出明显的过度活动(图2l和附图3)。重要的是,给予氯氮平-N-氧化物(0.3 mg kg⁻¹)对野生型或M1敲除小鼠的活动没有显著影响,但能将M1-设计师受体动物的过度活动降低至与溶剂处理的对照动物相似的水平(图2g、l和附图3)。这些数据表明,M1毒蕈碱乙酰胆碱受体不仅参与调节运动行为,而且通过选择性激动该受体亚型来调控运动行为是具有“可成药性”的。
图4 M1毒蕈碱乙酰胆碱受体生理反应的双模信号传导解析
a–c, 热图显示野生型(a,\(n = 13\))、M1敲除(b,\(n = 4\))或M1-PD(c,\(n = 14\))小鼠在高架十字迷宫开放臂或闭臂中的活动分布(蓝色,0秒停留;红色,5秒停留)。
d, 野生型(\(n = 21\))、M1敲除(\(n = 15\))或M1-PD(\(n = 14\))小鼠的焦虑水平(以开臂进入次数/闭臂进入次数与总进入次数的比值计算)。
e, 野生型(\(n = 13\))、M1敲除(\(n = 15\))或M1-PD(\(n = 14\))小鼠在高架十字迷宫中的总运动距离。
d和e中的数据以均值 ± 标准误表示,采用单因素方差分析与Dunnett多重比较检验进行统计学分析。** *\(P < 0.05\);***\(P < 0.001\)。
f, 野生型或M1-PD小鼠在Y迷宫自发交替范式测试中评估空间工作记忆,测试时长为8分钟。数据以均值 ± 标准误表示,采用Student's \(t\)-检验进行分析。*\(P < 0.05\)。
g–i, 野生型(g)、M1敲除(h)和M1-PD(i)小鼠在旷场测试中的代表性轨迹图。
j, 野生型、M1敲除和M1-PD小鼠在旷场测试中10分钟内的总运动距离。数据以均值 ± 标准误表示,采用单因素方差分析与Dunnett多重比较检验进行统计学分析。*\(P < 0.05\)。
k, 野生型、M1敲除和M1-PD小鼠的基础运动活动,通过体内遥测记录评估。数据展示了8只小鼠在24小时内的平均运动活动量 ± 标准误,该期间的总运动活动量通过曲线下面积计算得出。
l, 在野生型、M1敲除和M1-PD小鼠中测量给予毛果芸香碱(1 mg kg⁻¹)后的唾液分泌量。数据以均值 ± 标准误表示,\(n = 5–7\)只小鼠。数据分析采用单因素方差分析与Dunnett多重比较检验,与野生型小鼠进行比较。*\(P < 0.05\)。
m, M1毒蕈碱乙酰胆碱受体生理反应示意图,展示位于G蛋白依赖性信号下游的反应(唾液分泌和运动行为)与位于磷酸化依赖性信号下游的反应(抗焦虑行为和空间工作记忆)。
04.磷酸化缺陷型M1毒蕈碱乙酰胆碱受体小鼠的构建
与许多其他G蛋白偶联受体类似,毒蕈碱受体通过两条信号通路发挥作用——即通过异源三聚体G蛋白和受体磷酸化调控通路。后者包括受体与抑制蛋白接头蛋白的偶联,以及激活抑制蛋白依赖性过程(如受体内化)。研究人员构建了一个M1毒蕈碱乙酰胆碱受体突变体,该突变体缺失了质谱鉴定的所有磷酸化位点以及第三细胞内环和C末端尾部的其他潜在位点(共20个丝氨酸→丙氨酸替换;图3a),并在HEK293细胞中表达。该突变受体表现出强劲的Gq/11/钙动员偶联能力(图3b和附表3),同时显示出激动剂介导的抑制蛋白募集受体的能力减弱(图3c和附表3)以及受体内化缺陷(图3d和附图4)。这些结果与磷酸化缺陷型受体具有“G蛋白偏向性”的观点一致。
图3 G蛋白偏向性M1-PD敲入小鼠的构建
a, M1受体的蛇形图,显示了为构建M1-PD受体而引入的突变位点。
b, 在瞬时转染小鼠M1野生型或M1-PD毒蕈碱乙酰胆碱受体的HEK细胞中,刺激诱导的细胞内Ca²⁺释放情况。
c, 使用表达M1野生型或M1-PD受体的PathHunter HEK细胞测定β-抑制蛋白向M1毒蕈碱乙酰胆碱受体的募集情况。图中显示了对完全毒蕈碱受体激动剂氧化震颤素-M的浓度-响应曲线。数据以均值 ± 标准误表示,来自三次独立实验,并以野生型M1毒蕈碱乙酰胆碱受体对氧化震颤素-M的最大响应百分比表示。
d, 代表性图像显示,带HA标签的M1野生型或M1-PD在CHO细胞中的定位情况,细胞在固定前用溶剂或100 μM卡巴胆碱刺激1小时,并用4%多聚甲醛固定(×63物镜)。所示数据代表四次独立实验的结果。
e, [³H]-N-甲基东莨菪碱与野生型M1-HA或M1-PD小鼠制备的皮质膜的结合情况(\(n = 4\))。数据以均值 ± 标准误表示。
f, 逆转录定量聚合酶链式反应显示野生型和M1-PD小鼠海马或皮质中M1毒蕈碱乙酰胆碱受体RNA的转录水平。数据表示为与β-肌动蛋白RNA转录的比值(\(n = 3\)只小鼠)。
g, h, 使用HA标签抗体,通过蛋白质印迹分析检测野生型M1-HA、M1-PD和M1敲除小鼠海马(g)或皮质(h)制备的膜溶物中M1毒蕈碱乙酰胆碱受体的表达水平。所示数据为每种基因型各两只小鼠的结果;至少在另外两次独立实验中获得了类似数据(附图6)。Na⁺/K⁺-ATP酶的表达作为上样对照。
i, j, 在卡巴胆碱刺激下,野生型或M1-PD小鼠皮质组织中[³⁵S]-GTPγS与Gq/11的结合刺激(i)或Ins(1,4,5)P₃的积累情况(j)。数据以均值 ± 标准误表示,来自三次独立实验,[³⁵S]-GTPγS结合实验设重复,Ins(1,4,5)P₃积累实验设三重复,并以野生型受体的最大响应进行归一化。
研究人员构建了一种表达磷酸化缺陷型小鼠M1毒蕈碱乙酰胆碱受体的受体敲入小鼠(命名为M1-PD;附图5)。通过将该小鼠品系与野生型对照(表达C端带有标签的野生型M1毒蕈碱乙酰胆碱受体)及M1敲除动物进行比较,旨在区分位于受体磷酸化依赖性信号下游的生理反应与由G蛋白激活介导的生理反应。放射性配体结合实验确定,M1-PD小鼠皮质中的毒蕈碱受体表达量与野生型小鼠相当(图3e)。M1-PD小鼠皮质和海马中突变受体的转录水平与对照动物中M1毒蕈碱乙酰胆碱受体的水平无显著差异(图3f),蛋白质印迹结果定量分析显示,M1-PD小鼠海马和皮质中突变型M1毒蕈碱乙酰胆碱受体的表达水平无显著差异(图3g、h和附图6)。最后,在皮质组织提取物中,M1毒蕈碱乙酰胆碱受体与磷酸化缺陷型变体与异源三聚体G蛋白的偶联能力相当(通过[³⁵S]-GTPγS实验和肌醇1,4,5-三磷酸质量实验测定)(图3i、j和附表3)。
05.由M1毒蕈碱乙酰胆碱受体磷酸化调控的神经生理学
研究人员首先测试了M1-PD小鼠的抗焦虑反应,发现与M1敲除小鼠类似,M1-PD小鼠进入高架十字迷宫开放臂的次数更少(图4a–e)。这表明上述研究中揭示的抗焦虑反应是由受体磷酸化依赖性信号介导的。同样,在测试空间工作记忆的Y迷宫自发交替范式实验中,M1-PD小鼠表现出显著缺陷,表明与M1毒蕈碱乙酰胆碱受体介导的抗焦虑反应一样,这种行为也受到受体磷酸化的正向调控(图4f)。
最初在旷场测试中评估运动能力时,M1-PD小鼠并未表现出M1敲除小鼠的过度活跃表型(图4g–j)。相反,在该测试(图4j)和高架十字迷宫(图4e)中,M1-PD小鼠的总运动距离均少于野生型小鼠,表明这些小鼠表现为活动减退。有趣的是,尽管通过遥测技术监测24小时内的活动情况显示M1敲除小鼠过度活跃(与上述数据一致),但在此范式下,M1-PD小鼠的活动水平与野生型小鼠相当(图4k)。从这些运动研究中得出的结论是:与抗焦虑和空间工作记忆反应相反,运动行为的调控不依赖于受体磷酸化,而似乎是以G蛋白依赖性的方式调控的。在某些测试(高架十字迷宫和Y迷宫)中观察到的活动减退反应可能表明,尽管运动行为由G蛋白通路介导,但M1毒蕈碱乙酰胆碱受体介导的运动可能受受体磷酸化调控而发生脱敏,因此当磷酸化位点被移除时,表现为活动减退(图4e、j)。
研究人员接下来将这些中枢反应与M1毒蕈碱乙酰胆碱受体介导的唾液分泌进行了比较。如前所述,M1敲除小鼠对毒蕈碱受体激动剂毛果芸香碱的唾液分泌反应减弱,但M1-PD小鼠并未表现出这一现象(图4l),表明该反应同样位于G蛋白依赖性信号的下游(总结于图4m)。
06.M1毒蕈碱乙酰胆碱受体磷酸化可减少不良反应
为了进一步探究受体磷酸化依赖性信号在M1毒蕈碱乙酰胆碱受体介导反应中的作用,研究人员构建了一个去除磷酸化位点的M1-设计师受体(图5a)。在体外肌醇磷酸实验中,这种磷酸化缺陷型设计师受体被氯氮平-N-氧化物激活的效力和效能与可完全磷酸化的M1-设计师受体版本相当(附图7a和附表1)。与受体磷酸化在驱动磷酸化/抑制蛋白依赖性过程中的作用一致,磷酸化缺陷型M1-设计师受体在激动剂(氯氮平-N-氧化物)介导的受体内化方面存在缺陷(附图7b、c)。
图5 M1-DREADD PD小鼠表现出癫痫样惊厥
a, M1毒蕈碱乙酰胆碱受体的蛇形图,显示了为构建M1-DREADD PD受体而引入的突变位点。
b, 逆转录定量聚合酶链式反应显示野生型、M1-DREADD或M1-DREADD PD小鼠海马或皮质中M1毒蕈碱乙酰胆碱受体RNA的转录水平。数据表示为与β-肌动蛋白RNA转录的比值(\(n = 3\)只小鼠)。
c, d, 使用HA标签抗体,通过蛋白质印迹分析检测野生型、M1-DREADD、M1-DREADD PD和M1敲除小鼠海马(c)或皮质(d)制备的膜溶物中M1毒蕈碱乙酰胆碱受体的表达水平。所示数据为每种基因型各两只小鼠的结果。至少在另外两次独立实验中获得了类似数据(附图2)。Na⁺/K⁺-ATP酶的表达作为上样对照。
e, f, 在卡巴胆碱或氯氮平-N-氧化物刺激下,M1-DREADD(e)或M1-DREADD PD(f)小鼠皮质膜制备物中[³⁵S]-GTPγS结合刺激情况。数据以均值 ± 标准误表示,来自3–4次独立实验,每次实验设重复,并以野生型受体的最大响应进行归一化。
g, 代表性M1-DREADD PD小鼠在记录基础皮层活动15分钟后给予氯氮平-N-氧化物(0.3 mg kg⁻¹,腹腔注射)后的原始皮层脑电图信号(上图)及其频谱图(下图)。
h, M1-DREADD或M1-DREADD PD小鼠在接受溶剂(5%葡萄糖)或氯氮平-N-氧化物(0.3 mg kg⁻¹)处理后的归一化皮层脑电图功率。数据以均值 ± 标准误表示,来自4–8只小鼠,采用重复测量方差分析进行统计学分析,\(F(3, 3,119) = 5.53\),\(P = 0.029\)。
研究人员构建了一种表达磷酸化缺陷型M1-设计师受体以替代M1毒蕈碱乙酰胆碱受体的敲入小鼠,命名为M1-DREADD PD(附图8a、b)。M1-DREADD PD突变体的转录水平与M1-DREADD小鼠中的M1-设计师受体以及野生型小鼠中的M1毒蕈碱乙酰胆碱受体相当(图5b)。
可能由于M1-设计师受体不被内源性乙酰胆碱激活,该受体在工程化小鼠海马(而非皮质)中的表达水平略高于野生型受体(附图2)。M1-DREADD PD的表达水平也显著高于野生型受体(图5c、d和附图2)。重要的是,在海马或皮质中,M1-DREADD与M1-DREADD PD受体的表达水平之间无统计学显著差异(附图2)。这一点很重要,因为M1-DREADD作为M1-DREADD PD的对照(在观察M1-DREADD PD的表型时,研究人员不使用野生型小鼠作为对照)。
在来源于M1-DREADD和M1-DREADD PD的皮质膜中进行G蛋白偶联实验,结果显示氯氮平-N-氧化物在两种变体中均能强劲刺激Gq/11偶联增加,而卡巴胆碱则无显著增加(图5e、f和附表2)。当根据表达水平进行归一化时,研究人员估计M1-DREADD PD向Gq的信号传导比M1-DREADD高约1.5倍。
07.氯氮平-N-氧化物在M1-DREADD PD小鼠中诱发不良反应
此处的目的是探究在表达M1-设计师受体的动物中由氯氮平-N-氧化物给药所介导的生理反应,在M1-DREADD PD小鼠中是否同样产生响应。通过这种方式,研究人员可以进一步预测偏向性M1毒蕈碱乙酰胆碱受体选择性配体的影响。然而,对M1-DREADD PD小鼠给予氯氮平-N-氧化物导致了严重的不良反应(附表4),包括中枢和外周反应,如胃肠道紊乱和唾液过度分泌。这些不良反应此前曾与毒蕈碱药物研发项目的失败相关联。然而,最引人注目的是严重的惊厥发作,表现为胆碱能颞叶“癫痫样”惊厥的特征。研究人员进一步利用皮层脑电图记录来监测皮层活动,对这些惊厥进行了表征(图5g、h)。氯氮平-N-氧化物给药后,脑电图功率先降低,随后出现散发的惊厥活动,并在给药后约25–35分钟发展为完全的癫痫样发作。重要的是,相同剂量的氯氮平-N-氧化物(0.3 mg kg⁻¹)给予M1-DREADD小鼠时未产生不良反应(附表4)。
08.G蛋白偏向性系统表现出增强的不良反应
这些数据预示,与无偏向性配体相比,偏向于G蛋白依赖性信号而非受体磷酸化依赖性信号的毒蕈碱受体配体将表现出更多的不良反应。为了验证这一预测,研究人员对两种特征明确的毒蕈碱配体——毛果芸香碱和GSK1034702的G蛋白/受体磷酸化偏向性进行了分析。这两种配体均能刺激Gq/11依赖性信号,导致肌醇磷酸积累和细胞外信号调节激酶1/2的激活(图6a、b)。同样,毛果芸香碱和GSK1034702均能刺激M1毒蕈碱乙酰胆碱受体第三细胞内环中丝氨酸228的磷酸化,该位点先前被证实对激动剂介导的磷酸化高度敏感(附表5和图6c)。将浓度-响应曲线拟合至激动作用的操作模型后,研究人员得出了每种响应的转导系数。通过将这些系数与天然配体乙酰胆碱的转导系数进行比较,计算了两种配体在G蛋白偶联与受体磷酸化之间的偏向性,以偏向因子表示(附表6和图6d)。该分析确定毛果芸香碱表现出向G蛋白信号的偏向性,而GSK1034702在G蛋白偶联与受体磷酸化通路之间无偏向性(附表6和图6d)。
图6 毛果芸香碱表现出G蛋白偏向性,而GSK1034702无偏向性
a–c, 在稳定表达小鼠野生型M1毒蕈碱乙酰胆碱受体的CHO细胞中,由乙酰胆碱、毛果芸香碱或GSK1034702刺激的肌醇磷酸积累(a)、ERK1/2磷酸化(b)或M1毒蕈碱乙酰胆碱受体丝氨酸228位点磷酸化(c)。虚线显示作为先前数据集一部分生成的数据,以供比较。
d, 在野生型M1受体上,乙酰胆碱、毛果芸香碱和GSK1034702对肌醇磷酸、ERK1/2磷酸化或丝氨酸228磷酸化通路的优先信号偏向(\((\Delta\Delta\log_{10}(\tau/K_A))\))。数据以均值 ± 标准误表示,并以乙酰胆碱作为参考配体。
e,f, 给予递增浓度的GSK1034702(e)或毛果芸香碱(f)后,雄性Sprague–Dawley大鼠额叶皮质中肌醇磷酸积累的刺激情况。SKF38393是一种D1多巴胺受体激动剂。数据以均值 ± 标准误表示,来自3–4只大鼠,采用双因素方差分析及Dunnett多重比较检验进行统计学分析。*\(P < 0.05\),**\(P < 0.01\)与溶剂组相比;#\(P < 0.05\),##\(P < 0.01\)与氯化锂组相比。
为探究这些配体在体内是否表现出相似特性,研究人员分别给予毛果芸香碱(30 mg kg⁻¹)或GSK1034702(10 mg kg⁻¹),剂量设定为在大鼠中产生相当水平的受体占有率(60%),该占有率通过M1毒蕈碱乙酰胆碱受体正电子发射断层扫描示踪剂LSN317217634测定(附图9)。在此剂量下,毛果芸香碱引起强烈的肌醇磷酸反应,其强度约为GSK1034702观察到的反应的两倍(图6e、f),这支持了以下结论:在脑组织中,毛果芸香碱在刺激G蛋白依赖性信号方面比GSK1034702更有效。
随后,研究人员在野生型和M1-PD小鼠中,使用产生相当受体占有率的剂量,检测了这两种配体引发的不良事件。与偏向G蛋白信号的配体会表现出更显著不良反应的观点一致,研究人员观察到毛果芸香碱在野生型和M1-PD小鼠中均诱导了严重的中枢(震颤、抓握和惊厥)和外周(竖毛、流泪和腹泻)不良反应(附表7)。有趣的是,在毛果芸香碱作用下,唾液分泌是M1-PD小鼠中唯一反应低于野生型小鼠的反应(附表7)。尽管这可能提示唾液分泌位于受体磷酸化依赖性信号的下游,但图5l中的数据表明该反应完全由G蛋白介导。与毛果芸香碱相反,无刺激偏向性的GSK1034702在野生型小鼠中未表现出不良反应。仅当在M1-PD小鼠中(即受体呈现G蛋白偏向性的小鼠背景中),该配体才观察到中枢和外周不良反应(附表8)。
研究人员接下来利用其实验室先前的发现,即GSK1034702与M1毒蕈碱乙酰胆碱受体结合的双位特性意味着GSK1034702在野生型M1毒蕈碱乙酰胆碱受体和M1-设计师受体上均充当激动剂。这意味着该配体可独特地用作野生型小鼠以及M1-DREADD和M1-DREADD PD小鼠中的激动剂。在M1-DREADD小鼠中无不良反应的剂量(30 mg kg⁻¹)下给予GSK1034702,在M1-DREADD PD小鼠中引起了严重的外周和中枢不良反应(附表9),这一结果与以下观点一致:偏向G蛋白偶联的M1毒蕈碱乙酰胆碱受体信号转导会导致中枢和外周胆碱能不良反应。
二、讨论
在本研究中,我们构建了一系列新型化学遗传学和G蛋白偏向性小鼠模型,这些模型不仅揭示了磷酸化/抑制蛋白依赖性信号在M1毒蕈碱乙酰胆碱受体介导的学习记忆和抗焦虑行为中的重要性,还证实了M1毒蕈碱乙酰胆碱受体的磷酸化状态在减轻胆碱能不良反应方面具有显著作用。这些不良反应,无论是外周SLUDGE效应(流涎、流泪、排尿、排便、胃肠功能紊乱和呕吐)还是中枢不良反应(如惊厥),一直是靶向毒蕈碱受体治疗阿尔茨海默病成功道路上的主要障碍之一。此外,我们的数据表明,在阿尔茨海默病中靶向M1毒蕈碱乙酰胆碱受体,除了促认知的益处外,还可能对相关的行为异常(包括焦虑和过度活跃)产生影响。
通过采用设计师受体敲入小鼠,我们不仅模拟了M1毒蕈碱乙酰胆碱受体的药理学激活,还揭示了乙酰胆碱神经信号传导的某些特性,因为在阿尔茨海默病中受损的胆碱能张力,似乎可以通过药物模拟,从而在胆碱能传递受损的背景下实现“正常”的神经元活动和行为反应。
在本研究中,我们试图通过绘制G蛋白依赖性与磷酸化/抑制蛋白通路下游的反应(总结于图4m),来探究利用偏向性配体概念引导M1毒蕈碱乙酰胆碱受体信号转向治疗有益通路的潜力。为此,我们通过删除所有潜在的磷酸化受体位点,构建了G蛋白偏向性的M1毒蕈碱乙酰胆碱受体。有趣的是,去除这些磷酸化位点减少但并未消除受体与抑制蛋白的相互作用。这与其他G蛋白偶联受体亚型不同,在后者中去除磷酸化位点会导致受体/抑制蛋白相互作用几乎完全缺失。G蛋白偶联受体可通过与磷酸传感器相互作用的磷酸化依赖性方式以及通过激活传感器相互作用的方式与抑制蛋白结合。结构研究正在揭示这种双模结合的分子基础,特别是不同的磷酸化模式驱动不同抑制蛋白构象和信号输出的作用,这支持了磷酸化条形码的假说。在此,我们报告M1毒蕈碱乙酰胆碱受体与抑制蛋白的相互作用(类似于先前报道的M3毒蕈碱乙酰胆碱受体)仅部分依赖于受体的磷酸化状态,这表明抑制蛋白上的激活传感器在M1毒蕈碱乙酰胆碱受体与抑制蛋白的相互作用中发挥着与磷酸化传感器同等重要的作用。这在解释M1-PD小鼠的表型数据时可能具有重要意义,因为本文中鉴定为依赖于M1毒蕈碱乙酰胆碱受体磷酸化状态的表型,未必是由于受体/抑制蛋白相互作用缺陷所致。相反,受体磷酸化可能以不依赖于抑制蛋白的方式,对神经元中M1毒蕈碱乙酰胆碱受体信号复合物的正确组装/活性至关重要。
经典理论认为,G蛋白偶联受体的磷酸化与受体脱敏有关。因此,对M1-PD小鼠中观察到的表型的一种解释可能是受体无法进行磷酸化依赖性脱敏。运动反应的分析表明了这一点。与先前研究一致,我们证实M1敲除小鼠活动过度,表明M1毒蕈碱乙酰胆碱受体在降低运动方面发挥作用。然而,M1-PD小鼠活动减退,这一结果可能意味着磷酸化缺陷型受体本身过度活跃,可能是由于缺乏脱敏所致。
然而,当我们评估来自M1-PD小鼠组织的膜和皮质切片中磷酸化缺陷型受体与G蛋白信号的偶联时,仅观察到与G蛋白信号偶联的微弱增加。此外,本文中我们证明位于G蛋白偶联下游的唾液分泌在M1-PD小鼠中未受显著影响。这些数据指向一个有趣的假说:某些由G蛋白信号介导的生理反应(如运动)会被受体磷酸化所脱敏,而另一些(如唾液分泌)则不会。
在一系列利用G蛋白偏向性M1-设计师受体突变小鼠的实验中,偏向性信号对胆碱能不良反应的影响得到了惊人的揭示。在该小鼠品系中,我们发现给予氯氮平-N-氧化物(在野生型或M1-DREADD小鼠中不会引起不良反应的浓度)会导致严重的外周和中枢不良反应,包括惊厥。尽管先前研究明确表明不良反应可能与毒蕈碱配体的效能程度(包括正向别构调节剂的协同程度和内在活性)有关,但此前没有任何迹象表明受体磷酸化依赖性过程可能在调节不良反应中发挥作用。根据我们的数据,可以预见,信号偏向于G蛋白偶联而非受体磷酸化通路的配体可能表现出更明显的胆碱能不良反应。毛果芸香碱的情况即是如此,我们在此证明它既具有G蛋白偏向性,又具有显著的致惊厥活性(与先前研究一致)。相比之下,无偏向性的GSK1034702在给予与毛果芸香碱相同受体占有率的剂量时,在野生型小鼠中未引起惊厥。有趣的是,GSK1034702仅在磷酸化缺陷型、G蛋白偏向性小鼠(即M1-PD和M1-DREADD PD)中诱导了胆碱能不良反应。这些数据表明,偏向于受体磷酸化依赖性信号的毒蕈碱配体介导不良反应的倾向较低。
我们的研究进一步证实了越来越多的证据,即M1毒蕈碱乙酰胆碱受体可能是以前认为由M2/M3受体介导的外周不良反应的原因。因此,仅仅开发高选择性的M1毒蕈碱乙酰胆碱受体激动剂可能不足以避免胆碱能副作用。这一结论得到了相关报告的支持,即选择性M1毒蕈碱乙酰胆碱受体正向别构调节剂也表现出胆碱能不良反应。这些不良反应似乎与内在活性有关,因为具有高激动剂活性的正向别构调节剂也表现出胆碱能不良反应,而缺乏内在激动剂活性的正向别构调节剂则不诱导胆碱能毒性。目前尚不清楚,且肯定值得进一步研究的一点是,具有高激动剂活性和显著不良反应的正向别构调节剂是否也偏向于G蛋白信号传导。支持这一可能性的数据是,那些诱导不良反应的正向别构调节剂(如PF-06767832和PF-06827443)在纹状体组织中也能刺激强劲的肌醇磷酸信号(表明存在Gq信号传导)。
综合这些研究并考虑到我们的发现,我们得出结论:为了在针对阿尔茨海默病多种症状的治疗中最大限度地减少胆碱能副作用,同时实现最大的临床疗效,下一代M1毒蕈碱乙酰胆碱受体配体除了应具有高选择性和经过精细校准的效能外,还应能够驱动受体磷酸化依赖性信号传导。
三、方法
01.动物饲养
所有小鼠均以纯合子形式在C57BL/6J背景上繁育。除非另有说明,实验使用8-12周龄的雄性和雌性动物。小鼠可自由摄取标准鼠粮,并在实验前于动物设施中适应至少一周。动物护理遵循国家动物实验指南。所有实验均根据1986年《动物(科学程序)法》在英国内政部颁发的项目许可证下进行。
02.M1-PD、M1-DREADD和M1-DREADD PD敲入动物的构建
为构建敲入动物,研究人员构建了包含loxP-Stop-loxP盒的载体,该盒位于编码M1-HA、M1-PD、M1-DREADD或M1-DREADD PD的序列上游,并将其插入M1毒蕈碱乙酰胆碱受体基因(Chmr1)的编码外显子(外显子3)中。所有构建体均带有附加在C末端的HA表位序列(YPYDVPDYA)。M1-DREADD是人源化(人源化突变V5A、S254T、K320R、G337A和V413I)M1毒蕈碱乙酰胆碱受体的编码序列,带有Y106C和A196C两个突变(图1a)。M1-PD是小鼠M1毒蕈碱乙酰胆碱受体的编码序列,在第三细胞内环和C末端尾部具有将20个丝氨酸残基替换为丙氨酸的突变(图4a)。M1-DREADD PD是人源化M1-DREADD的编码序列,加上第三细胞内环和C末端尾部的20个丝氨酸→丙氨酸突变(图6a)。
包含毒蕈碱受体突变体编码序列的打靶载体随后转染至源自C57BL/6J小鼠的胚胎干细胞中,并筛选新霉素抗性胚胎干细胞。通过聚合酶链式反应和Southern印迹验证同源重组。将重组后的胚胎干细胞克隆注射入囊胚以产生嵌合小鼠。将嵌合小鼠与C57BL/6和表达Cre重组酶的小鼠交配,得以产生杂合子小鼠。杂合子动物通过交配产生纯合子品系。M1敲除小鼠是条件性M1-PD小鼠,其在M1-PD互补DNA上游表达两侧带有loxP位点的转录终止盒。如需获得这些小鼠,请联系通讯作者。M1-HA、M1-PD、M1-DREADD、M1-DREADD PD和M1敲除小鼠的构建由genOway公司完成。
需要注意的是,M1敲除品系是使用条件性M1-PD小鼠构建的,该品系未与任何Cre删除品系杂交,因此保留了起始密码子上游的终止盒完整,从而产生不表达受体的小鼠品系(构建体总结见附图4)。
03.逆转录定量聚合酶链式反应
使用Qiagen脂质组织RNeasy试剂盒,按照制造商说明,从野生型、M1-DREADD、M1-DREADD PD、M1-PD或M1敲除小鼠的海马或皮质中分离RNA。使用Nanodrop定量RNA浓度,每个反应使用1 μg总RNA模板,采用SuperScript III第一链合成SuperMix进行互补DNA合成。将RNA/水(共8 μl)、2 μl逆转录酶和10 μl 2×逆转录反应混合液混合,在25°C孵育10分钟,随后在50°C孵育30分钟,然后在85°C孵育5分钟。随后将样品在冰上冷却。每个反应均在存在和不存在逆转录酶的情况下进行(-逆转录对照)。最后,互补DNA与1 μl(2 U)大肠杆菌RNA酶H在37°C孵育20分钟,随后储存于-20°C直至进行逆转录定量聚合酶链式反应。
对于逆转录定量聚合酶链式反应,使用以下M1毒蕈碱乙酰胆碱受体引物(稀释度300:300):
F:5′ CAAGTGGCATTCATCGGGATCACC
R:5′ GAGAAAGTGCCAATGATGAGATCAGC
每个反应设三复孔。每个反应总体积为25 μl,包含12.5 μl SYBR Green Master Mix、0.75 μl F引物(10 μM储备液)、0.75 μl R引物(10 μM储备液)、10 μl水和1 μl互补DNA(或-逆转录样本)。
04.IP1积累实验
将人M1野生型或M1-DREADD PD构建体稳定表达于CHO-Flp-In细胞中,在含有10%胎牛血清和1%青霉素/链霉素的Ham's F12培养基中,在潮霉素B选择压力(400 μg ml⁻¹)下培养至T75 cm²培养瓶汇合。收集细胞,以1,000g离心3分钟,然后以1.43 × 10⁶细胞 ml⁻¹的浓度重悬于1×刺激缓冲液(HEPES,10 mM;CaCl₂,1 mM;MgCl₂,0.5 mM;KCl,4.2 mM;NaCl,146 mM;葡萄糖,5.5 mM;LiCl,50 mM;pH 7.4)中。将测试化合物(每孔7 μl)和细胞悬液(每孔7 μl)加入384孔白色ProxiPlate板中。短暂离心后,将板在37°C孵育45分钟。将IP1-d2缀合物和抗IP1穴状化合物Tb缀合物用裂解缓冲液按1:30稀释,每孔各加入3 μl。将板在37°C孵育1小时,使用Envision读板仪检测d2缀合IP1(发射波长665 nm)与Lumi4-Tb穴状化合物缀合抗IP1抗体(发射波长620 nm)之间的荧光共振能量转移。结果由665/620 nm比值计算得出,并以乙酰胆碱刺激的最大响应进行归一化。
05.ERK1/2磷酸化
使用CisBio Phospho-ERK细胞检测试剂盒评估ERK1/2磷酸化(Thr 202/Tyr 204)的刺激情况。将稳定表达人M1毒蕈碱乙酰胆碱受体的CHO Flp-In细胞培养至汇合单层,实验前进行血清饥饿过夜。用100 μl PBS洗涤细胞,在无血清F12培养基中于37°C孵育。用测试化合物在37°C下刺激细胞5分钟,终体积200 μl。通过快速吸弃培养液并加入50 μl补充有封闭试剂的裂解缓冲液终止刺激。裂解液在室温下轻轻振荡30分钟。随后,将16 μl该裂解液转移至384孔白色ProxiPlate板中,与4 μl预混的抗体溶液在室温下孵育2小时。使用PHERAstar读板仪测定荧光发射(665 nm和620 nm)。
06.细胞培养与转染(用于Ca²⁺和β-抑制蛋白募集实验)
将小鼠野生型或小鼠磷酸化缺陷型M1毒蕈碱乙酰胆碱受体(位于ProLink载体中)转染至PathHunter HEK293:β-arrestin:EA细胞中,并在抗生素(G418)筛选下培养,以获得稳定表达该受体的细胞池。将HEK293 wt/pdM1:β-arrestin:EA细胞维持在含有L-谷氨酰胺并添加胎牛血清(10%体积比)、潮霉素B(250 μg ml⁻¹)和遗传霉素G418(500 μg ml⁻¹)的DMEM培养基中,于37°C、5% CO₂条件下培养。
07.细胞内Ca²⁺测量
将PathHunter HEK293 wt/pdM1:β-arrestin细胞以每孔40,000个细胞、90 μl细胞培养基的密度接种于96孔透明底黑色板中,于37°C、5% CO₂下孵育过夜,形成汇合单层。实验当天,向96孔细胞板的每孔中加入30 μl含有0.02% pluronic酸:2.5 mM丙磺舒(1:1)的4× Ca²⁺无洗脱检测试剂盒4,于37°C、5% CO₂下孵育30分钟。随后使用FlexStation 3在时间维度上监测激动剂诱导的细胞内钙离子浓度变化。在加入一系列M1毒蕈碱乙酰胆碱受体激动剂之前,监测基础荧光16秒,之后继续记录荧光变化60秒。对激动剂的响应表示为从基线到峰值的荧光变化。最大荧光值取激动剂加入后初始峰的最高点,最小荧光值取激动剂加入前的背景荧光值。
08.抑制蛋白募集实验
将PathHunter HEK293 wt/pdM1:β-arrestin细胞以每孔8,000个细胞、20 μl细胞培养基的密度接种于白色透明底384孔ViewPlate板中,于37°C、5% CO₂下孵育过夜,形成汇合单层。实验当天,移除旧培养基,替换为含有0.1%牛血清白蛋白(重量/体积)和20 mM HEPES的HBSS缓冲液(pH 7.4)。用一系列M1毒蕈碱乙酰胆碱受体激动剂刺激细胞2小时(5 μl),之后加入25 μl专有Flash检测试剂,在室温下避光孵育15分钟。使用ClarioStar系统,采用无滤光片的发光程序读取发光信号。
09.蛋白质印迹分析
0901.膜提取物的制备
膜提取物的制备方案参考文献4、51中的膜制备方法。简述如下:将海马组织在含有蛋白酶抑制剂的25 mM磷酸钠缓冲液(pH 7.4)中以3–5 µg振幅进行超声匀浆。样品随后在4°C下以20,000g离心30分钟。将沉淀物与1.2%洋地黄皂苷溶于25 mM磷酸钠和5 mM MgCl₂缓冲液(pH 7.4)中,在4°C下过夜旋转孵育。样品在4°C下以20,000g离心30分钟后,将上清液(膜提取物)转移至新的微量离心管中,储存于-80°C备用。使用Micro BCA蛋白检测试剂盒,按照制造商说明测定蛋白浓度。
0902.蛋白质印迹分析
样品与含有5% β-巯基乙醇的Laemmli上样缓冲液在37°C下孵育30分钟,然后上样至7.5% SDS-Tris-甘氨酸聚丙烯酰胺凝胶中。样品在±100 V电压下运行,随后转移至硝酸纤维素膜上,膜用含5%脱脂奶粉的TBS-T(含0.1% Tween-20的TBS,pH 7.4)封闭2小时。随后将膜与相应的一抗在4°C下孵育过夜,然后用TBS-T洗涤三次(每次10分钟),并与辣根过氧化物酶偶联的相应二抗(1:5,000)孵育。使用ECL检测系统进行蛋白可视化。
0903.[³H]-NMS结合实验
将小鼠海马或皮质的膜制备物(每管50 μg)与含有递增浓度(0.1–5 nM)[³H]-NMS的结合缓冲液(HEPES,50 mM;NaCl,110 mM;KCl,5.4 mM;CaCl₂,1.8 mM;MgSO₄,1 mM;葡萄糖,25 mM;蔗糖,58 mM;pH 7.4)在37°C下孵育1小时。通过快速过滤至GF/B玻璃微纤维滤纸上,并用冰冷的0.9% NaCl洗涤三次,将膜结合配体与游离配体分离。通过液体闪烁计数测定膜结合放射性。非特异性结合通过在孵育[³H]-NMS时加入阿托品(1 μM)来确定。
0904.[³⁵S]-GTPγS实验
人道处死M1野生型、M1-DREADD、M1-PD和M1-DREADD PD小鼠(8–12周龄),然后在冰上解剖皮质组织。将组织悬浮于冰冷的缓冲液A(含0.9%(重量/体积)NaCl、10 mM HEPES、0.2%(重量/体积)EDTA,pH 7.4)中,使用Polytron匀浆器匀浆(4 × 5秒脉冲)。使用Eppendorf 5810R台式离心机在4°C下以200g离心5分钟。收集上清液并按上述方法重新匀浆。随后使用Beckman Coulter Avanti JXN-26离心机(JA-25.25转子)在4°C下以40,000g离心20分钟。弃去上清液,将沉淀重悬于10 ml冰冷的缓冲液B(10 mM HEPES,10 mM EDTA,pH 7.4)中。匀浆沉淀,加入GTP(终浓度1 mM),将悬液在37°C下孵育15分钟。随后在4°C下以40,000g离心20分钟,将沉淀重悬于15 ml冰冷的缓冲液C(10 mM HEPES,0.1 mM EDTA,pH 7.4)中,并如前所述重新匀浆。悬液再次在4°C下以40,000g离心20分钟。将最终沉淀重悬于缓冲液C中,使用Bradford法估算蛋白浓度。然后将匀浆液在最终储存缓冲液中进一步稀释,得到2 mg ml⁻¹的浓度。
[³⁵S]-GTPγS结合和Gα亚基免疫沉淀按先前描述进行。具体而言,将M1野生型、M1-DREADD、M1-PD或M1-DREADD PD膜稀释于含有终浓度1 μM GDP的分析缓冲液(HEPES,10 mM;NaCl,100 mM;MgCl₂,10 mM;pH 7.4)中。将膜(75 μg,总分析体积200 μl)加入[³⁵S]-GTPγS(终浓度1 nM)和激动剂(卡巴胆碱或氯氮平-N-氧化物)中,在30°C下孵育5分钟。通过加入1 ml冰冷的分析缓冲液并立即转移至冰浴终止反应。样品离心(20,000g,6分钟,4°C),通过加入50 μl冰冷的裂解缓冲液(100 mM Tris HCl,200 mM NaCl,1 mM EDTA,1.25% Igepal和0.2% SDS,pH 7.4)并在摇床上于4°C孵育1小时,使膜沉淀溶解。完全重溶蛋白后,加入50 μl不含SDS的裂解缓冲液。使用稀释度为1:100的正常兔血清和TE缓冲液(10 mM Tris HCl,10 mM EDTA,pH 8.0)中的3%(重量/体积)蛋白A-琼脂糖珠,在4°C下孵育60分钟,预清除溶解蛋白。通过离心(20,000g,6分钟,4°C)收集蛋白A-琼脂糖珠和不溶物,将100 μl上清液转移至含有Gq特异性抗血清的新管中,在4°C下孵育过夜。向样品中加入蛋白A-琼脂糖珠,涡旋混合,在4°C下旋转90分钟,然后离心(10,000g,1分钟,4°C)。吸去上清液,用冰冷的裂解缓冲液(不含SDS)洗涤蛋白A-琼脂糖珠三次。将回收的珠子与1 ml FloScint-IV闪烁液混合,通过液体闪烁计数测定放射性。
0905.Ins(1,4,5)P₃质量实验
通过颈椎脱臼法人道处死M1-HA或M1-PD小鼠。暴露大脑并转移至冰冷的平皿中,解剖大脑皮层。使用McIlwain组织切碎机将大脑皮层交叉切割成300 μm × 300 μm的碎块。将所得组织碎块分散于Krebs-Henseleit缓冲液中,通过多次更换缓冲液洗涤,然后在振荡水浴中于37°C振荡60分钟。在此期间,让组织碎块自然沉降,每10分钟更换一次缓冲液。
洗涤期结束后,让大脑皮层碎块自然沉降,将25 μl“压积”组织等分试样转移至含有250 μl KHB的平底5 ml管中。每管用O₂/CO₂(95:5)充气,盖上盖子,放回37°C振荡水浴中。加入药物(使总孵育体积达到300 μl),再次用O₂/CO₂(95:5)充气,并按图注所示时间继续孵育。加入等体积冰冷的1 M三氯乙酸终止孵育,管子在冰上提取30分钟。随后离心(2,000g,20分钟,4°C)。回收上清液,使用二氯二氟甲烷/三正辛胺法中和,并如前所述精确测定肌醇1,4,5-三磷酸浓度。通过加入1 M NaOH溶解组织碎块沉淀。使用Lowry法测定每次孵育的蛋白浓度。这使得IP₃质量积累可表示为每毫克蛋白的pmol IP₃。
0906.高架十字迷宫
小鼠在高架十字迷宫测试室中适应过夜,测试前保持黑暗状态。对于接受溶剂(5%葡萄糖)或氯氮平-N-氧化物(0.3 mg kg⁻¹)处理的小鼠,在测试前30分钟通过腹腔注射给药。
高架十字迷宫由四个不透明的臂(50 × 10 cm)组成:两个闭臂(黑色壁,高30 cm)和两个光照较弱的开臂。将小鼠放置于高架十字迷宫的中心,面向闭臂。使用ANY-maze软件追踪小鼠5分钟,并监测在此期间进入闭臂或开臂的次数。焦虑水平计算为开臂进入次数占总进入次数的百分比。每只动物测试后用70%乙醇清洁迷宫。
10.旷场
在行为测试套件中过夜适应后,使用旷场测试评估一般运动活动。将小鼠放置于透明有机玻璃方形场地(50 × 50 cm)中,使用ANY-maze软件记录10分钟内的活动。
11.Y迷宫
小鼠在测试前于行为测试套件中过夜适应。对于接受溶剂(5%葡萄糖)或氯氮平-N-氧化物(0.3 mg kg⁻¹)处理的测试,在测试开始前30分钟通过腹腔注射给药。将小鼠放置于Y迷宫(灰色、非反射底板)的中心,该迷宫有三个相同的臂(A、B和C;道宽5 cm,臂长35 cm,臂高10 cm)。使用ANY-maze软件记录活动。自发交替行为通过测量“ABC”序列(任意顺序)的数量占8分钟测试期间总三联体序列的比例来计算。
12.体内遥测
使用遥测系统在完整的自由活动的野生型、M1-PD和M1敲除小鼠中测量基础运动活动。在异氟烷(1.5–2%)麻醉和卡洛芬(5 mg kg⁻¹,皮下注射)镇痛下,将TA-F10植入式探头植入腹腔。植入期间,将小鼠置于恒温垫上。手术后,小鼠单笼饲养,恢复一周后进行实验。连续三天直接从应答器获取基础运动活动数据,期间不打扰动物。在家笼中记录运动活动。接收器通过MX2矩阵直接连接到PC的单个计算机端口,从而可以测定所有参数。每60秒收集一次数据,使用Ponemah采集系统进行数据收集和初步处理。
13.生物节律分析
将Ponemah采集系统收集的遥测实验数据按10分钟序列分组,计算平均值用于进一步分析。使用ChronosFit程序进行分析,采用傅里叶分析和逐步回归技术。
14.脑电图记录
1401.手术与记录
动物(M1-HA野生型、M1-DREADD、M1-DREADD PD)使用异氟烷(1–1.5%)麻醉,并置于立体定位框架中。使用电动推剪剃除动物头部毛发,并用乙醇(70%)和聚维酮碘清洁。在切口部位皮下注射利多卡因(2%,0.1–0.3 mg),并皮下注射卡洛芬(5 mg kg⁻¹)以提供术后镇痛。整个手术过程中使用加热垫维持动物体温在37°C。在颅骨上固定五颗骨钉,两颗位于额部区域(前囟后+1.5 mm,前囟中侧±1 mm)用作额叶皮层脑电图电极,两颗位于顶部区域(前囟后−2 mm,前囟中侧±2 mm)用作顶叶皮层脑电图电极,一颗位于小脑作为接地和参考电极。带有螺钉的电极用牙科水泥固定作为头部固定装置。头部固定装置手术后,动物至少恢复五天方可进行实验。在五天的适应期内,每天对动物进行抓取操作,将其置于记录室中,并将动物头部与记录线缆连接。适应期后的第二天,将动物放回记录室进行电生理记录。
记录过程如下:最初15分钟用于记录基础皮层活动。随后给小鼠腹腔注射溶剂(5%葡萄糖)或氯氮平-N-氧化物(0.3 mg kg⁻¹),并继续记录脑电图45分钟。在整个实验过程中,小鼠可自由活动,并监测其是否发生惊厥。
动物用异氟烷(1–1.5%)麻醉后,置于立体定位框架中,然后使用反馈温度控制器维持体温在37°C。在切口部位皮下注射利多卡因(2%,0.1 ml),并在背部皮下注射卡洛芬(5 mg kg⁻¹)。切口后,暴露并清洁颅骨。植入四颗骨钉用于皮层脑电图记录。另一颗螺钉植入小脑上方作为接地和参考电极。所有螺钉与连接器相连,并用牙科水泥覆盖。动物至少恢复五天。
电生理记录流程详见其他文献。简述如下:将动物置于开放箱中,通过连接16通道放大器板(RHD2132,Intan Technologies)和接口线缆进行记录。信号相对于小脑骨钉进行放大,并以1,000 Hz的频率数字化(RHD2132和RHD 2000,Intan Technologies)。每次记录包括15分钟基线记录、腹腔注射(氯氮平-N-氧化物0.3 mg kg⁻¹或溶剂)以及至少45分钟的再次记录。
所有离线分析使用MATLAB(版本R2018b)进行。由于所有四个脑电图通道提供的信号质量相似,仅使用右侧额叶区域的信号。为计算频谱图,应用多窗口谱估计方法(http://chronux.org/)。为评估信号功率,每1秒计算一次均方根值,并以dB为单位缩放。将缩放后的信号经1/300 Hz低通滤波后,相对于基线(前5分钟信号的平均值)进行归一化,以便在不同实验之间进行比较。
1402.唾液分泌测量
通过腹腔注射氯胺酮(100 mg kg⁻¹)/美托咪定(0.25 mg kg⁻¹)麻醉小鼠。之后,给小鼠注射毛果芸香碱(1 mg ml⁻¹,腹腔注射),并在35分钟内每5分钟记录一次GF/B滤纸上的唾液分泌量(以毫克计)。
1403.受体内化的免疫细胞化学
将稳定表达HA标签(C末端)小鼠M1野生型、小鼠M1-PD、人源化M1-DREADD或人源化M1-DREADD PD的CHO细胞培养24小时,在涂有0.01%多聚-d-赖氨酸的13 mm玻璃盖玻片上达到60–80%汇合度。用100 µM卡巴胆碱(野生型和M1-PD)或氯氮平-N-氧化物(M1-DREADD或M1-DREADD PD)刺激细胞1小时,用4%多聚甲醛(在TBS缓冲液中)固定,并用含0.1% Triton X-100的TBS缓冲液中的2%牛血清白蛋白进行封闭和透化处理。与抗EEA1多克隆抗体在4°C下孵育过夜,与抗HA抗体在室温下孵育2小时。用TBS缓冲液洗涤三次后,与AlexaFluor 594抗兔和AlexaFluor 488抗兔二抗在室温下孵育2小时,然后用TBS快速洗涤三次。将带有染色细胞的盖玻片用含DAPI的VECTASHIELD HardSet抗淬灭封片剂封片于载玻片上。使用LSM 880共聚焦激光扫描显微镜获取数据。
15.大鼠肌醇磷酸积累
1501.样本收集
大鼠(Sprague–Dawley,250–275 g)脑样本(额叶皮质)在动物接受以下处理后收集:水溶剂或LiCl(100 mg kg⁻¹,皮下注射),30分钟后给予水溶剂或东莨菪碱(1 mg kg⁻¹,皮下注射)或SKF38393(20 mg kg⁻¹,皮下注射),给药体积为1 ml kg⁻¹。LiCl给药后一小时,给大鼠注射毛果芸香碱(10、30或100 mg kg⁻¹,皮下注射)或GSK1034702(3、10或30 mg kg⁻¹,腹腔注射)。两小时后处死大鼠,在干冰上收集额叶皮质。样品储存于-70°C,用于肌醇磷酸的液相色谱-串联质谱分析。
1502.质谱分析
样品用探头超声破碎仪在8级强度下匀浆10秒,匀浆液为5倍体积的含0.1%甲酸和50%甲醇的50%乙腈。然后样品以13,000g离心12分钟。将100 μl上清液转移至200 μl蒸馏水中,取20 μl等分试样进样至液相色谱/质谱仪。使用肌醇2-单磷酸双(环己基铵)盐在50%乙腈+50%甲醇+0.1%甲酸中制备标准品(10、30、100、300和1,000 ng g⁻¹或ml⁻¹)。肌醇磷酸分析使用Agilent 6410系列三重四极杆液相色谱-串联质谱仪,配备MassHunter数据分析软件,安装电喷雾离子源,以负离子模式运行。通过监测肌醇磷酸的前体离子(质荷比为259)并靶向其产物离子(质荷比设为78.9)进行检测。色谱分离采用Zorbax RX-SIL高效液相色谱柱,流动相为含3%乙腈的水溶液,总体含0.1%甲酸,流速为0.7 ml min⁻¹。每次进样后均可见到与标准品保留时间和预期分子量相符的清晰色谱峰。根据这些峰面积对分析物进行定量。
16.体内受体占有率
1601.活体阶段
雄性Sprague–Dawley大鼠(每个剂量组n=4)购自Harlan,体重在200至300 g之间。给予毛果芸香碱或GSK1034702,剂量为0.03、0.1、0.3、1、3和10 mg kg⁻¹,以生成剂量反应曲线。动物仅接受溶剂或受试化合物,给药体积为10 ml kg⁻¹。在剂量反应研究中,大鼠在溶剂或化合物给药30分钟后,通过尾静脉静脉注射非标记示踪剂LSN317217634,剂量为10 mg kg⁻¹,给药体积为大鼠0.5 ml kg⁻¹、小鼠5 ml kg⁻¹。示踪剂给药20分钟后,通过颈椎脱臼处死动物。取出大脑并进行解剖。额叶皮质和小脑用于示踪剂测量,剩余大脑和血浆用于化合物暴露分析。受体占有率被认为是在示踪剂给药时间点t测定的。研究在Covance Alnwick或Greenfield进行。
1602.组织制备与示踪剂分析
称量额叶皮质和小脑样本,置于冰上的锥形离心管中。每管加入四倍体积(重量/体积)含0.1%甲酸的乙腈。然后使用超声探头匀浆样品,并使用台式离心机以14,000 r.p.m.离心20分钟。取50 μl上清液加入96孔板中的150 μl无菌水中进行稀释,用于液相色谱-串联质谱分析。使用API 4000质谱仪进行LSN3172176的分析。
色谱分离采用Agilent Zorbax Eclipse XDB-C18柱,流动相为梯度洗脱的15–90%乙腈水溶液,总体含0.1%甲酸。通过监测前体离子到产物离子的跃迁(质荷比386.3 → 128.0)来完成LSN3172176的检测。通过向未经处理的大鼠或小鼠脑组织样本中加入已知量的示踪剂,并按上述方法处理,制备标准品。
1603.受体占有率测定
受体占有率采用比值法计算。测量每个皮质和小脑样本中的示踪剂水平。对每只动物生成皮质水平(总结合)与小脑水平(非特异性结合)的比值。溶剂处理组的比值代表0%占有率,比值为1(即皮质结合等于小脑结合)代表100%占有率。毛果芸香碱和GSK1034702预处理组的比值在溶剂处理组比值(0%占有率)和1(100%占有率)之间进行线性插值,以确定M1受体占有率的百分比。对于毛果芸香碱和GSK1034702的剂量反应,使用GraphPad Prism 6.0版将曲线拟合为四参数逻辑函数,底部和顶部分别固定在0%和100%,并由软件计算达到50%受体占有率的剂量。数值以均值±标准误表示。
17.数据分析
功能性浓度-响应曲线根据四参数逻辑方程进行拟合(以确定最小和最大渐近线、对数半最大有效浓度及斜率;GraphPad Prism 6)。为评估激动剂偏向性,采用修正形式的激动作用操作模型对相同的浓度-响应曲线进行分析,重新构建后直接得出转导比(log(τ/KA)),其中basal表示无激动剂存在时的响应,Em表示检测系统的最大响应,KA表示激动剂的平衡解离常数,[A]表示激动剂浓度,τ是激动剂偶联效率(或效能)的指标,n是将激动剂占据与响应联系起来的转导函数斜率。在分析中,每条通路上所有激动剂曲线族均以全局方式拟合到该模型,参数basal、Em和n在所有激动剂之间共享。对于完全激动剂,logKA被约束为零值,而对于部分激动剂,则通过曲线拟合程序直接估计;log(τ/KA)参数作为每种激动剂活性的独特度量进行估计。激动剂偏向因子按先前描述的方法计算。
深圳市富临神通科技有限公司(原“东莞市富临塑胶原料有限公司”)是Intan Technologies中国代理商,采购Headstages、芯片、记录系统、刺激/记录系统、SPI线、芯片等电生理产品请立即联系我们。
编辑 | 富临塑胶
联系电话:13798761514(微信同号)
邮箱:li@fulinsujiao.com
公司地址:深圳市光明区艾维普思大厦808
临床1期核酸药物
2026-03-24
·知乎专栏
胆碱酯酶抑制剂是目前针对阿尔茨海默病的一线对症治疗药物,但存在疗效低和不良反应的问题。M1毒蕈碱乙酰胆碱受体代表了一个潜在的替代治疗靶点;然而,针对这一G蛋白偶联受体的药物研发项目此前均告失败,主要原因在于胆碱能不良反应。利用新型化学遗传学和磷酸化缺陷型G蛋白偏向性小鼠模型,并结合一系列探针分子工具,研究人员发现了以往未被重视的、可通过药物干预的M1毒蕈碱乙酰胆碱受体神经过程,包括焦虑样行为和过度活动。通过绘制调控这些反应的上游信号通路,研究人员确定了受体磷酸化依赖性信号传导在驱动临床相关结果以及控制包括“癫痫样”惊厥在内的不良反应中的重要性。研究人员得出结论:能促进受体磷酸化依赖性信号传导的M1毒蕈碱乙酰胆碱受体配体,除了能驱动学习记忆和抗焦虑行为等对治疗阿尔茨海默病有益的反应外,还能预防胆碱能不良反应。阿尔茨海默病是最常见的痴呆形式,在英国影响约85万人,在全球范围内影响5000万人。与许多形式的痴呆一样,阿尔茨海默病伴随一系列症状,不仅包括记忆丧失,还包括焦虑和激越等行为障碍。目前尚未出现能够同时应对阿尔茨海默病多种症状的单一治疗策略。相反,阿尔茨海默病症状治疗的基础策略主要集中于通过抑制乙酰胆碱酯酶(负责分解乙酰胆碱的酶)来恢复胆碱能传递,从而解决认知缺陷。然而,这种治疗的疗效受到严重剂量相关胆碱能不良反应的限制,这主要是由于全身(包括中枢和外周)胆碱能系统的整体上调所致。一个被广泛考虑的替代策略是靶向M1毒蕈碱乙酰胆碱受体,这是响应乙酰胆碱的G蛋白偶联受体家族的五个成员之一,在临床前动物研究中显示出促认知效果。然而,尽管在临床试验中出现了一些积极的疗效迹象,偏向M1/M4的正位激动剂呫诺美林和双位激动剂GSK1034702仍因胆碱能不良反应而失败。另一种旨在通过增加对M1毒蕈碱乙酰胆碱受体的选择性来减少不良反应的策略,是靶向非保守的别构位点以正向调节受体活性,即所谓的正向别构调节剂。尽管已发现多种结构不同的M1毒蕈碱乙酰胆碱受体选择性正向别构调节剂,其中一些分子同样遇到了不良反应的问题,包括颞叶癫痫。这些研究突显了人们对M1选择性正向别构调节剂和正位/双位配体在决定临床有效反应与不良反应方面的微妙药理和结构特性严重缺乏认识。此外,利用偏向信号传导的概念来规避曾阻碍药物开发的毒蕈碱不良反应的潜力,也同样缺乏深入的研究。为了充分发挥M1毒蕈碱乙酰胆碱受体偏向性配体的潜力,有必要解析介导临床相关M1毒蕈碱乙酰胆碱受体神经过程的体内信号通路,并将其与导致不良反应的通路区分开来。研究人员通过构建G蛋白偏向性M1毒蕈碱乙酰胆碱受体来应对这一挑战,具体方法是去除受体磷酸化位点,从而使受体与磷酸化/抑制蛋白依赖性信号传导脱钩。通过将该G蛋白偏向性受体敲入野生型M1毒蕈碱乙酰胆碱受体的基因位点,研究人员能够将神经和外周反应归因于G蛋白依赖性信号传导或受体磷酸化/抑制蛋白依赖性信号传导。研究人员将这一方法与设计师药物专一性激活设计师受体技术的独特应用相结合,该技术在M1毒蕈碱乙酰胆碱受体的跨膜结构域3和5引入突变,产生了一种对天然配体乙酰胆碱无响应、但能被原本惰性的化学配体氯氮平-N-氧化物激活的受体突变体。尽管许多其他研究使用毒蕈碱型设计师受体来探究G蛋白信号传导在神经过程中的作用,但在此,通过将M1-设计师受体的编码序列敲入M1毒蕈碱乙酰胆碱受体基因位点,研究人员在一项旨在明确M1毒蕈碱乙酰胆碱受体作用及其“可成药性”的研究中,用M1-设计师受体替代野生型M1毒蕈碱乙酰胆碱受体进行表达。研究人员预测,这种突变小鼠的表型将模拟M1受体敲除小鼠,因为M1-设计师受体不对乙酰胆碱产生响应。然而,与M1受体敲除小鼠不同的是,在M1-设计师受体小鼠中,由M1毒蕈碱乙酰胆碱受体活性缺失导致的缺陷可通过给予氯氮平-N-氧化物得到纠正。通过这种方式,研究人员认为可以定义那些可能通过药物激活M1毒蕈碱乙酰胆碱受体来实现靶向的生理反应。利用这一方法,结合M1-PD小鼠以及表达磷酸化缺陷型M1-设计师受体的小鼠品系,研究人员在此阐述了,对M1毒蕈碱乙酰胆碱受体的药物靶向不仅影响学习和记忆,还能纠正焦虑相关行为和过度活跃的紊乱,这表明M1毒蕈碱乙酰胆碱受体配体有望治疗与阿尔茨海默病相关的广泛症状。通过绘制M1毒蕈碱乙酰胆碱受体神经反应和不良反应背后的双模信号通路,研究人员进一步得出结论:偏向于M1毒蕈碱乙酰胆碱受体磷酸化/抑制蛋白依赖性信号传导的配体,将在临床相关反应中发挥疗效,同时最大限度地减少包括“癫痫样”惊厥在内的不良反应。 一、结果 01.M1-设计师受体小鼠的构建与先前研究一致,在中国仓鼠卵巢细胞中表达的人源野生型M1毒蕈碱乙酰胆碱受体,在肌醇磷酸积累实验中可被乙酰胆碱强效激活,而对氯氮平-N-氧化物的响应很弱(附表1)。表达M1-设计师受体的中国仓鼠卵巢细胞则表现出乙酰胆碱效价显著降低(5,000倍),同时对氯氮平-N-氧化物呈现纳摩尔级别的响应(附表1)。研究人员通过将M1毒蕈碱乙酰胆碱受体基因座进行靶向改造,经同源重组后用M1-设计师受体的编码序列替换M1毒蕈碱乙酰胆碱受体的编码序列(图1a和附图1a-c),由此获得的小鼠在海马和皮质中转录的M1-设计师受体信使RNA水平与野生型动物(指表达C端带有HA表位标签的野生型M1毒蕈碱乙酰胆碱受体的动物)中M1毒蕈碱乙酰胆碱受体的水平无显著差异(图1b)。此外,对照小鼠皮质中M1-设计师受体蛋白的表达水平与野生型受体相当,但在海马中的表达水平略高(图1c、d和附图2)。M1-设计师受体小鼠表现出正常的繁殖能力和健康指标。 图1 M1-设计师受体敲入小鼠的构建a, M1受体的蛇形图,显示了为构建M1-设计师受体而引入的突变位点。 b, 逆转录定量聚合酶链式反应显示野生型或M1-设计师受体小鼠海马或皮质中M1毒蕈碱乙酰胆碱受体RNA的转录水平。数据表示为与β-肌动蛋白RNA转录的比值(\(n = 3\)只小鼠)。 c, d, 使用HA标签抗体,通过蛋白质印迹分析检测野生型M1-HA、M1-设计师受体和M1敲除小鼠海马(c)或皮质(d)制备的膜溶物中M1毒蕈碱乙酰胆碱受体的表达水平。所示数据为每种基因型各两只小鼠的结果。至少在另外两次独立实验中获得了类似数据(附图2)。Na⁺/K⁺-ATP酶的表达作为上样对照。 e, f, 在卡巴胆碱或氯氮平-N-氧化物刺激下,野生型(e)或M1-设计师受体(f)小鼠皮质膜制备物中[³⁵S]-GTPγS结合的刺激情况。数据表示为均值 ± 标准误,来自三到四次独立实验,每次实验设重复,并以野生型受体的最大响应进行归一化。为了确定工程化小鼠中表达的M1-设计师受体对毒蕈碱配体的敏感性,研究人员在从皮质制备的膜中评估了M1-设计师受体与异源三聚体G蛋白的偶联情况。在这些实验中,氯氮平-N-氧化物在野生型动物的皮质膜中没有显示出显著活性,而毒蕈碱激动剂卡巴胆碱则使[³⁵S]GTPγS结合出现强劲增加(图1e和附表2)。相比之下,在M1-设计师受体小鼠的皮质膜中,氯氮平-N-氧化物引起Gq/11偶联的显著增强,而卡巴胆碱则未能刺激出反应(图1f和附表2)。(原“东莞市富临塑胶原料有限公司”)是 Intan Technologies中国代理商,我们为客户提供Headstages、芯片、记录系统、刺激/记录系统、SPI线等电生理产品。因此,在工程化小鼠中,M1-设计师受体的表达水平与野生型小鼠中的M1毒蕈碱乙酰胆碱受体接近相当,该受体不被天然配体乙酰胆碱激活,但能被氯氮平-N-氧化物激活。02.氯氮平-N-氧化物降低M1-设计师受体小鼠的过度焦虑水平在高架十字迷宫测试中,M1敲除动物表现出焦虑样表型增强,表现为与对照小鼠相比,进入开放臂的次数显著减少(图2a–g)。与对照相比,M1-设计师受体小鼠同样表现出进入开放臂的次数(占总进入次数的比例)减少,表明M1-设计师受体动物模拟了M1敲除小鼠的表型(图2a–g)。M1-设计师受体小鼠表现出的过度焦虑表型,通过在行为测试前30分钟给予氯氮平-N-氧化物(0.3 mg kg⁻¹)得以恢复到正常水平(图2a–g)。研究人员实验室先前的药代动力学研究确定,给予0.3 mg kg⁻¹氯氮平-N-氧化物(腹腔注射)可达到约50 nM的血浆暴露浓度,该浓度足以在体内完全激活病毒表达的毒蕈碱型设计师受体。重要的是,氯氮平-N-氧化物(0.3 mg kg⁻¹,腹腔注射)对野生型小鼠和M1敲除小鼠在高架十字迷宫测试中的表现均无影响(图2a–g)。这些数据表明,M1毒蕈碱乙酰胆碱受体功能缺失导致焦虑样表型,而该表型可通过选择性配体直接激活M1毒蕈碱乙酰胆碱受体来恢复。图2 M1-设计师受体小鼠的表型改变可通过氯氮平-N-氧化物得到纠正a–f, 热图显示野生型(a、d)、M1敲除(b、e)和M1-设计师受体(c、f)小鼠在高架十字迷宫测试中的活动分布,分别在接受溶剂对照(a–c)或0.3 mg kg⁻¹氯氮平-N-氧化物(d–f)(腹腔注射,测试前30分钟)给药后的情况。热图表示每治疗组2–6只小鼠的活动停留时间(蓝色,0秒停留;红色,5秒停留)。g, 野生型、M1敲除或M1-设计师受体小鼠在高架十字迷宫测试前接受溶剂或氯氮平-N-氧化物处理后的平均焦虑水平(以开臂进入次数/闭臂进入次数与总进入次数的比值计算)。数据来自16–26只小鼠,采用双因素方差分析及Sidak多重比较检验进行统计学分析(*P < 0.05,**P < 0.01,***P < 0.001)。h–j, 野生型(h)、M1敲除(i)或M1-设计师受体(j)小鼠在旷场测试中的代表性运动轨迹图。k, 野生型、M1敲除或M1-设计师受体小鼠在旷场测试中10分钟内的平均运动距离。插图:每种基因型小鼠在10分钟内的总运动距离。数据表示为3–5只小鼠的均值。l, 野生型、M1敲除或M1-设计师受体小鼠在Y迷宫测试前30分钟给予溶剂或0.3 mg kg⁻¹氯氮平-N-氧化物处理后,8分钟内的总运动距离。数据表示为均值 ± 标准误,来自4–13只小鼠,采用双因素方差分析及Sidak多重比较检验进行统计学分析(**P < 0.01;***P < 0.001)。03.M1-设计师受体小鼠的过度活跃可通过氯氮平-N-氧化物恢复与先前研究一致,研究人员证实M1敲除小鼠在旷场测试中表现出过度活跃(图2h–k;另见后续图4e、g–k)。由于M1-设计师受体对天然配体乙酰胆碱无响应,表达M1-设计师受体的小鼠同样表现出过度活动表型(图2h–k)。在Y迷宫范式测试中,M1敲除和M1-设计师受体动物也均表现出明显的过度活动(图2l和附图3)。重要的是,给予氯氮平-N-氧化物(0.3 mg kg⁻¹)对野生型或M1敲除小鼠的活动没有显著影响,但能将M1-设计师受体动物的过度活动降低至与溶剂处理的对照动物相似的水平(图2g、l和附图3)。这些数据表明,M1毒蕈碱乙酰胆碱受体不仅参与调节运动行为,而且通过选择性激动该受体亚型来调控运动行为是具有“可成药性”的。图4 M1毒蕈碱乙酰胆碱受体生理反应的双模信号传导解析a–c, 热图显示野生型(a,\(n = 13\))、M1敲除(b,\(n = 4\))或M1-PD(c,\(n = 14\))小鼠在高架十字迷宫开放臂或闭臂中的活动分布(蓝色,0秒停留;红色,5秒停留)。d, 野生型(\(n = 21\))、M1敲除(\(n = 15\))或M1-PD(\(n = 14\))小鼠的焦虑水平(以开臂进入次数/闭臂进入次数与总进入次数的比值计算)。e, 野生型(\(n = 13\))、M1敲除(\(n = 15\))或M1-PD(\(n = 14\))小鼠在高架十字迷宫中的总运动距离。d和e中的数据以均值 ± 标准误表示,采用单因素方差分析与Dunnett多重比较检验进行统计学分析。** *\(P < 0.05\);***\(P < 0.001\)。f, 野生型或M1-PD小鼠在Y迷宫自发交替范式测试中评估空间工作记忆,测试时长为8分钟。数据以均值 ± 标准误表示,采用Student's \(t\)-检验进行分析。*\(P < 0.05\)。g–i, 野生型(g)、M1敲除(h)和M1-PD(i)小鼠在旷场测试中的代表性轨迹图。j, 野生型、M1敲除和M1-PD小鼠在旷场测试中10分钟内的总运动距离。数据以均值 ± 标准误表示,采用单因素方差分析与Dunnett多重比较检验进行统计学分析。*\(P < 0.05\)。k, 野生型、M1敲除和M1-PD小鼠的基础运动活动,通过体内遥测记录评估。数据展示了8只小鼠在24小时内的平均运动活动量 ± 标准误,该期间的总运动活动量通过曲线下面积计算得出。l, 在野生型、M1敲除和M1-PD小鼠中测量给予毛果芸香碱(1 mg kg⁻¹)后的唾液分泌量。数据以均值 ± 标准误表示,\(n = 5–7\)只小鼠。数据分析采用单因素方差分析与Dunnett多重比较检验,与野生型小鼠进行比较。*\(P < 0.05\)。m, M1毒蕈碱乙酰胆碱受体生理反应示意图,展示位于G蛋白依赖性信号下游的反应(唾液分泌和运动行为)与位于磷酸化依赖性信号下游的反应(抗焦虑行为和空间工作记忆)。04.磷酸化缺陷型M1毒蕈碱乙酰胆碱受体小鼠的构建与许多其他G蛋白偶联受体类似,毒蕈碱受体通过两条信号通路发挥作用——即通过异源三聚体G蛋白和受体磷酸化调控通路。后者包括受体与抑制蛋白接头蛋白的偶联,以及激活抑制蛋白依赖性过程(如受体内化)。研究人员构建了一个M1毒蕈碱乙酰胆碱受体突变体,该突变体缺失了质谱鉴定的所有磷酸化位点以及第三细胞内环和C末端尾部的其他潜在位点(共20个丝氨酸→丙氨酸替换;图3a),并在HEK293细胞中表达。该突变受体表现出强劲的Gq/11/钙动员偶联能力(图3b和附表3),同时显示出激动剂介导的抑制蛋白募集受体的能力减弱(图3c和附表3)以及受体内化缺陷(图3d和附图4)。这些结果与磷酸化缺陷型受体具有“G蛋白偏向性”的观点一致。图3 G蛋白偏向性M1-PD敲入小鼠的构建a, M1受体的蛇形图,显示了为构建M1-PD受体而引入的突变位点。 b, 在瞬时转染小鼠M1野生型或M1-PD毒蕈碱乙酰胆碱受体的HEK细胞中,刺激诱导的细胞内Ca²⁺释放情况。 c, 使用表达M1野生型或M1-PD受体的PathHunter HEK细胞测定β-抑制蛋白向M1毒蕈碱乙酰胆碱受体的募集情况。图中显示了对完全毒蕈碱受体激动剂氧化震颤素-M的浓度-响应曲线。数据以均值 ± 标准误表示,来自三次独立实验,并以野生型M1毒蕈碱乙酰胆碱受体对氧化震颤素-M的最大响应百分比表示。 d, 代表性图像显示,带HA标签的M1野生型或M1-PD在CHO细胞中的定位情况,细胞在固定前用溶剂或100 μM卡巴胆碱刺激1小时,并用4%多聚甲醛固定(×63物镜)。所示数据代表四次独立实验的结果。 e, [³H]-N-甲基东莨菪碱与野生型M1-HA或M1-PD小鼠制备的皮质膜的结合情况(\(n = 4\))。数据以均值 ± 标准误表示。 f, 逆转录定量聚合酶链式反应显示野生型和M1-PD小鼠海马或皮质中M1毒蕈碱乙酰胆碱受体RNA的转录水平。数据表示为与β-肌动蛋白RNA转录的比值(\(n = 3\)只小鼠)。 g, h, 使用HA标签抗体,通过蛋白质印迹分析检测野生型M1-HA、M1-PD和M1敲除小鼠海马(g)或皮质(h)制备的膜溶物中M1毒蕈碱乙酰胆碱受体的表达水平。所示数据为每种基因型各两只小鼠的结果;至少在另外两次独立实验中获得了类似数据(附图6)。Na⁺/K⁺-ATP酶的表达作为上样对照。 i, j, 在卡巴胆碱刺激下,野生型或M1-PD小鼠皮质组织中[³⁵S]-GTPγS与Gq/11的结合刺激(i)或Ins(1,4,5)P₃的积累情况(j)。数据以均值 ± 标准误表示,来自三次独立实验,[³⁵S]-GTPγS结合实验设重复,Ins(1,4,5)P₃积累实验设三重复,并以野生型受体的最大响应进行归一化。研究人员构建了一种表达磷酸化缺陷型小鼠M1毒蕈碱乙酰胆碱受体的受体敲入小鼠(命名为M1-PD;附图5)。通过将该小鼠品系与野生型对照(表达C端带有标签的野生型M1毒蕈碱乙酰胆碱受体)及M1敲除动物进行比较,旨在区分位于受体磷酸化依赖性信号下游的生理反应与由G蛋白激活介导的生理反应。放射性配体结合实验确定,M1-PD小鼠皮质中的毒蕈碱受体表达量与野生型小鼠相当(图3e)。M1-PD小鼠皮质和海马中突变受体的转录水平与对照动物中M1毒蕈碱乙酰胆碱受体的水平无显著差异(图3f),蛋白质印迹结果定量分析显示,M1-PD小鼠海马和皮质中突变型M1毒蕈碱乙酰胆碱受体的表达水平无显著差异(图3g、h和附图6)。最后,在皮质组织提取物中,M1毒蕈碱乙酰胆碱受体与磷酸化缺陷型变体与异源三聚体G蛋白的偶联能力相当(通过[³⁵S]-GTPγS实验和肌醇1,4,5-三磷酸质量实验测定)(图3i、j和附表3)。05.由M1毒蕈碱乙酰胆碱受体磷酸化调控的神经生理学研究人员首先测试了M1-PD小鼠的抗焦虑反应,发现与M1敲除小鼠类似,M1-PD小鼠进入高架十字迷宫开放臂的次数更少(图4a–e)。这表明上述研究中揭示的抗焦虑反应是由受体磷酸化依赖性信号介导的。同样,在测试空间工作记忆的Y迷宫自发交替范式实验中,M1-PD小鼠表现出显著缺陷,表明与M1毒蕈碱乙酰胆碱受体介导的抗焦虑反应一样,这种行为也受到受体磷酸化的正向调控(图4f)。最初在旷场测试中评估运动能力时,M1-PD小鼠并未表现出M1敲除小鼠的过度活跃表型(图4g–j)。相反,在该测试(图4j)和高架十字迷宫(图4e)中,M1-PD小鼠的总运动距离均少于野生型小鼠,表明这些小鼠表现为活动减退。有趣的是,尽管通过遥测技术监测24小时内的活动情况显示M1敲除小鼠过度活跃(与上述数据一致),但在此范式下,M1-PD小鼠的活动水平与野生型小鼠相当(图4k)。从这些运动研究中得出的结论是:与抗焦虑和空间工作记忆反应相反,运动行为的调控不依赖于受体磷酸化,而似乎是以G蛋白依赖性的方式调控的。在某些测试(高架十字迷宫和Y迷宫)中观察到的活动减退反应可能表明,尽管运动行为由G蛋白通路介导,但M1毒蕈碱乙酰胆碱受体介导的运动可能受受体磷酸化调控而发生脱敏,因此当磷酸化位点被移除时,表现为活动减退(图4e、j)。研究人员接下来将这些中枢反应与M1毒蕈碱乙酰胆碱受体介导的唾液分泌进行了比较。如前所述,M1敲除小鼠对毒蕈碱受体激动剂毛果芸香碱的唾液分泌反应减弱,但M1-PD小鼠并未表现出这一现象(图4l),表明该反应同样位于G蛋白依赖性信号的下游(总结于图4m)。06.M1毒蕈碱乙酰胆碱受体磷酸化可减少不良反应为了进一步探究受体磷酸化依赖性信号在M1毒蕈碱乙酰胆碱受体介导反应中的作用,研究人员构建了一个去除磷酸化位点的M1-设计师受体(图5a)。在体外肌醇磷酸实验中,这种磷酸化缺陷型设计师受体被氯氮平-N-氧化物激活的效力和效能与可完全磷酸化的M1-设计师受体版本相当(附图7a和附表1)。与受体磷酸化在驱动磷酸化/抑制蛋白依赖性过程中的作用一致,磷酸化缺陷型M1-设计师受体在激动剂(氯氮平-N-氧化物)介导的受体内化方面存在缺陷(附图7b、c)。图5 M1-DREADD PD小鼠表现出癫痫样惊厥a, M1毒蕈碱乙酰胆碱受体的蛇形图,显示了为构建M1-DREADD PD受体而引入的突变位点。 b, 逆转录定量聚合酶链式反应显示野生型、M1-DREADD或M1-DREADD PD小鼠海马或皮质中M1毒蕈碱乙酰胆碱受体RNA的转录水平。数据表示为与β-肌动蛋白RNA转录的比值(\(n = 3\)只小鼠)。 c, d, 使用HA标签抗体,通过蛋白质印迹分析检测野生型、M1-DREADD、M1-DREADD PD和M1敲除小鼠海马(c)或皮质(d)制备的膜溶物中M1毒蕈碱乙酰胆碱受体的表达水平。所示数据为每种基因型各两只小鼠的结果。至少在另外两次独立实验中获得了类似数据(附图2)。Na⁺/K⁺-ATP酶的表达作为上样对照。 e, f, 在卡巴胆碱或氯氮平-N-氧化物刺激下,M1-DREADD(e)或M1-DREADD PD(f)小鼠皮质膜制备物中[³⁵S]-GTPγS结合刺激情况。数据以均值 ± 标准误表示,来自3–4次独立实验,每次实验设重复,并以野生型受体的最大响应进行归一化。 g, 代表性M1-DREADD PD小鼠在记录基础皮层活动15分钟后给予氯氮平-N-氧化物(0.3 mg kg⁻¹,腹腔注射)后的原始皮层脑电图信号(上图)及其频谱图(下图)。 h, M1-DREADD或M1-DREADD PD小鼠在接受溶剂(5%葡萄糖)或氯氮平-N-氧化物(0.3 mg kg⁻¹)处理后的归一化皮层脑电图功率。数据以均值 ± 标准误表示,来自4–8只小鼠,采用重复测量方差分析进行统计学分析,\(F(3, 3,119) = 5.53\),\(P = 0.029\)。研究人员构建了一种表达磷酸化缺陷型M1-设计师受体以替代M1毒蕈碱乙酰胆碱受体的敲入小鼠,命名为M1-DREADD PD(附图8a、b)。M1-DREADD PD突变体的转录水平与M1-DREADD小鼠中的M1-设计师受体以及野生型小鼠中的M1毒蕈碱乙酰胆碱受体相当(图5b)。可能由于M1-设计师受体不被内源性乙酰胆碱激活,该受体在工程化小鼠海马(而非皮质)中的表达水平略高于野生型受体(附图2)。M1-DREADD PD的表达水平也显著高于野生型受体(图5c、d和附图2)。重要的是,在海马或皮质中,M1-DREADD与M1-DREADD PD受体的表达水平之间无统计学显著差异(附图2)。这一点很重要,因为M1-DREADD作为M1-DREADD PD的对照(在观察M1-DREADD PD的表型时,研究人员不使用野生型小鼠作为对照)。在来源于M1-DREADD和M1-DREADD PD的皮质膜中进行G蛋白偶联实验,结果显示氯氮平-N-氧化物在两种变体中均能强劲刺激Gq/11偶联增加,而卡巴胆碱则无显著增加(图5e、f和附表2)。当根据表达水平进行归一化时,研究人员估计M1-DREADD PD向Gq的信号传导比M1-DREADD高约1.5倍。07.氯氮平-N-氧化物在M1-DREADD PD小鼠中诱发不良反应此处的目的是探究在表达M1-设计师受体的动物中由氯氮平-N-氧化物给药所介导的生理反应,在M1-DREADD PD小鼠中是否同样产生响应。通过这种方式,研究人员可以进一步预测偏向性M1毒蕈碱乙酰胆碱受体选择性配体的影响。然而,对M1-DREADD PD小鼠给予氯氮平-N-氧化物导致了严重的不良反应(附表4),包括中枢和外周反应,如胃肠道紊乱和唾液过度分泌。这些不良反应此前曾与毒蕈碱药物研发项目的失败相关联。然而,最引人注目的是严重的惊厥发作,表现为胆碱能颞叶“癫痫样”惊厥的特征。研究人员进一步利用皮层脑电图记录来监测皮层活动,对这些惊厥进行了表征(图5g、h)。氯氮平-N-氧化物给药后,脑电图功率先降低,随后出现散发的惊厥活动,并在给药后约25–35分钟发展为完全的癫痫样发作。重要的是,相同剂量的氯氮平-N-氧化物(0.3 mg kg⁻¹)给予M1-DREADD小鼠时未产生不良反应(附表4)。08.G蛋白偏向性系统表现出增强的不良反应这些数据预示,与无偏向性配体相比,偏向于G蛋白依赖性信号而非受体磷酸化依赖性信号的毒蕈碱受体配体将表现出更多的不良反应。为了验证这一预测,研究人员对两种特征明确的毒蕈碱配体——毛果芸香碱和GSK1034702的G蛋白/受体磷酸化偏向性进行了分析。这两种配体均能刺激Gq/11依赖性信号,导致肌醇磷酸积累和细胞外信号调节激酶1/2的激活(图6a、b)。同样,毛果芸香碱和GSK1034702均能刺激M1毒蕈碱乙酰胆碱受体第三细胞内环中丝氨酸228的磷酸化,该位点先前被证实对激动剂介导的磷酸化高度敏感(附表5和图6c)。将浓度-响应曲线拟合至激动作用的操作模型后,研究人员得出了每种响应的转导系数。通过将这些系数与天然配体乙酰胆碱的转导系数进行比较,计算了两种配体在G蛋白偶联与受体磷酸化之间的偏向性,以偏向因子表示(附表6和图6d)。该分析确定毛果芸香碱表现出向G蛋白信号的偏向性,而GSK1034702在G蛋白偶联与受体磷酸化通路之间无偏向性(附表6和图6d)。图6 毛果芸香碱表现出G蛋白偏向性,而GSK1034702无偏向性a–c, 在稳定表达小鼠野生型M1毒蕈碱乙酰胆碱受体的CHO细胞中,由乙酰胆碱、毛果芸香碱或GSK1034702刺激的肌醇磷酸积累(a)、ERK1/2磷酸化(b)或M1毒蕈碱乙酰胆碱受体丝氨酸228位点磷酸化(c)。虚线显示作为先前数据集一部分生成的数据,以供比较。d, 在野生型M1受体上,乙酰胆碱、毛果芸香碱和GSK1034702对肌醇磷酸、ERK1/2磷酸化或丝氨酸228磷酸化通路的优先信号偏向(\((\Delta\Delta\log_{10}(\tau/K_A))\))。数据以均值 ± 标准误表示,并以乙酰胆碱作为参考配体。e,f, 给予递增浓度的GSK1034702(e)或毛果芸香碱(f)后,雄性Sprague–Dawley大鼠额叶皮质中肌醇磷酸积累的刺激情况。SKF38393是一种D1多巴胺受体激动剂。数据以均值 ± 标准误表示,来自3–4只大鼠,采用双因素方差分析及Dunnett多重比较检验进行统计学分析。*\(P < 0.05\),**\(P < 0.01\)与溶剂组相比;#\(P < 0.05\),##\(P < 0.01\)与氯化锂组相比。为探究这些配体在体内是否表现出相似特性,研究人员分别给予毛果芸香碱(30 mg kg⁻¹)或GSK1034702(10 mg kg⁻¹),剂量设定为在大鼠中产生相当水平的受体占有率(60%),该占有率通过M1毒蕈碱乙酰胆碱受体正电子发射断层扫描示踪剂LSN317217634测定(附图9)。在此剂量下,毛果芸香碱引起强烈的肌醇磷酸反应,其强度约为GSK1034702观察到的反应的两倍(图6e、f),这支持了以下结论:在脑组织中,毛果芸香碱在刺激G蛋白依赖性信号方面比GSK1034702更有效。随后,研究人员在野生型和M1-PD小鼠中,使用产生相当受体占有率的剂量,检测了这两种配体引发的不良事件。与偏向G蛋白信号的配体会表现出更显著不良反应的观点一致,研究人员观察到毛果芸香碱在野生型和M1-PD小鼠中均诱导了严重的中枢(震颤、抓握和惊厥)和外周(竖毛、流泪和腹泻)不良反应(附表7)。有趣的是,在毛果芸香碱作用下,唾液分泌是M1-PD小鼠中唯一反应低于野生型小鼠的反应(附表7)。尽管这可能提示唾液分泌位于受体磷酸化依赖性信号的下游,但图5l中的数据表明该反应完全由G蛋白介导。与毛果芸香碱相反,无刺激偏向性的GSK1034702在野生型小鼠中未表现出不良反应。仅当在M1-PD小鼠中(即受体呈现G蛋白偏向性的小鼠背景中),该配体才观察到中枢和外周不良反应(附表8)。研究人员接下来利用其实验室先前的发现,即GSK1034702与M1毒蕈碱乙酰胆碱受体结合的双位特性意味着GSK1034702在野生型M1毒蕈碱乙酰胆碱受体和M1-设计师受体上均充当激动剂。这意味着该配体可独特地用作野生型小鼠以及M1-DREADD和M1-DREADD PD小鼠中的激动剂。在M1-DREADD小鼠中无不良反应的剂量(30 mg kg⁻¹)下给予GSK1034702,在M1-DREADD PD小鼠中引起了严重的外周和中枢不良反应(附表9),这一结果与以下观点一致:偏向G蛋白偶联的M1毒蕈碱乙酰胆碱受体信号转导会导致中枢和外周胆碱能不良反应。 二、讨论 在本研究中,我们构建了一系列新型化学遗传学和G蛋白偏向性小鼠模型,这些模型不仅揭示了磷酸化/抑制蛋白依赖性信号在M1毒蕈碱乙酰胆碱受体介导的学习记忆和抗焦虑行为中的重要性,还证实了M1毒蕈碱乙酰胆碱受体的磷酸化状态在减轻胆碱能不良反应方面具有显著作用。这些不良反应,无论是外周SLUDGE效应(流涎、流泪、排尿、排便、胃肠功能紊乱和呕吐)还是中枢不良反应(如惊厥),一直是靶向毒蕈碱受体治疗阿尔茨海默病成功道路上的主要障碍之一。此外,我们的数据表明,在阿尔茨海默病中靶向M1毒蕈碱乙酰胆碱受体,除了促认知的益处外,还可能对相关的行为异常(包括焦虑和过度活跃)产生影响。通过采用设计师受体敲入小鼠,我们不仅模拟了M1毒蕈碱乙酰胆碱受体的药理学激活,还揭示了乙酰胆碱神经信号传导的某些特性,因为在阿尔茨海默病中受损的胆碱能张力,似乎可以通过药物模拟,从而在胆碱能传递受损的背景下实现“正常”的神经元活动和行为反应。在本研究中,我们试图通过绘制G蛋白依赖性与磷酸化/抑制蛋白通路下游的反应(总结于图4m),来探究利用偏向性配体概念引导M1毒蕈碱乙酰胆碱受体信号转向治疗有益通路的潜力。为此,我们通过删除所有潜在的磷酸化受体位点,构建了G蛋白偏向性的M1毒蕈碱乙酰胆碱受体。有趣的是,去除这些磷酸化位点减少但并未消除受体与抑制蛋白的相互作用。这与其他G蛋白偶联受体亚型不同,在后者中去除磷酸化位点会导致受体/抑制蛋白相互作用几乎完全缺失。G蛋白偶联受体可通过与磷酸传感器相互作用的磷酸化依赖性方式以及通过激活传感器相互作用的方式与抑制蛋白结合。结构研究正在揭示这种双模结合的分子基础,特别是不同的磷酸化模式驱动不同抑制蛋白构象和信号输出的作用,这支持了磷酸化条形码的假说。在此,我们报告M1毒蕈碱乙酰胆碱受体与抑制蛋白的相互作用(类似于先前报道的M3毒蕈碱乙酰胆碱受体)仅部分依赖于受体的磷酸化状态,这表明抑制蛋白上的激活传感器在M1毒蕈碱乙酰胆碱受体与抑制蛋白的相互作用中发挥着与磷酸化传感器同等重要的作用。这在解释M1-PD小鼠的表型数据时可能具有重要意义,因为本文中鉴定为依赖于M1毒蕈碱乙酰胆碱受体磷酸化状态的表型,未必是由于受体/抑制蛋白相互作用缺陷所致。相反,受体磷酸化可能以不依赖于抑制蛋白的方式,对神经元中M1毒蕈碱乙酰胆碱受体信号复合物的正确组装/活性至关重要。经典理论认为,G蛋白偶联受体的磷酸化与受体脱敏有关。因此,对M1-PD小鼠中观察到的表型的一种解释可能是受体无法进行磷酸化依赖性脱敏。运动反应的分析表明了这一点。与先前研究一致,我们证实M1敲除小鼠活动过度,表明M1毒蕈碱乙酰胆碱受体在降低运动方面发挥作用。然而,M1-PD小鼠活动减退,这一结果可能意味着磷酸化缺陷型受体本身过度活跃,可能是由于缺乏脱敏所致。然而,当我们评估来自M1-PD小鼠组织的膜和皮质切片中磷酸化缺陷型受体与G蛋白信号的偶联时,仅观察到与G蛋白信号偶联的微弱增加。此外,本文中我们证明位于G蛋白偶联下游的唾液分泌在M1-PD小鼠中未受显著影响。这些数据指向一个有趣的假说:某些由G蛋白信号介导的生理反应(如运动)会被受体磷酸化所脱敏,而另一些(如唾液分泌)则不会。在一系列利用G蛋白偏向性M1-设计师受体突变小鼠的实验中,偏向性信号对胆碱能不良反应的影响得到了惊人的揭示。在该小鼠品系中,我们发现给予氯氮平-N-氧化物(在野生型或M1-DREADD小鼠中不会引起不良反应的浓度)会导致严重的外周和中枢不良反应,包括惊厥。尽管先前研究明确表明不良反应可能与毒蕈碱配体的效能程度(包括正向别构调节剂的协同程度和内在活性)有关,但此前没有任何迹象表明受体磷酸化依赖性过程可能在调节不良反应中发挥作用。根据我们的数据,可以预见,信号偏向于G蛋白偶联而非受体磷酸化通路的配体可能表现出更明显的胆碱能不良反应。毛果芸香碱的情况即是如此,我们在此证明它既具有G蛋白偏向性,又具有显著的致惊厥活性(与先前研究一致)。相比之下,无偏向性的GSK1034702在给予与毛果芸香碱相同受体占有率的剂量时,在野生型小鼠中未引起惊厥。有趣的是,GSK1034702仅在磷酸化缺陷型、G蛋白偏向性小鼠(即M1-PD和M1-DREADD PD)中诱导了胆碱能不良反应。这些数据表明,偏向于受体磷酸化依赖性信号的毒蕈碱配体介导不良反应的倾向较低。我们的研究进一步证实了越来越多的证据,即M1毒蕈碱乙酰胆碱受体可能是以前认为由M2/M3受体介导的外周不良反应的原因。因此,仅仅开发高选择性的M1毒蕈碱乙酰胆碱受体激动剂可能不足以避免胆碱能副作用。这一结论得到了相关报告的支持,即选择性M1毒蕈碱乙酰胆碱受体正向别构调节剂也表现出胆碱能不良反应。这些不良反应似乎与内在活性有关,因为具有高激动剂活性的正向别构调节剂也表现出胆碱能不良反应,而缺乏内在激动剂活性的正向别构调节剂则不诱导胆碱能毒性。目前尚不清楚,且肯定值得进一步研究的一点是,具有高激动剂活性和显著不良反应的正向别构调节剂是否也偏向于G蛋白信号传导。支持这一可能性的数据是,那些诱导不良反应的正向别构调节剂(如PF-06767832和PF-06827443)在纹状体组织中也能刺激强劲的肌醇磷酸信号(表明存在Gq信号传导)。综合这些研究并考虑到我们的发现,我们得出结论:为了在针对阿尔茨海默病多种症状的治疗中最大限度地减少胆碱能副作用,同时实现最大的临床疗效,下一代M1毒蕈碱乙酰胆碱受体配体除了应具有高选择性和经过精细校准的效能外,还应能够驱动受体磷酸化依赖性信号传导。 三、方法 01.动物饲养所有小鼠均以纯合子形式在C57BL/6J背景上繁育。除非另有说明,实验使用8-12周龄的雄性和雌性动物。小鼠可自由摄取标准鼠粮,并在实验前于动物设施中适应至少一周。动物护理遵循国家动物实验指南。所有实验均根据1986年《动物(科学程序)法》在英国内政部颁发的项目许可证下进行。02.M1-PD、M1-DREADD和M1-DREADD PD敲入动物的构建为构建敲入动物,研究人员构建了包含loxP-Stop-loxP盒的载体,该盒位于编码M1-HA、M1-PD、M1-DREADD或M1-DREADD PD的序列上游,并将其插入M1毒蕈碱乙酰胆碱受体基因(Chmr1)的编码外显子(外显子3)中。所有构建体均带有附加在C末端的HA表位序列(YPYDVPDYA)。M1-DREADD是人源化(人源化突变V5A、S254T、K320R、G337A和V413I)M1毒蕈碱乙酰胆碱受体的编码序列,带有Y106C和A196C两个突变(图1a)。M1-PD是小鼠M1毒蕈碱乙酰胆碱受体的编码序列,在第三细胞内环和C末端尾部具有将20个丝氨酸残基替换为丙氨酸的突变(图4a)。M1-DREADD PD是人源化M1-DREADD的编码序列,加上第三细胞内环和C末端尾部的20个丝氨酸→丙氨酸突变(图6a)。包含毒蕈碱受体突变体编码序列的打靶载体随后转染至源自C57BL/6J小鼠的胚胎干细胞中,并筛选新霉素抗性胚胎干细胞。通过聚合酶链式反应和Southern印迹验证同源重组。将重组后的胚胎干细胞克隆注射入囊胚以产生嵌合小鼠。将嵌合小鼠与C57BL/6和表达Cre重组酶的小鼠交配,得以产生杂合子小鼠。杂合子动物通过交配产生纯合子品系。M1敲除小鼠是条件性M1-PD小鼠,其在M1-PD互补DNA上游表达两侧带有loxP位点的转录终止盒。如需获得这些小鼠,请联系通讯作者。M1-HA、M1-PD、M1-DREADD、M1-DREADD PD和M1敲除小鼠的构建由genOway公司完成。需要注意的是,M1敲除品系是使用条件性M1-PD小鼠构建的,该品系未与任何Cre删除品系杂交,因此保留了起始密码子上游的终止盒完整,从而产生不表达受体的小鼠品系(构建体总结见附图4)。03.逆转录定量聚合酶链式反应使用Qiagen脂质组织RNeasy试剂盒,按照制造商说明,从野生型、M1-DREADD、M1-DREADD PD、M1-PD或M1敲除小鼠的海马或皮质中分离RNA。使用Nanodrop定量RNA浓度,每个反应使用1 μg总RNA模板,采用SuperScript III第一链合成SuperMix进行互补DNA合成。将RNA/水(共8 μl)、2 μl逆转录酶和10 μl 2×逆转录反应混合液混合,在25°C孵育10分钟,随后在50°C孵育30分钟,然后在85°C孵育5分钟。随后将样品在冰上冷却。每个反应均在存在和不存在逆转录酶的情况下进行(-逆转录对照)。最后,互补DNA与1 μl(2 U)大肠杆菌RNA酶H在37°C孵育20分钟,随后储存于-20°C直至进行逆转录定量聚合酶链式反应。对于逆转录定量聚合酶链式反应,使用以下M1毒蕈碱乙酰胆碱受体引物(稀释度300:300):F:5′ CAAGTGGCATTCATCGGGATCACC R:5′ GAGAAAGTGCCAATGATGAGATCAGC 每个反应设三复孔。每个反应总体积为25 μl,包含12.5 μl SYBR Green Master Mix、0.75 μl F引物(10 μM储备液)、0.75 μl R引物(10 μM储备液)、10 μl水和1 μl互补DNA(或-逆转录样本)。04.IP1积累实验将人M1野生型或M1-DREADD PD构建体稳定表达于CHO-Flp-In细胞中,在含有10%胎牛血清和1%青霉素/链霉素的Ham's F12培养基中,在潮霉素B选择压力(400 μg ml⁻¹)下培养至T75 cm²培养瓶汇合。收集细胞,以1,000g离心3分钟,然后以1.43 × 10⁶细胞 ml⁻¹的浓度重悬于1×刺激缓冲液(HEPES,10 mM;CaCl₂,1 mM;MgCl₂,0.5 mM;KCl,4.2 mM;NaCl,146 mM;葡萄糖,5.5 mM;LiCl,50 mM;pH 7.4)中。将测试化合物(每孔7 μl)和细胞悬液(每孔7 μl)加入384孔白色ProxiPlate板中。短暂离心后,将板在37°C孵育45分钟。将IP1-d2缀合物和抗IP1穴状化合物Tb缀合物用裂解缓冲液按1:30稀释,每孔各加入3 μl。将板在37°C孵育1小时,使用Envision读板仪检测d2缀合IP1(发射波长665 nm)与Lumi4-Tb穴状化合物缀合抗IP1抗体(发射波长620 nm)之间的荧光共振能量转移。结果由665/620 nm比值计算得出,并以乙酰胆碱刺激的最大响应进行归一化。05.ERK1/2磷酸化使用CisBio Phospho-ERK细胞检测试剂盒评估ERK1/2磷酸化(Thr 202/Tyr 204)的刺激情况。将稳定表达人M1毒蕈碱乙酰胆碱受体的CHO Flp-In细胞培养至汇合单层,实验前进行血清饥饿过夜。用100 μl PBS洗涤细胞,在无血清F12培养基中于37°C孵育。用测试化合物在37°C下刺激细胞5分钟,终体积200 μl。通过快速吸弃培养液并加入50 μl补充有封闭试剂的裂解缓冲液终止刺激。裂解液在室温下轻轻振荡30分钟。随后,将16 μl该裂解液转移至384孔白色ProxiPlate板中,与4 μl预混的抗体溶液在室温下孵育2小时。使用PHERAstar读板仪测定荧光发射(665 nm和620 nm)。06.细胞培养与转染(用于Ca²⁺和β-抑制蛋白募集实验)将小鼠野生型或小鼠磷酸化缺陷型M1毒蕈碱乙酰胆碱受体(位于ProLink载体中)转染至PathHunter HEK293:β-arrestin:EA细胞中,并在抗生素(G418)筛选下培养,以获得稳定表达该受体的细胞池。将HEK293 wt/pdM1:β-arrestin:EA细胞维持在含有L-谷氨酰胺并添加胎牛血清(10%体积比)、潮霉素B(250 μg ml⁻¹)和遗传霉素G418(500 μg ml⁻¹)的DMEM培养基中,于37°C、5% CO₂条件下培养。07.细胞内Ca²⁺测量将PathHunter HEK293 wt/pdM1:β-arrestin细胞以每孔40,000个细胞、90 μl细胞培养基的密度接种于96孔透明底黑色板中,于37°C、5% CO₂下孵育过夜,形成汇合单层。实验当天,向96孔细胞板的每孔中加入30 μl含有0.02% pluronic酸:2.5 mM丙磺舒(1:1)的4× Ca²⁺无洗脱检测试剂盒4,于37°C、5% CO₂下孵育30分钟。随后使用FlexStation 3在时间维度上监测激动剂诱导的细胞内钙离子浓度变化。在加入一系列M1毒蕈碱乙酰胆碱受体激动剂之前,监测基础荧光16秒,之后继续记录荧光变化60秒。对激动剂的响应表示为从基线到峰值的荧光变化。最大荧光值取激动剂加入后初始峰的最高点,最小荧光值取激动剂加入前的背景荧光值。08.抑制蛋白募集实验将PathHunter HEK293 wt/pdM1:β-arrestin细胞以每孔8,000个细胞、20 μl细胞培养基的密度接种于白色透明底384孔ViewPlate板中,于37°C、5% CO₂下孵育过夜,形成汇合单层。实验当天,移除旧培养基,替换为含有0.1%牛血清白蛋白(重量/体积)和20 mM HEPES的HBSS缓冲液(pH 7.4)。用一系列M1毒蕈碱乙酰胆碱受体激动剂刺激细胞2小时(5 μl),之后加入25 μl专有Flash检测试剂,在室温下避光孵育15分钟。使用ClarioStar系统,采用无滤光片的发光程序读取发光信号。09.蛋白质印迹分析0901.膜提取物的制备膜提取物的制备方案参考文献4、51中的膜制备方法。简述如下:将海马组织在含有蛋白酶抑制剂的25 mM磷酸钠缓冲液(pH 7.4)中以3–5 µg振幅进行超声匀浆。样品随后在4°C下以20,000g离心30分钟。将沉淀物与1.2%洋地黄皂苷溶于25 mM磷酸钠和5 mM MgCl₂缓冲液(pH 7.4)中,在4°C下过夜旋转孵育。样品在4°C下以20,000g离心30分钟后,将上清液(膜提取物)转移至新的微量离心管中,储存于-80°C备用。使用Micro BCA蛋白检测试剂盒,按照制造商说明测定蛋白浓度。0902.蛋白质印迹分析样品与含有5% β-巯基乙醇的Laemmli上样缓冲液在37°C下孵育30分钟,然后上样至7.5% SDS-Tris-甘氨酸聚丙烯酰胺凝胶中。样品在±100 V电压下运行,随后转移至硝酸纤维素膜上,膜用含5%脱脂奶粉的TBS-T(含0.1% Tween-20的TBS,pH 7.4)封闭2小时。随后将膜与相应的一抗在4°C下孵育过夜,然后用TBS-T洗涤三次(每次10分钟),并与辣根过氧化物酶偶联的相应二抗(1:5,000)孵育。使用ECL检测系统进行蛋白可视化。0903.[³H]-NMS结合实验将小鼠海马或皮质的膜制备物(每管50 μg)与含有递增浓度(0.1–5 nM)[³H]-NMS的结合缓冲液(HEPES,50 mM;NaCl,110 mM;KCl,5.4 mM;CaCl₂,1.8 mM;MgSO₄,1 mM;葡萄糖,25 mM;蔗糖,58 mM;pH 7.4)在37°C下孵育1小时。通过快速过滤至GF/B玻璃微纤维滤纸上,并用冰冷的0.9% NaCl洗涤三次,将膜结合配体与游离配体分离。通过液体闪烁计数测定膜结合放射性。非特异性结合通过在孵育[³H]-NMS时加入阿托品(1 μM)来确定。0904.[³⁵S]-GTPγS实验人道处死M1野生型、M1-DREADD、M1-PD和M1-DREADD PD小鼠(8–12周龄),然后在冰上解剖皮质组织。将组织悬浮于冰冷的缓冲液A(含0.9%(重量/体积)NaCl、10 mM HEPES、0.2%(重量/体积)EDTA,pH 7.4)中,使用Polytron匀浆器匀浆(4 × 5秒脉冲)。使用Eppendorf 5810R台式离心机在4°C下以200g离心5分钟。收集上清液并按上述方法重新匀浆。随后使用Beckman Coulter Avanti JXN-26离心机(JA-25.25转子)在4°C下以40,000g离心20分钟。弃去上清液,将沉淀重悬于10 ml冰冷的缓冲液B(10 mM HEPES,10 mM EDTA,pH 7.4)中。匀浆沉淀,加入GTP(终浓度1 mM),将悬液在37°C下孵育15分钟。随后在4°C下以40,000g离心20分钟,将沉淀重悬于15 ml冰冷的缓冲液C(10 mM HEPES,0.1 mM EDTA,pH 7.4)中,并如前所述重新匀浆。悬液再次在4°C下以40,000g离心20分钟。将最终沉淀重悬于缓冲液C中,使用Bradford法估算蛋白浓度。然后将匀浆液在最终储存缓冲液中进一步稀释,得到2 mg ml⁻¹的浓度。[³⁵S]-GTPγS结合和Gα亚基免疫沉淀按先前描述进行。具体而言,将M1野生型、M1-DREADD、M1-PD或M1-DREADD PD膜稀释于含有终浓度1 μM GDP的分析缓冲液(HEPES,10 mM;NaCl,100 mM;MgCl₂,10 mM;pH 7.4)中。将膜(75 μg,总分析体积200 μl)加入[³⁵S]-GTPγS(终浓度1 nM)和激动剂(卡巴胆碱或氯氮平-N-氧化物)中,在30°C下孵育5分钟。通过加入1 ml冰冷的分析缓冲液并立即转移至冰浴终止反应。样品离心(20,000g,6分钟,4°C),通过加入50 μl冰冷的裂解缓冲液(100 mM Tris HCl,200 mM NaCl,1 mM EDTA,1.25% Igepal和0.2% SDS,pH 7.4)并在摇床上于4°C孵育1小时,使膜沉淀溶解。完全重溶蛋白后,加入50 μl不含SDS的裂解缓冲液。使用稀释度为1:100的正常兔血清和TE缓冲液(10 mM Tris HCl,10 mM EDTA,pH 8.0)中的3%(重量/体积)蛋白A-琼脂糖珠,在4°C下孵育60分钟,预清除溶解蛋白。通过离心(20,000g,6分钟,4°C)收集蛋白A-琼脂糖珠和不溶物,将100 μl上清液转移至含有Gq特异性抗血清的新管中,在4°C下孵育过夜。向样品中加入蛋白A-琼脂糖珠,涡旋混合,在4°C下旋转90分钟,然后离心(10,000g,1分钟,4°C)。吸去上清液,用冰冷的裂解缓冲液(不含SDS)洗涤蛋白A-琼脂糖珠三次。将回收的珠子与1 ml FloScint-IV闪烁液混合,通过液体闪烁计数测定放射性。0905.Ins(1,4,5)P₃质量实验通过颈椎脱臼法人道处死M1-HA或M1-PD小鼠。暴露大脑并转移至冰冷的平皿中,解剖大脑皮层。使用McIlwain组织切碎机将大脑皮层交叉切割成300 μm × 300 μm的碎块。将所得组织碎块分散于Krebs-Henseleit缓冲液中,通过多次更换缓冲液洗涤,然后在振荡水浴中于37°C振荡60分钟。在此期间,让组织碎块自然沉降,每10分钟更换一次缓冲液。洗涤期结束后,让大脑皮层碎块自然沉降,将25 μl“压积”组织等分试样转移至含有250 μl KHB的平底5 ml管中。每管用O₂/CO₂(95:5)充气,盖上盖子,放回37°C振荡水浴中。加入药物(使总孵育体积达到300 μl),再次用O₂/CO₂(95:5)充气,并按图注所示时间继续孵育。加入等体积冰冷的1 M三氯乙酸终止孵育,管子在冰上提取30分钟。随后离心(2,000g,20分钟,4°C)。回收上清液,使用二氯二氟甲烷/三正辛胺法中和,并如前所述精确测定肌醇1,4,5-三磷酸浓度。通过加入1 M NaOH溶解组织碎块沉淀。使用Lowry法测定每次孵育的蛋白浓度。这使得IP₃质量积累可表示为每毫克蛋白的pmol IP₃。0906.高架十字迷宫小鼠在高架十字迷宫测试室中适应过夜,测试前保持黑暗状态。对于接受溶剂(5%葡萄糖)或氯氮平-N-氧化物(0.3 mg kg⁻¹)处理的小鼠,在测试前30分钟通过腹腔注射给药。高架十字迷宫由四个不透明的臂(50 × 10 cm)组成:两个闭臂(黑色壁,高30 cm)和两个光照较弱的开臂。将小鼠放置于高架十字迷宫的中心,面向闭臂。使用ANY-maze软件追踪小鼠5分钟,并监测在此期间进入闭臂或开臂的次数。焦虑水平计算为开臂进入次数占总进入次数的百分比。每只动物测试后用70%乙醇清洁迷宫。10.旷场在行为测试套件中过夜适应后,使用旷场测试评估一般运动活动。将小鼠放置于透明有机玻璃方形场地(50 × 50 cm)中,使用ANY-maze软件记录10分钟内的活动。11.Y迷宫小鼠在测试前于行为测试套件中过夜适应。对于接受溶剂(5%葡萄糖)或氯氮平-N-氧化物(0.3 mg kg⁻¹)处理的测试,在测试开始前30分钟通过腹腔注射给药。将小鼠放置于Y迷宫(灰色、非反射底板)的中心,该迷宫有三个相同的臂(A、B和C;道宽5 cm,臂长35 cm,臂高10 cm)。使用ANY-maze软件记录活动。自发交替行为通过测量“ABC”序列(任意顺序)的数量占8分钟测试期间总三联体序列的比例来计算。12.体内遥测使用遥测系统在完整的自由活动的野生型、M1-PD和M1敲除小鼠中测量基础运动活动。在异氟烷(1.5–2%)麻醉和卡洛芬(5 mg kg⁻¹,皮下注射)镇痛下,将TA-F10植入式探头植入腹腔。植入期间,将小鼠置于恒温垫上。手术后,小鼠单笼饲养,恢复一周后进行实验。连续三天直接从应答器获取基础运动活动数据,期间不打扰动物。在家笼中记录运动活动。接收器通过MX2矩阵直接连接到PC的单个计算机端口,从而可以测定所有参数。每60秒收集一次数据,使用Ponemah采集系统进行数据收集和初步处理。13.生物节律分析将Ponemah采集系统收集的遥测实验数据按10分钟序列分组,计算平均值用于进一步分析。使用ChronosFit程序进行分析,采用傅里叶分析和逐步回归技术。14.脑电图记录1401.手术与记录动物(M1-HA野生型、M1-DREADD、M1-DREADD PD)使用异氟烷(1–1.5%)麻醉,并置于立体定位框架中。使用电动推剪剃除动物头部毛发,并用乙醇(70%)和聚维酮碘清洁。在切口部位皮下注射利多卡因(2%,0.1–0.3 mg),并皮下注射卡洛芬(5 mg kg⁻¹)以提供术后镇痛。整个手术过程中使用加热垫维持动物体温在37°C。在颅骨上固定五颗骨钉,两颗位于额部区域(前囟后+1.5 mm,前囟中侧±1 mm)用作额叶皮层脑电图电极,两颗位于顶部区域(前囟后−2 mm,前囟中侧±2 mm)用作顶叶皮层脑电图电极,一颗位于小脑作为接地和参考电极。带有螺钉的电极用牙科水泥固定作为头部固定装置。头部固定装置手术后,动物至少恢复五天方可进行实验。在五天的适应期内,每天对动物进行抓取操作,将其置于记录室中,并将动物头部与记录线缆连接。适应期后的第二天,将动物放回记录室进行电生理记录。记录过程如下:最初15分钟用于记录基础皮层活动。随后给小鼠腹腔注射溶剂(5%葡萄糖)或氯氮平-N-氧化物(0.3 mg kg⁻¹),并继续记录脑电图45分钟。在整个实验过程中,小鼠可自由活动,并监测其是否发生惊厥。动物用异氟烷(1–1.5%)麻醉后,置于立体定位框架中,然后使用反馈温度控制器维持体温在37°C。在切口部位皮下注射利多卡因(2%,0.1 ml),并在背部皮下注射卡洛芬(5 mg kg⁻¹)。切口后,暴露并清洁颅骨。植入四颗骨钉用于皮层脑电图记录。另一颗螺钉植入小脑上方作为接地和参考电极。所有螺钉与连接器相连,并用牙科水泥覆盖。动物至少恢复五天。电生理记录流程详见其他文献。简述如下:将动物置于开放箱中,通过连接16通道放大器板(RHD2132,Intan Technologies)和接口线缆进行记录。信号相对于小脑骨钉进行放大,并以1,000 Hz的频率数字化(RHD2132和RHD 2000,Intan Technologies)。每次记录包括15分钟基线记录、腹腔注射(氯氮平-N-氧化物0.3 mg kg⁻¹或溶剂)以及至少45分钟的再次记录。所有离线分析使用MATLAB(版本R2018b)进行。由于所有四个脑电图通道提供的信号质量相似,仅使用右侧额叶区域的信号。为计算频谱图,应用多窗口谱估计方法()。为评估信号功率,每1秒计算一次均方根值,并以dB为单位缩放。将缩放后的信号经1/300 Hz低通滤波后,相对于基线(前5分钟信号的平均值)进行归一化,以便在不同实验之间进行比较。1402.唾液分泌测量通过腹腔注射氯胺酮(100 mg kg⁻¹)/美托咪定(0.25 mg kg⁻¹)麻醉小鼠。之后,给小鼠注射毛果芸香碱(1 mg ml⁻¹,腹腔注射),并在35分钟内每5分钟记录一次GF/B滤纸上的唾液分泌量(以毫克计)。1403.受体内化的免疫细胞化学将稳定表达HA标签(C末端)小鼠M1野生型、小鼠M1-PD、人源化M1-DREADD或人源化M1-DREADD PD的CHO细胞培养24小时,在涂有0.01%多聚-d-赖氨酸的13 mm玻璃盖玻片上达到60–80%汇合度。用100 µM卡巴胆碱(野生型和M1-PD)或氯氮平-N-氧化物(M1-DREADD或M1-DREADD PD)刺激细胞1小时,用4%多聚甲醛(在TBS缓冲液中)固定,并用含0.1% Triton X-100的TBS缓冲液中的2%牛血清白蛋白进行封闭和透化处理。与抗EEA1多克隆抗体在4°C下孵育过夜,与抗HA抗体在室温下孵育2小时。用TBS缓冲液洗涤三次后,与AlexaFluor 594抗兔和AlexaFluor 488抗兔二抗在室温下孵育2小时,然后用TBS快速洗涤三次。将带有染色细胞的盖玻片用含DAPI的VECTASHIELD HardSet抗淬灭封片剂封片于载玻片上。使用LSM 880共聚焦激光扫描显微镜获取数据。15.大鼠肌醇磷酸积累1501.样本收集大鼠(Sprague–Dawley,250–275 g)脑样本(额叶皮质)在动物接受以下处理后收集:水溶剂或LiCl(100 mg kg⁻¹,皮下注射),30分钟后给予水溶剂或东莨菪碱(1 mg kg⁻¹,皮下注射)或SKF38393(20 mg kg⁻¹,皮下注射),给药体积为1 ml kg⁻¹。LiCl给药后一小时,给大鼠注射毛果芸香碱(10、30或100 mg kg⁻¹,皮下注射)或GSK1034702(3、10或30 mg kg⁻¹,腹腔注射)。两小时后处死大鼠,在干冰上收集额叶皮质。样品储存于-70°C,用于肌醇磷酸的液相色谱-串联质谱分析。1502.质谱分析样品用探头超声破碎仪在8级强度下匀浆10秒,匀浆液为5倍体积的含0.1%甲酸和50%甲醇的50%乙腈。然后样品以13,000g离心12分钟。将100 μl上清液转移至200 μl蒸馏水中,取20 μl等分试样进样至液相色谱/质谱仪。使用肌醇2-单磷酸双(环己基铵)盐在50%乙腈+50%甲醇+0.1%甲酸中制备标准品(10、30、100、300和1,000 ng g⁻¹或ml⁻¹)。肌醇磷酸分析使用Agilent 6410系列三重四极杆液相色谱-串联质谱仪,配备MassHunter数据分析软件,安装电喷雾离子源,以负离子模式运行。通过监测肌醇磷酸的前体离子(质荷比为259)并靶向其产物离子(质荷比设为78.9)进行检测。色谱分离采用Zorbax RX-SIL高效液相色谱柱,流动相为含3%乙腈的水溶液,总体含0.1%甲酸,流速为0.7 ml min⁻¹。每次进样后均可见到与标准品保留时间和预期分子量相符的清晰色谱峰。根据这些峰面积对分析物进行定量。16.体内受体占有率1601.活体阶段雄性Sprague–Dawley大鼠(每个剂量组n=4)购自Harlan,体重在200至300 g之间。给予毛果芸香碱或GSK1034702,剂量为0.03、0.1、0.3、1、3和10 mg kg⁻¹,以生成剂量反应曲线。动物仅接受溶剂或受试化合物,给药体积为10 ml kg⁻¹。在剂量反应研究中,大鼠在溶剂或化合物给药30分钟后,通过尾静脉静脉注射非标记示踪剂LSN317217634,剂量为10 mg kg⁻¹,给药体积为大鼠0.5 ml kg⁻¹、小鼠5 ml kg⁻¹。示踪剂给药20分钟后,通过颈椎脱臼处死动物。取出大脑并进行解剖。额叶皮质和小脑用于示踪剂测量,剩余大脑和血浆用于化合物暴露分析。受体占有率被认为是在示踪剂给药时间点t测定的。研究在Covance Alnwick或Greenfield进行。1602.组织制备与示踪剂分析称量额叶皮质和小脑样本,置于冰上的锥形离心管中。每管加入四倍体积(重量/体积)含0.1%甲酸的乙腈。然后使用超声探头匀浆样品,并使用台式离心机以14,000 r.p.m.离心20分钟。取50 μl上清液加入96孔板中的150 μl无菌水中进行稀释,用于液相色谱-串联质谱分析。使用API 4000质谱仪进行LSN3172176的分析。色谱分离采用Agilent Zorbax Eclipse XDB-C18柱,流动相为梯度洗脱的15–90%乙腈水溶液,总体含0.1%甲酸。通过监测前体离子到产物离子的跃迁(质荷比386.3 → 128.0)来完成LSN3172176的检测。通过向未经处理的大鼠或小鼠脑组织样本中加入已知量的示踪剂,并按上述方法处理,制备标准品。1603.受体占有率测定受体占有率采用比值法计算。测量每个皮质和小脑样本中的示踪剂水平。对每只动物生成皮质水平(总结合)与小脑水平(非特异性结合)的比值。溶剂处理组的比值代表0%占有率,比值为1(即皮质结合等于小脑结合)代表100%占有率。毛果芸香碱和GSK1034702预处理组的比值在溶剂处理组比值(0%占有率)和1(100%占有率)之间进行线性插值,以确定M1受体占有率的百分比。对于毛果芸香碱和GSK1034702的剂量反应,使用GraphPad Prism 6.0版将曲线拟合为四参数逻辑函数,底部和顶部分别固定在0%和100%,并由软件计算达到50%受体占有率的剂量。数值以均值±标准误表示。17.数据分析功能性浓度-响应曲线根据四参数逻辑方程进行拟合(以确定最小和最大渐近线、对数半最大有效浓度及斜率;GraphPad Prism 6)。为评估激动剂偏向性,采用修正形式的激动作用操作模型对相同的浓度-响应曲线进行分析,重新构建后直接得出转导比(log(τ/KA)),其中basal表示无激动剂存在时的响应,Em表示检测系统的最大响应,KA表示激动剂的平衡解离常数,[A]表示激动剂浓度,τ是激动剂偶联效率(或效能)的指标,n是将激动剂占据与响应联系起来的转导函数斜率。在分析中,每条通路上所有激动剂曲线族均以全局方式拟合到该模型,参数basal、Em和n在所有激动剂之间共享。对于完全激动剂,logKA被约束为零值,而对于部分激动剂,则通过曲线拟合程序直接估计;log(τ/KA)参数作为每种激动剂活性的独特度量进行估计。激动剂偏向因子按先前描述的方法计算。
临床1期核酸药物
2026-03-24
·百度百家
胆碱酯酶抑制剂是目前针对阿尔茨海默病的一线对症治疗药物,但存在疗效低和不良反应的问题。M1毒蕈碱乙酰胆碱受体代表了一个潜在的替代治疗靶点;然而,针对这一G蛋白偶联受体的药物研发项目此前均告失败,主要原因在于胆碱能不良反应。利用新型化学遗传学和磷酸化缺陷型G蛋白偏向性小鼠模型,并结合一系列探针分子工具,研究人员发现了以往未被重视的、可通过药物干预的M1毒蕈碱乙酰胆碱受体神经过程,包括焦虑样行为和过度活动。通过绘制调控这些反应的上游信号通路,研究人员确定了受体磷酸化依赖性信号传导在驱动临床相关结果以及控制包括“癫痫样”惊厥在内的不良反应中的重要性。研究人员得出结论:能促进受体磷酸化依赖性信号传导的M1毒蕈碱乙酰胆碱受体配体,除了能驱动学习记忆和抗焦虑行为等对治疗阿尔茨海默病有益的反应外,还能预防胆碱能不良反应。
阿尔茨海默病是最常见的痴呆形式,在英国影响约85万人,在全球范围内影响5000万人。与许多形式的痴呆一样,阿尔茨海默病伴随一系列症状,不仅包括记忆丧失,还包括焦虑和激越等行为障碍。目前尚未出现能够同时应对阿尔茨海默病多种症状的单一治疗策略。相反,阿尔茨海默病症状治疗的基础策略主要集中于通过抑制乙酰胆碱酯酶(负责分解乙酰胆碱的酶)来恢复胆碱能传递,从而解决认知缺陷。然而,这种治疗的疗效受到严重剂量相关胆碱能不良反应的限制,这主要是由于全身(包括中枢和外周)胆碱能系统的整体上调所致。
一个被广泛考虑的替代策略是靶向M1毒蕈碱乙酰胆碱受体,这是响应乙酰胆碱的G蛋白偶联受体家族的五个成员之一,在临床前动物研究中显示出促认知效果。然而,尽管在临床试验中出现了一些积极的疗效迹象,偏向M1/M4的正位激动剂呫诺美林和双位激动剂GSK1034702仍因胆碱能不良反应而失败。另一种旨在通过增加对M1毒蕈碱乙酰胆碱受体的选择性来减少不良反应的策略,是靶向非保守的别构位点以正向调节受体活性,即所谓的正向别构调节剂。尽管已发现多种结构不同的M1毒蕈碱乙酰胆碱受体选择性正向别构调节剂,其中一些分子同样遇到了不良反应的问题,包括颞叶癫痫。这些研究突显了人们对M1选择性正向别构调节剂和正位/双位配体在决定临床有效反应与不良反应方面的微妙药理和结构特性严重缺乏认识。
此外,利用偏向信号传导的概念来规避曾阻碍药物开发的毒蕈碱不良反应的潜力,也同样缺乏深入的研究。为了充分发挥M1毒蕈碱乙酰胆碱受体偏向性配体的潜力,有必要解析介导临床相关M1毒蕈碱乙酰胆碱受体神经过程的体内信号通路,并将其与导致不良反应的通路区分开来。研究人员通过构建G蛋白偏向性M1毒蕈碱乙酰胆碱受体来应对这一挑战,具体方法是去除受体磷酸化位点,从而使受体与磷酸化/抑制蛋白依赖性信号传导脱钩。通过将该G蛋白偏向性受体敲入野生型M1毒蕈碱乙酰胆碱受体的基因位点,研究人员能够将神经和外周反应归因于G蛋白依赖性信号传导或受体磷酸化/抑制蛋白依赖性信号传导。
研究人员将这一方法与设计师药物专一性激活设计师受体技术的独特应用相结合,该技术在M1毒蕈碱乙酰胆碱受体的跨膜结构域3和5引入突变,产生了一种对天然配体乙酰胆碱无响应、但能被原本惰性的化学配体氯氮平-N-氧化物激活的受体突变体。尽管许多其他研究使用毒蕈碱型设计师受体来探究G蛋白信号传导在神经过程中的作用,但在此,通过将M1-设计师受体的编码序列敲入M1毒蕈碱乙酰胆碱受体基因位点,研究人员在一项旨在明确M1毒蕈碱乙酰胆碱受体作用及其“可成药性”的研究中,用M1-设计师受体替代野生型M1毒蕈碱乙酰胆碱受体进行表达。研究人员预测,这种突变小鼠的表型将模拟M1受体敲除小鼠,因为M1-设计师受体不对乙酰胆碱产生响应。然而,与M1受体敲除小鼠不同的是,在M1-设计师受体小鼠中,由M1毒蕈碱乙酰胆碱受体活性缺失导致的缺陷可通过给予氯氮平-N-氧化物得到纠正。通过这种方式,研究人员认为可以定义那些可能通过药物激活M1毒蕈碱乙酰胆碱受体来实现靶向的生理反应。利用这一方法,结合M1-PD小鼠以及表达磷酸化缺陷型M1-设计师受体的小鼠品系,研究人员在此阐述了,对M1毒蕈碱乙酰胆碱受体的药物靶向不仅影响学习和记忆,还能纠正焦虑相关行为和过度活跃的紊乱,这表明M1毒蕈碱乙酰胆碱受体配体有望治疗与阿尔茨海默病相关的广泛症状。通过绘制M1毒蕈碱乙酰胆碱受体神经反应和不良反应背后的双模信号通路,研究人员进一步得出结论:偏向于M1毒蕈碱乙酰胆碱受体磷酸化/抑制蛋白依赖性信号传导的配体,将在临床相关反应中发挥疗效,同时最大限度地减少包括“癫痫样”惊厥在内的不良反应。
一、结果
01.M1-设计师受体小鼠的构建
与先前研究一致,在中国仓鼠卵巢细胞中表达的人源野生型M1毒蕈碱乙酰胆碱受体,在肌醇磷酸积累实验中可被乙酰胆碱强效激活,而对氯氮平-N-氧化物的响应很弱(附表1)。表达M1-设计师受体的中国仓鼠卵巢细胞则表现出乙酰胆碱效价显著降低(5,000倍),同时对氯氮平-N-氧化物呈现纳摩尔级别的响应(附表1)。研究人员通过将M1毒蕈碱乙酰胆碱受体基因座进行靶向改造,经同源重组后用M1-设计师受体的编码序列替换M1毒蕈碱乙酰胆碱受体的编码序列(图1a和附图1a-c),由此获得的小鼠在海马和皮质中转录的M1-设计师受体信使RNA水平与野生型动物(指表达C端带有HA表位标签的野生型M1毒蕈碱乙酰胆碱受体的动物)中M1毒蕈碱乙酰胆碱受体的水平无显著差异(图1b)。此外,对照小鼠皮质中M1-设计师受体蛋白的表达水平与野生型受体相当,但在海马中的表达水平略高(图1c、d和附图2)。M1-设计师受体小鼠表现出正常的繁殖能力和健康指标。
图1 M1-设计师受体敲入小鼠的构建
a, M1受体的蛇形图,显示了为构建M1-设计师受体而引入的突变位点。
b, 逆转录定量聚合酶链式反应显示野生型或M1-设计师受体小鼠海马或皮质中M1毒蕈碱乙酰胆碱受体RNA的转录水平。数据表示为与β-肌动蛋白RNA转录的比值(\(n = 3\)只小鼠)。
c, d, 使用HA标签抗体,通过蛋白质印迹分析检测野生型M1-HA、M1-设计师受体和M1敲除小鼠海马(c)或皮质(d)制备的膜溶物中M1毒蕈碱乙酰胆碱受体的表达水平。所示数据为每种基因型各两只小鼠的结果。至少在另外两次独立实验中获得了类似数据(附图2)。Na⁺/K⁺-ATP酶的表达作为上样对照。
e, f, 在卡巴胆碱或氯氮平-N-氧化物刺激下,野生型(e)或M1-设计师受体(f)小鼠皮质膜制备物中[³⁵S]-GTPγS结合的刺激情况。数据表示为均值 ± 标准误,来自三到四次独立实验,每次实验设重复,并以野生型受体的最大响应进行归一化。
为了确定工程化小鼠中表达的M1-设计师受体对毒蕈碱配体的敏感性,研究人员在从皮质制备的膜中评估了M1-设计师受体与异源三聚体G蛋白的偶联情况。在这些实验中,氯氮平-N-氧化物在野生型动物的皮质膜中没有显示出显著活性,而毒蕈碱激动剂卡巴胆碱则使[³⁵S]GTPγS结合出现强劲增加(图1e和附表2)。相比之下,在M1-设计师受体小鼠的皮质膜中,氯氮平-N-氧化物引起Gq/11偶联的显著增强,而卡巴胆碱则未能刺激出反应(图1f和附表2)。
深圳市富临神通科技有限公司(原“东莞市富临塑胶原料有限公司”)是 Intan Technologies中国代理商,我们为客户提供Headstages、芯片、记录系统、刺激/记录系统、SPI线等电生理产品。
因此,在工程化小鼠中,M1-设计师受体的表达水平与野生型小鼠中的M1毒蕈碱乙酰胆碱受体接近相当,该受体不被天然配体乙酰胆碱激活,但能被氯氮平-N-氧化物激活。
02.氯氮平-N-氧化物降低M1-设计师受体小鼠的过度焦虑水平
在高架十字迷宫测试中,M1敲除动物表现出焦虑样表型增强,表现为与对照小鼠相比,进入开放臂的次数显著减少(图2a–g)。与对照相比,M1-设计师受体小鼠同样表现出进入开放臂的次数(占总进入次数的比例)减少,表明M1-设计师受体动物模拟了M1敲除小鼠的表型(图2a–g)。M1-设计师受体小鼠表现出的过度焦虑表型,通过在行为测试前30分钟给予氯氮平-N-氧化物(0.3 mg kg⁻¹)得以恢复到正常水平(图2a–g)。研究人员实验室先前的药代动力学研究确定,给予0.3 mg kg⁻¹氯氮平-N-氧化物(腹腔注射)可达到约50 nM的血浆暴露浓度,该浓度足以在体内完全激活病毒表达的毒蕈碱型设计师受体。重要的是,氯氮平-N-氧化物(0.3 mg kg⁻¹,腹腔注射)对野生型小鼠和M1敲除小鼠在高架十字迷宫测试中的表现均无影响(图2a–g)。这些数据表明,M1毒蕈碱乙酰胆碱受体功能缺失导致焦虑样表型,而该表型可通过选择性配体直接激活M1毒蕈碱乙酰胆碱受体来恢复。
图2 M1-设计师受体小鼠的表型改变可通过氯氮平-N-氧化物得到纠正
a–f, 热图显示野生型(a、d)、M1敲除(b、e)和M1-设计师受体(c、f)小鼠在高架十字迷宫测试中的活动分布,分别在接受溶剂对照(a–c)或0.3 mg kg⁻¹氯氮平-N-氧化物(d–f)(腹腔注射,测试前30分钟)给药后的情况。热图表示每治疗组2–6只小鼠的活动停留时间(蓝色,0秒停留;红色,5秒停留)。
g, 野生型、M1敲除或M1-设计师受体小鼠在高架十字迷宫测试前接受溶剂或氯氮平-N-氧化物处理后的平均焦虑水平(以开臂进入次数/闭臂进入次数与总进入次数的比值计算)。数据来自16–26只小鼠,采用双因素方差分析及Sidak多重比较检验进行统计学分析(*P < 0.05,**P < 0.01,***P < 0.001)。
h–j, 野生型(h)、M1敲除(i)或M1-设计师受体(j)小鼠在旷场测试中的代表性运动轨迹图。
k, 野生型、M1敲除或M1-设计师受体小鼠在旷场测试中10分钟内的平均运动距离。插图:每种基因型小鼠在10分钟内的总运动距离。数据表示为3–5只小鼠的均值。
l, 野生型、M1敲除或M1-设计师受体小鼠在Y迷宫测试前30分钟给予溶剂或0.3 mg kg⁻¹氯氮平-N-氧化物处理后,8分钟内的总运动距离。数据表示为均值 ± 标准误,来自4–13只小鼠,采用双因素方差分析及Sidak多重比较检验进行统计学分析(**P < 0.01;***P < 0.001)。
03.M1-设计师受体小鼠的过度活跃可通过氯氮平-N-氧化物恢复
与先前研究一致,研究人员证实M1敲除小鼠在旷场测试中表现出过度活跃(图2h–k;另见后续图4e、g–k)。由于M1-设计师受体对天然配体乙酰胆碱无响应,表达M1-设计师受体的小鼠同样表现出过度活动表型(图2h–k)。在Y迷宫范式测试中,M1敲除和M1-设计师受体动物也均表现出明显的过度活动(图2l和附图3)。重要的是,给予氯氮平-N-氧化物(0.3 mg kg⁻¹)对野生型或M1敲除小鼠的活动没有显著影响,但能将M1-设计师受体动物的过度活动降低至与溶剂处理的对照动物相似的水平(图2g、l和附图3)。这些数据表明,M1毒蕈碱乙酰胆碱受体不仅参与调节运动行为,而且通过选择性激动该受体亚型来调控运动行为是具有“可成药性”的。
图4 M1毒蕈碱乙酰胆碱受体生理反应的双模信号传导解析
a–c, 热图显示野生型(a,\(n = 13\))、M1敲除(b,\(n = 4\))或M1-PD(c,\(n = 14\))小鼠在高架十字迷宫开放臂或闭臂中的活动分布(蓝色,0秒停留;红色,5秒停留)。
d, 野生型(\(n = 21\))、M1敲除(\(n = 15\))或M1-PD(\(n = 14\))小鼠的焦虑水平(以开臂进入次数/闭臂进入次数与总进入次数的比值计算)。
e, 野生型(\(n = 13\))、M1敲除(\(n = 15\))或M1-PD(\(n = 14\))小鼠在高架十字迷宫中的总运动距离。
d和e中的数据以均值 ± 标准误表示,采用单因素方差分析与Dunnett多重比较检验进行统计学分析。** *\(P < 0.05\);***\(P < 0.001\)。
f, 野生型或M1-PD小鼠在Y迷宫自发交替范式测试中评估空间工作记忆,测试时长为8分钟。数据以均值 ± 标准误表示,采用Student's \(t\)-检验进行分析。*\(P < 0.05\)。
g–i, 野生型(g)、M1敲除(h)和M1-PD(i)小鼠在旷场测试中的代表性轨迹图。
j, 野生型、M1敲除和M1-PD小鼠在旷场测试中10分钟内的总运动距离。数据以均值 ± 标准误表示,采用单因素方差分析与Dunnett多重比较检验进行统计学分析。*\(P < 0.05\)。
k, 野生型、M1敲除和M1-PD小鼠的基础运动活动,通过体内遥测记录评估。数据展示了8只小鼠在24小时内的平均运动活动量 ± 标准误,该期间的总运动活动量通过曲线下面积计算得出。
l, 在野生型、M1敲除和M1-PD小鼠中测量给予毛果芸香碱(1 mg kg⁻¹)后的唾液分泌量。数据以均值 ± 标准误表示,\(n = 5–7\)只小鼠。数据分析采用单因素方差分析与Dunnett多重比较检验,与野生型小鼠进行比较。*\(P < 0.05\)。
m, M1毒蕈碱乙酰胆碱受体生理反应示意图,展示位于G蛋白依赖性信号下游的反应(唾液分泌和运动行为)与位于磷酸化依赖性信号下游的反应(抗焦虑行为和空间工作记忆)。
04.磷酸化缺陷型M1毒蕈碱乙酰胆碱受体小鼠的构建
与许多其他G蛋白偶联受体类似,毒蕈碱受体通过两条信号通路发挥作用——即通过异源三聚体G蛋白和受体磷酸化调控通路。后者包括受体与抑制蛋白接头蛋白的偶联,以及激活抑制蛋白依赖性过程(如受体内化)。研究人员构建了一个M1毒蕈碱乙酰胆碱受体突变体,该突变体缺失了质谱鉴定的所有磷酸化位点以及第三细胞内环和C末端尾部的其他潜在位点(共20个丝氨酸→丙氨酸替换;图3a),并在HEK293细胞中表达。该突变受体表现出强劲的Gq/11/钙动员偶联能力(图3b和附表3),同时显示出激动剂介导的抑制蛋白募集受体的能力减弱(图3c和附表3)以及受体内化缺陷(图3d和附图4)。这些结果与磷酸化缺陷型受体具有“G蛋白偏向性”的观点一致。
图3 G蛋白偏向性M1-PD敲入小鼠的构建
a, M1受体的蛇形图,显示了为构建M1-PD受体而引入的突变位点。
b, 在瞬时转染小鼠M1野生型或M1-PD毒蕈碱乙酰胆碱受体的HEK细胞中,刺激诱导的细胞内Ca²⁺释放情况。
c, 使用表达M1野生型或M1-PD受体的PathHunter HEK细胞测定β-抑制蛋白向M1毒蕈碱乙酰胆碱受体的募集情况。图中显示了对完全毒蕈碱受体激动剂氧化震颤素-M的浓度-响应曲线。数据以均值 ± 标准误表示,来自三次独立实验,并以野生型M1毒蕈碱乙酰胆碱受体对氧化震颤素-M的最大响应百分比表示。
d, 代表性图像显示,带HA标签的M1野生型或M1-PD在CHO细胞中的定位情况,细胞在固定前用溶剂或100 μM卡巴胆碱刺激1小时,并用4%多聚甲醛固定(×63物镜)。所示数据代表四次独立实验的结果。
e, [³H]-N-甲基东莨菪碱与野生型M1-HA或M1-PD小鼠制备的皮质膜的结合情况(\(n = 4\))。数据以均值 ± 标准误表示。
f, 逆转录定量聚合酶链式反应显示野生型和M1-PD小鼠海马或皮质中M1毒蕈碱乙酰胆碱受体RNA的转录水平。数据表示为与β-肌动蛋白RNA转录的比值(\(n = 3\)只小鼠)。
g, h, 使用HA标签抗体,通过蛋白质印迹分析检测野生型M1-HA、M1-PD和M1敲除小鼠海马(g)或皮质(h)制备的膜溶物中M1毒蕈碱乙酰胆碱受体的表达水平。所示数据为每种基因型各两只小鼠的结果;至少在另外两次独立实验中获得了类似数据(附图6)。Na⁺/K⁺-ATP酶的表达作为上样对照。
i, j, 在卡巴胆碱刺激下,野生型或M1-PD小鼠皮质组织中[³⁵S]-GTPγS与Gq/11的结合刺激(i)或Ins(1,4,5)P₃的积累情况(j)。数据以均值 ± 标准误表示,来自三次独立实验,[³⁵S]-GTPγS结合实验设重复,Ins(1,4,5)P₃积累实验设三重复,并以野生型受体的最大响应进行归一化。
研究人员构建了一种表达磷酸化缺陷型小鼠M1毒蕈碱乙酰胆碱受体的受体敲入小鼠(命名为M1-PD;附图5)。通过将该小鼠品系与野生型对照(表达C端带有标签的野生型M1毒蕈碱乙酰胆碱受体)及M1敲除动物进行比较,旨在区分位于受体磷酸化依赖性信号下游的生理反应与由G蛋白激活介导的生理反应。放射性配体结合实验确定,M1-PD小鼠皮质中的毒蕈碱受体表达量与野生型小鼠相当(图3e)。M1-PD小鼠皮质和海马中突变受体的转录水平与对照动物中M1毒蕈碱乙酰胆碱受体的水平无显著差异(图3f),蛋白质印迹结果定量分析显示,M1-PD小鼠海马和皮质中突变型M1毒蕈碱乙酰胆碱受体的表达水平无显著差异(图3g、h和附图6)。最后,在皮质组织提取物中,M1毒蕈碱乙酰胆碱受体与磷酸化缺陷型变体与异源三聚体G蛋白的偶联能力相当(通过[³⁵S]-GTPγS实验和肌醇1,4,5-三磷酸质量实验测定)(图3i、j和附表3)。
05.由M1毒蕈碱乙酰胆碱受体磷酸化调控的神经生理学
研究人员首先测试了M1-PD小鼠的抗焦虑反应,发现与M1敲除小鼠类似,M1-PD小鼠进入高架十字迷宫开放臂的次数更少(图4a–e)。这表明上述研究中揭示的抗焦虑反应是由受体磷酸化依赖性信号介导的。同样,在测试空间工作记忆的Y迷宫自发交替范式实验中,M1-PD小鼠表现出显著缺陷,表明与M1毒蕈碱乙酰胆碱受体介导的抗焦虑反应一样,这种行为也受到受体磷酸化的正向调控(图4f)。
最初在旷场测试中评估运动能力时,M1-PD小鼠并未表现出M1敲除小鼠的过度活跃表型(图4g–j)。相反,在该测试(图4j)和高架十字迷宫(图4e)中,M1-PD小鼠的总运动距离均少于野生型小鼠,表明这些小鼠表现为活动减退。有趣的是,尽管通过遥测技术监测24小时内的活动情况显示M1敲除小鼠过度活跃(与上述数据一致),但在此范式下,M1-PD小鼠的活动水平与野生型小鼠相当(图4k)。从这些运动研究中得出的结论是:与抗焦虑和空间工作记忆反应相反,运动行为的调控不依赖于受体磷酸化,而似乎是以G蛋白依赖性的方式调控的。在某些测试(高架十字迷宫和Y迷宫)中观察到的活动减退反应可能表明,尽管运动行为由G蛋白通路介导,但M1毒蕈碱乙酰胆碱受体介导的运动可能受受体磷酸化调控而发生脱敏,因此当磷酸化位点被移除时,表现为活动减退(图4e、j)。
研究人员接下来将这些中枢反应与M1毒蕈碱乙酰胆碱受体介导的唾液分泌进行了比较。如前所述,M1敲除小鼠对毒蕈碱受体激动剂毛果芸香碱的唾液分泌反应减弱,但M1-PD小鼠并未表现出这一现象(图4l),表明该反应同样位于G蛋白依赖性信号的下游(总结于图4m)。
06.M1毒蕈碱乙酰胆碱受体磷酸化可减少不良反应
为了进一步探究受体磷酸化依赖性信号在M1毒蕈碱乙酰胆碱受体介导反应中的作用,研究人员构建了一个去除磷酸化位点的M1-设计师受体(图5a)。在体外肌醇磷酸实验中,这种磷酸化缺陷型设计师受体被氯氮平-N-氧化物激活的效力和效能与可完全磷酸化的M1-设计师受体版本相当(附图7a和附表1)。与受体磷酸化在驱动磷酸化/抑制蛋白依赖性过程中的作用一致,磷酸化缺陷型M1-设计师受体在激动剂(氯氮平-N-氧化物)介导的受体内化方面存在缺陷(附图7b、c)。
图5 M1-DREADD PD小鼠表现出癫痫样惊厥
a, M1毒蕈碱乙酰胆碱受体的蛇形图,显示了为构建M1-DREADD PD受体而引入的突变位点。
b, 逆转录定量聚合酶链式反应显示野生型、M1-DREADD或M1-DREADD PD小鼠海马或皮质中M1毒蕈碱乙酰胆碱受体RNA的转录水平。数据表示为与β-肌动蛋白RNA转录的比值(\(n = 3\)只小鼠)。
c, d, 使用HA标签抗体,通过蛋白质印迹分析检测野生型、M1-DREADD、M1-DREADD PD和M1敲除小鼠海马(c)或皮质(d)制备的膜溶物中M1毒蕈碱乙酰胆碱受体的表达水平。所示数据为每种基因型各两只小鼠的结果。至少在另外两次独立实验中获得了类似数据(附图2)。Na⁺/K⁺-ATP酶的表达作为上样对照。
e, f, 在卡巴胆碱或氯氮平-N-氧化物刺激下,M1-DREADD(e)或M1-DREADD PD(f)小鼠皮质膜制备物中[³⁵S]-GTPγS结合刺激情况。数据以均值 ± 标准误表示,来自3–4次独立实验,每次实验设重复,并以野生型受体的最大响应进行归一化。
g, 代表性M1-DREADD PD小鼠在记录基础皮层活动15分钟后给予氯氮平-N-氧化物(0.3 mg kg⁻¹,腹腔注射)后的原始皮层脑电图信号(上图)及其频谱图(下图)。
h, M1-DREADD或M1-DREADD PD小鼠在接受溶剂(5%葡萄糖)或氯氮平-N-氧化物(0.3 mg kg⁻¹)处理后的归一化皮层脑电图功率。数据以均值 ± 标准误表示,来自4–8只小鼠,采用重复测量方差分析进行统计学分析,\(F(3, 3,119) = 5.53\),\(P = 0.029\)。
研究人员构建了一种表达磷酸化缺陷型M1-设计师受体以替代M1毒蕈碱乙酰胆碱受体的敲入小鼠,命名为M1-DREADD PD(附图8a、b)。M1-DREADD PD突变体的转录水平与M1-DREADD小鼠中的M1-设计师受体以及野生型小鼠中的M1毒蕈碱乙酰胆碱受体相当(图5b)。
可能由于M1-设计师受体不被内源性乙酰胆碱激活,该受体在工程化小鼠海马(而非皮质)中的表达水平略高于野生型受体(附图2)。M1-DREADD PD的表达水平也显著高于野生型受体(图5c、d和附图2)。重要的是,在海马或皮质中,M1-DREADD与M1-DREADD PD受体的表达水平之间无统计学显著差异(附图2)。这一点很重要,因为M1-DREADD作为M1-DREADD PD的对照(在观察M1-DREADD PD的表型时,研究人员不使用野生型小鼠作为对照)。
在来源于M1-DREADD和M1-DREADD PD的皮质膜中进行G蛋白偶联实验,结果显示氯氮平-N-氧化物在两种变体中均能强劲刺激Gq/11偶联增加,而卡巴胆碱则无显著增加(图5e、f和附表2)。当根据表达水平进行归一化时,研究人员估计M1-DREADD PD向Gq的信号传导比M1-DREADD高约1.5倍。
07.氯氮平-N-氧化物在M1-DREADD PD小鼠中诱发不良反应
此处的目的是探究在表达M1-设计师受体的动物中由氯氮平-N-氧化物给药所介导的生理反应,在M1-DREADD PD小鼠中是否同样产生响应。通过这种方式,研究人员可以进一步预测偏向性M1毒蕈碱乙酰胆碱受体选择性配体的影响。然而,对M1-DREADD PD小鼠给予氯氮平-N-氧化物导致了严重的不良反应(附表4),包括中枢和外周反应,如胃肠道紊乱和唾液过度分泌。这些不良反应此前曾与毒蕈碱药物研发项目的失败相关联。然而,最引人注目的是严重的惊厥发作,表现为胆碱能颞叶“癫痫样”惊厥的特征。研究人员进一步利用皮层脑电图记录来监测皮层活动,对这些惊厥进行了表征(图5g、h)。氯氮平-N-氧化物给药后,脑电图功率先降低,随后出现散发的惊厥活动,并在给药后约25–35分钟发展为完全的癫痫样发作。重要的是,相同剂量的氯氮平-N-氧化物(0.3 mg kg⁻¹)给予M1-DREADD小鼠时未产生不良反应(附表4)。
08.G蛋白偏向性系统表现出增强的不良反应
这些数据预示,与无偏向性配体相比,偏向于G蛋白依赖性信号而非受体磷酸化依赖性信号的毒蕈碱受体配体将表现出更多的不良反应。为了验证这一预测,研究人员对两种特征明确的毒蕈碱配体——毛果芸香碱和GSK1034702的G蛋白/受体磷酸化偏向性进行了分析。这两种配体均能刺激Gq/11依赖性信号,导致肌醇磷酸积累和细胞外信号调节激酶1/2的激活(图6a、b)。同样,毛果芸香碱和GSK1034702均能刺激M1毒蕈碱乙酰胆碱受体第三细胞内环中丝氨酸228的磷酸化,该位点先前被证实对激动剂介导的磷酸化高度敏感(附表5和图6c)。将浓度-响应曲线拟合至激动作用的操作模型后,研究人员得出了每种响应的转导系数。通过将这些系数与天然配体乙酰胆碱的转导系数进行比较,计算了两种配体在G蛋白偶联与受体磷酸化之间的偏向性,以偏向因子表示(附表6和图6d)。该分析确定毛果芸香碱表现出向G蛋白信号的偏向性,而GSK1034702在G蛋白偶联与受体磷酸化通路之间无偏向性(附表6和图6d)。
图6 毛果芸香碱表现出G蛋白偏向性,而GSK1034702无偏向性
a–c, 在稳定表达小鼠野生型M1毒蕈碱乙酰胆碱受体的CHO细胞中,由乙酰胆碱、毛果芸香碱或GSK1034702刺激的肌醇磷酸积累(a)、ERK1/2磷酸化(b)或M1毒蕈碱乙酰胆碱受体丝氨酸228位点磷酸化(c)。虚线显示作为先前数据集一部分生成的数据,以供比较。
d, 在野生型M1受体上,乙酰胆碱、毛果芸香碱和GSK1034702对肌醇磷酸、ERK1/2磷酸化或丝氨酸228磷酸化通路的优先信号偏向(\((\Delta\Delta\log_{10}(\tau/K_A))\))。数据以均值 ± 标准误表示,并以乙酰胆碱作为参考配体。
e,f, 给予递增浓度的GSK1034702(e)或毛果芸香碱(f)后,雄性Sprague–Dawley大鼠额叶皮质中肌醇磷酸积累的刺激情况。SKF38393是一种D1多巴胺受体激动剂。数据以均值 ± 标准误表示,来自3–4只大鼠,采用双因素方差分析及Dunnett多重比较检验进行统计学分析。*\(P < 0.05\),**\(P < 0.01\)与溶剂组相比;#\(P < 0.05\),##\(P < 0.01\)与氯化锂组相比。
为探究这些配体在体内是否表现出相似特性,研究人员分别给予毛果芸香碱(30 mg kg⁻¹)或GSK1034702(10 mg kg⁻¹),剂量设定为在大鼠中产生相当水平的受体占有率(60%),该占有率通过M1毒蕈碱乙酰胆碱受体正电子发射断层扫描示踪剂LSN317217634测定(附图9)。在此剂量下,毛果芸香碱引起强烈的肌醇磷酸反应,其强度约为GSK1034702观察到的反应的两倍(图6e、f),这支持了以下结论:在脑组织中,毛果芸香碱在刺激G蛋白依赖性信号方面比GSK1034702更有效。
随后,研究人员在野生型和M1-PD小鼠中,使用产生相当受体占有率的剂量,检测了这两种配体引发的不良事件。与偏向G蛋白信号的配体会表现出更显著不良反应的观点一致,研究人员观察到毛果芸香碱在野生型和M1-PD小鼠中均诱导了严重的中枢(震颤、抓握和惊厥)和外周(竖毛、流泪和腹泻)不良反应(附表7)。有趣的是,在毛果芸香碱作用下,唾液分泌是M1-PD小鼠中唯一反应低于野生型小鼠的反应(附表7)。尽管这可能提示唾液分泌位于受体磷酸化依赖性信号的下游,但图5l中的数据表明该反应完全由G蛋白介导。与毛果芸香碱相反,无刺激偏向性的GSK1034702在野生型小鼠中未表现出不良反应。仅当在M1-PD小鼠中(即受体呈现G蛋白偏向性的小鼠背景中),该配体才观察到中枢和外周不良反应(附表8)。
研究人员接下来利用其实验室先前的发现,即GSK1034702与M1毒蕈碱乙酰胆碱受体结合的双位特性意味着GSK1034702在野生型M1毒蕈碱乙酰胆碱受体和M1-设计师受体上均充当激动剂。这意味着该配体可独特地用作野生型小鼠以及M1-DREADD和M1-DREADD PD小鼠中的激动剂。在M1-DREADD小鼠中无不良反应的剂量(30 mg kg⁻¹)下给予GSK1034702,在M1-DREADD PD小鼠中引起了严重的外周和中枢不良反应(附表9),这一结果与以下观点一致:偏向G蛋白偶联的M1毒蕈碱乙酰胆碱受体信号转导会导致中枢和外周胆碱能不良反应。
二、讨论
在本研究中,我们构建了一系列新型化学遗传学和G蛋白偏向性小鼠模型,这些模型不仅揭示了磷酸化/抑制蛋白依赖性信号在M1毒蕈碱乙酰胆碱受体介导的学习记忆和抗焦虑行为中的重要性,还证实了M1毒蕈碱乙酰胆碱受体的磷酸化状态在减轻胆碱能不良反应方面具有显著作用。这些不良反应,无论是外周SLUDGE效应(流涎、流泪、排尿、排便、胃肠功能紊乱和呕吐)还是中枢不良反应(如惊厥),一直是靶向毒蕈碱受体治疗阿尔茨海默病成功道路上的主要障碍之一。此外,我们的数据表明,在阿尔茨海默病中靶向M1毒蕈碱乙酰胆碱受体,除了促认知的益处外,还可能对相关的行为异常(包括焦虑和过度活跃)产生影响。
通过采用设计师受体敲入小鼠,我们不仅模拟了M1毒蕈碱乙酰胆碱受体的药理学激活,还揭示了乙酰胆碱神经信号传导的某些特性,因为在阿尔茨海默病中受损的胆碱能张力,似乎可以通过药物模拟,从而在胆碱能传递受损的背景下实现“正常”的神经元活动和行为反应。
在本研究中,我们试图通过绘制G蛋白依赖性与磷酸化/抑制蛋白通路下游的反应(总结于图4m),来探究利用偏向性配体概念引导M1毒蕈碱乙酰胆碱受体信号转向治疗有益通路的潜力。为此,我们通过删除所有潜在的磷酸化受体位点,构建了G蛋白偏向性的M1毒蕈碱乙酰胆碱受体。有趣的是,去除这些磷酸化位点减少但并未消除受体与抑制蛋白的相互作用。这与其他G蛋白偶联受体亚型不同,在后者中去除磷酸化位点会导致受体/抑制蛋白相互作用几乎完全缺失。G蛋白偶联受体可通过与磷酸传感器相互作用的磷酸化依赖性方式以及通过激活传感器相互作用的方式与抑制蛋白结合。结构研究正在揭示这种双模结合的分子基础,特别是不同的磷酸化模式驱动不同抑制蛋白构象和信号输出的作用,这支持了磷酸化条形码的假说。在此,我们报告M1毒蕈碱乙酰胆碱受体与抑制蛋白的相互作用(类似于先前报道的M3毒蕈碱乙酰胆碱受体)仅部分依赖于受体的磷酸化状态,这表明抑制蛋白上的激活传感器在M1毒蕈碱乙酰胆碱受体与抑制蛋白的相互作用中发挥着与磷酸化传感器同等重要的作用。这在解释M1-PD小鼠的表型数据时可能具有重要意义,因为本文中鉴定为依赖于M1毒蕈碱乙酰胆碱受体磷酸化状态的表型,未必是由于受体/抑制蛋白相互作用缺陷所致。相反,受体磷酸化可能以不依赖于抑制蛋白的方式,对神经元中M1毒蕈碱乙酰胆碱受体信号复合物的正确组装/活性至关重要。
经典理论认为,G蛋白偶联受体的磷酸化与受体脱敏有关。因此,对M1-PD小鼠中观察到的表型的一种解释可能是受体无法进行磷酸化依赖性脱敏。运动反应的分析表明了这一点。与先前研究一致,我们证实M1敲除小鼠活动过度,表明M1毒蕈碱乙酰胆碱受体在降低运动方面发挥作用。然而,M1-PD小鼠活动减退,这一结果可能意味着磷酸化缺陷型受体本身过度活跃,可能是由于缺乏脱敏所致。
然而,当我们评估来自M1-PD小鼠组织的膜和皮质切片中磷酸化缺陷型受体与G蛋白信号的偶联时,仅观察到与G蛋白信号偶联的微弱增加。此外,本文中我们证明位于G蛋白偶联下游的唾液分泌在M1-PD小鼠中未受显著影响。这些数据指向一个有趣的假说:某些由G蛋白信号介导的生理反应(如运动)会被受体磷酸化所脱敏,而另一些(如唾液分泌)则不会。
在一系列利用G蛋白偏向性M1-设计师受体突变小鼠的实验中,偏向性信号对胆碱能不良反应的影响得到了惊人的揭示。在该小鼠品系中,我们发现给予氯氮平-N-氧化物(在野生型或M1-DREADD小鼠中不会引起不良反应的浓度)会导致严重的外周和中枢不良反应,包括惊厥。尽管先前研究明确表明不良反应可能与毒蕈碱配体的效能程度(包括正向别构调节剂的协同程度和内在活性)有关,但此前没有任何迹象表明受体磷酸化依赖性过程可能在调节不良反应中发挥作用。根据我们的数据,可以预见,信号偏向于G蛋白偶联而非受体磷酸化通路的配体可能表现出更明显的胆碱能不良反应。毛果芸香碱的情况即是如此,我们在此证明它既具有G蛋白偏向性,又具有显著的致惊厥活性(与先前研究一致)。相比之下,无偏向性的GSK1034702在给予与毛果芸香碱相同受体占有率的剂量时,在野生型小鼠中未引起惊厥。有趣的是,GSK1034702仅在磷酸化缺陷型、G蛋白偏向性小鼠(即M1-PD和M1-DREADD PD)中诱导了胆碱能不良反应。这些数据表明,偏向于受体磷酸化依赖性信号的毒蕈碱配体介导不良反应的倾向较低。
我们的研究进一步证实了越来越多的证据,即M1毒蕈碱乙酰胆碱受体可能是以前认为由M2/M3受体介导的外周不良反应的原因。因此,仅仅开发高选择性的M1毒蕈碱乙酰胆碱受体激动剂可能不足以避免胆碱能副作用。这一结论得到了相关报告的支持,即选择性M1毒蕈碱乙酰胆碱受体正向别构调节剂也表现出胆碱能不良反应。这些不良反应似乎与内在活性有关,因为具有高激动剂活性的正向别构调节剂也表现出胆碱能不良反应,而缺乏内在激动剂活性的正向别构调节剂则不诱导胆碱能毒性。目前尚不清楚,且肯定值得进一步研究的一点是,具有高激动剂活性和显著不良反应的正向别构调节剂是否也偏向于G蛋白信号传导。支持这一可能性的数据是,那些诱导不良反应的正向别构调节剂(如PF-06767832和PF-06827443)在纹状体组织中也能刺激强劲的肌醇磷酸信号(表明存在Gq信号传导)。
综合这些研究并考虑到我们的发现,我们得出结论:为了在针对阿尔茨海默病多种症状的治疗中最大限度地减少胆碱能副作用,同时实现最大的临床疗效,下一代M1毒蕈碱乙酰胆碱受体配体除了应具有高选择性和经过精细校准的效能外,还应能够驱动受体磷酸化依赖性信号传导。
三、方法
01.动物饲养
所有小鼠均以纯合子形式在C57BL/6J背景上繁育。除非另有说明,实验使用8-12周龄的雄性和雌性动物。小鼠可自由摄取标准鼠粮,并在实验前于动物设施中适应至少一周。动物护理遵循国家动物实验指南。所有实验均根据1986年《动物(科学程序)法》在英国内政部颁发的项目许可证下进行。
02.M1-PD、M1-DREADD和M1-DREADD PD敲入动物的构建
为构建敲入动物,研究人员构建了包含loxP-Stop-loxP盒的载体,该盒位于编码M1-HA、M1-PD、M1-DREADD或M1-DREADD PD的序列上游,并将其插入M1毒蕈碱乙酰胆碱受体基因(Chmr1)的编码外显子(外显子3)中。所有构建体均带有附加在C末端的HA表位序列(YPYDVPDYA)。M1-DREADD是人源化(人源化突变V5A、S254T、K320R、G337A和V413I)M1毒蕈碱乙酰胆碱受体的编码序列,带有Y106C和A196C两个突变(图1a)。M1-PD是小鼠M1毒蕈碱乙酰胆碱受体的编码序列,在第三细胞内环和C末端尾部具有将20个丝氨酸残基替换为丙氨酸的突变(图4a)。M1-DREADD PD是人源化M1-DREADD的编码序列,加上第三细胞内环和C末端尾部的20个丝氨酸→丙氨酸突变(图6a)。
包含毒蕈碱受体突变体编码序列的打靶载体随后转染至源自C57BL/6J小鼠的胚胎干细胞中,并筛选新霉素抗性胚胎干细胞。通过聚合酶链式反应和Southern印迹验证同源重组。将重组后的胚胎干细胞克隆注射入囊胚以产生嵌合小鼠。将嵌合小鼠与C57BL/6和表达Cre重组酶的小鼠交配,得以产生杂合子小鼠。杂合子动物通过交配产生纯合子品系。M1敲除小鼠是条件性M1-PD小鼠,其在M1-PD互补DNA上游表达两侧带有loxP位点的转录终止盒。如需获得这些小鼠,请联系通讯作者。M1-HA、M1-PD、M1-DREADD、M1-DREADD PD和M1敲除小鼠的构建由genOway公司完成。
需要注意的是,M1敲除品系是使用条件性M1-PD小鼠构建的,该品系未与任何Cre删除品系杂交,因此保留了起始密码子上游的终止盒完整,从而产生不表达受体的小鼠品系(构建体总结见附图4)。
03.逆转录定量聚合酶链式反应
使用Qiagen脂质组织RNeasy试剂盒,按照制造商说明,从野生型、M1-DREADD、M1-DREADD PD、M1-PD或M1敲除小鼠的海马或皮质中分离RNA。使用Nanodrop定量RNA浓度,每个反应使用1 μg总RNA模板,采用SuperScript III第一链合成SuperMix进行互补DNA合成。将RNA/水(共8 μl)、2 μl逆转录酶和10 μl 2×逆转录反应混合液混合,在25°C孵育10分钟,随后在50°C孵育30分钟,然后在85°C孵育5分钟。随后将样品在冰上冷却。每个反应均在存在和不存在逆转录酶的情况下进行(-逆转录对照)。最后,互补DNA与1 μl(2 U)大肠杆菌RNA酶H在37°C孵育20分钟,随后储存于-20°C直至进行逆转录定量聚合酶链式反应。
对于逆转录定量聚合酶链式反应,使用以下M1毒蕈碱乙酰胆碱受体引物(稀释度300:300):
F:5′ CAAGTGGCATTCATCGGGATCACC
R:5′ GAGAAAGTGCCAATGATGAGATCAGC
每个反应设三复孔。每个反应总体积为25 μl,包含12.5 μl SYBR Green Master Mix、0.75 μl F引物(10 μM储备液)、0.75 μl R引物(10 μM储备液)、10 μl水和1 μl互补DNA(或-逆转录样本)。
04.IP1积累实验
将人M1野生型或M1-DREADD PD构建体稳定表达于CHO-Flp-In细胞中,在含有10%胎牛血清和1%青霉素/链霉素的Ham's F12培养基中,在潮霉素B选择压力(400 μg ml⁻¹)下培养至T75 cm²培养瓶汇合。收集细胞,以1,000g离心3分钟,然后以1.43 × 10⁶细胞 ml⁻¹的浓度重悬于1×刺激缓冲液(HEPES,10 mM;CaCl₂,1 mM;MgCl₂,0.5 mM;KCl,4.2 mM;NaCl,146 mM;葡萄糖,5.5 mM;LiCl,50 mM;pH 7.4)中。将测试化合物(每孔7 μl)和细胞悬液(每孔7 μl)加入384孔白色ProxiPlate板中。短暂离心后,将板在37°C孵育45分钟。将IP1-d2缀合物和抗IP1穴状化合物Tb缀合物用裂解缓冲液按1:30稀释,每孔各加入3 μl。将板在37°C孵育1小时,使用Envision读板仪检测d2缀合IP1(发射波长665 nm)与Lumi4-Tb穴状化合物缀合抗IP1抗体(发射波长620 nm)之间的荧光共振能量转移。结果由665/620 nm比值计算得出,并以乙酰胆碱刺激的最大响应进行归一化。
05.ERK1/2磷酸化
使用CisBio Phospho-ERK细胞检测试剂盒评估ERK1/2磷酸化(Thr 202/Tyr 204)的刺激情况。将稳定表达人M1毒蕈碱乙酰胆碱受体的CHO Flp-In细胞培养至汇合单层,实验前进行血清饥饿过夜。用100 μl PBS洗涤细胞,在无血清F12培养基中于37°C孵育。用测试化合物在37°C下刺激细胞5分钟,终体积200 μl。通过快速吸弃培养液并加入50 μl补充有封闭试剂的裂解缓冲液终止刺激。裂解液在室温下轻轻振荡30分钟。随后,将16 μl该裂解液转移至384孔白色ProxiPlate板中,与4 μl预混的抗体溶液在室温下孵育2小时。使用PHERAstar读板仪测定荧光发射(665 nm和620 nm)。
06.细胞培养与转染(用于Ca²⁺和β-抑制蛋白募集实验)
将小鼠野生型或小鼠磷酸化缺陷型M1毒蕈碱乙酰胆碱受体(位于ProLink载体中)转染至PathHunter HEK293:β-arrestin:EA细胞中,并在抗生素(G418)筛选下培养,以获得稳定表达该受体的细胞池。将HEK293 wt/pdM1:β-arrestin:EA细胞维持在含有L-谷氨酰胺并添加胎牛血清(10%体积比)、潮霉素B(250 μg ml⁻¹)和遗传霉素G418(500 μg ml⁻¹)的DMEM培养基中,于37°C、5% CO₂条件下培养。
07.细胞内Ca²⁺测量
将PathHunter HEK293 wt/pdM1:β-arrestin细胞以每孔40,000个细胞、90 μl细胞培养基的密度接种于96孔透明底黑色板中,于37°C、5% CO₂下孵育过夜,形成汇合单层。实验当天,向96孔细胞板的每孔中加入30 μl含有0.02% pluronic酸:2.5 mM丙磺舒(1:1)的4× Ca²⁺无洗脱检测试剂盒4,于37°C、5% CO₂下孵育30分钟。随后使用FlexStation 3在时间维度上监测激动剂诱导的细胞内钙离子浓度变化。在加入一系列M1毒蕈碱乙酰胆碱受体激动剂之前,监测基础荧光16秒,之后继续记录荧光变化60秒。对激动剂的响应表示为从基线到峰值的荧光变化。最大荧光值取激动剂加入后初始峰的最高点,最小荧光值取激动剂加入前的背景荧光值。
08.抑制蛋白募集实验
将PathHunter HEK293 wt/pdM1:β-arrestin细胞以每孔8,000个细胞、20 μl细胞培养基的密度接种于白色透明底384孔ViewPlate板中,于37°C、5% CO₂下孵育过夜,形成汇合单层。实验当天,移除旧培养基,替换为含有0.1%牛血清白蛋白(重量/体积)和20 mM HEPES的HBSS缓冲液(pH 7.4)。用一系列M1毒蕈碱乙酰胆碱受体激动剂刺激细胞2小时(5 μl),之后加入25 μl专有Flash检测试剂,在室温下避光孵育15分钟。使用ClarioStar系统,采用无滤光片的发光程序读取发光信号。
09.蛋白质印迹分析
0901.膜提取物的制备
膜提取物的制备方案参考文献4、51中的膜制备方法。简述如下:将海马组织在含有蛋白酶抑制剂的25 mM磷酸钠缓冲液(pH 7.4)中以3–5 µg振幅进行超声匀浆。样品随后在4°C下以20,000g离心30分钟。将沉淀物与1.2%洋地黄皂苷溶于25 mM磷酸钠和5 mM MgCl₂缓冲液(pH 7.4)中,在4°C下过夜旋转孵育。样品在4°C下以20,000g离心30分钟后,将上清液(膜提取物)转移至新的微量离心管中,储存于-80°C备用。使用Micro BCA蛋白检测试剂盒,按照制造商说明测定蛋白浓度。
0902.蛋白质印迹分析
样品与含有5% β-巯基乙醇的Laemmli上样缓冲液在37°C下孵育30分钟,然后上样至7.5% SDS-Tris-甘氨酸聚丙烯酰胺凝胶中。样品在±100 V电压下运行,随后转移至硝酸纤维素膜上,膜用含5%脱脂奶粉的TBS-T(含0.1% Tween-20的TBS,pH 7.4)封闭2小时。随后将膜与相应的一抗在4°C下孵育过夜,然后用TBS-T洗涤三次(每次10分钟),并与辣根过氧化物酶偶联的相应二抗(1:5,000)孵育。使用ECL检测系统进行蛋白可视化。
0903.[³H]-NMS结合实验
将小鼠海马或皮质的膜制备物(每管50 μg)与含有递增浓度(0.1–5 nM)[³H]-NMS的结合缓冲液(HEPES,50 mM;NaCl,110 mM;KCl,5.4 mM;CaCl₂,1.8 mM;MgSO₄,1 mM;葡萄糖,25 mM;蔗糖,58 mM;pH 7.4)在37°C下孵育1小时。通过快速过滤至GF/B玻璃微纤维滤纸上,并用冰冷的0.9% NaCl洗涤三次,将膜结合配体与游离配体分离。通过液体闪烁计数测定膜结合放射性。非特异性结合通过在孵育[³H]-NMS时加入阿托品(1 μM)来确定。
0904.[³⁵S]-GTPγS实验
人道处死M1野生型、M1-DREADD、M1-PD和M1-DREADD PD小鼠(8–12周龄),然后在冰上解剖皮质组织。将组织悬浮于冰冷的缓冲液A(含0.9%(重量/体积)NaCl、10 mM HEPES、0.2%(重量/体积)EDTA,pH 7.4)中,使用Polytron匀浆器匀浆(4 × 5秒脉冲)。使用Eppendorf 5810R台式离心机在4°C下以200g离心5分钟。收集上清液并按上述方法重新匀浆。随后使用Beckman Coulter Avanti JXN-26离心机(JA-25.25转子)在4°C下以40,000g离心20分钟。弃去上清液,将沉淀重悬于10 ml冰冷的缓冲液B(10 mM HEPES,10 mM EDTA,pH 7.4)中。匀浆沉淀,加入GTP(终浓度1 mM),将悬液在37°C下孵育15分钟。随后在4°C下以40,000g离心20分钟,将沉淀重悬于15 ml冰冷的缓冲液C(10 mM HEPES,0.1 mM EDTA,pH 7.4)中,并如前所述重新匀浆。悬液再次在4°C下以40,000g离心20分钟。将最终沉淀重悬于缓冲液C中,使用Bradford法估算蛋白浓度。然后将匀浆液在最终储存缓冲液中进一步稀释,得到2 mg ml⁻¹的浓度。
[³⁵S]-GTPγS结合和Gα亚基免疫沉淀按先前描述进行。具体而言,将M1野生型、M1-DREADD、M1-PD或M1-DREADD PD膜稀释于含有终浓度1 μM GDP的分析缓冲液(HEPES,10 mM;NaCl,100 mM;MgCl₂,10 mM;pH 7.4)中。将膜(75 μg,总分析体积200 μl)加入[³⁵S]-GTPγS(终浓度1 nM)和激动剂(卡巴胆碱或氯氮平-N-氧化物)中,在30°C下孵育5分钟。通过加入1 ml冰冷的分析缓冲液并立即转移至冰浴终止反应。样品离心(20,000g,6分钟,4°C),通过加入50 μl冰冷的裂解缓冲液(100 mM Tris HCl,200 mM NaCl,1 mM EDTA,1.25% Igepal和0.2% SDS,pH 7.4)并在摇床上于4°C孵育1小时,使膜沉淀溶解。完全重溶蛋白后,加入50 μl不含SDS的裂解缓冲液。使用稀释度为1:100的正常兔血清和TE缓冲液(10 mM Tris HCl,10 mM EDTA,pH 8.0)中的3%(重量/体积)蛋白A-琼脂糖珠,在4°C下孵育60分钟,预清除溶解蛋白。通过离心(20,000g,6分钟,4°C)收集蛋白A-琼脂糖珠和不溶物,将100 μl上清液转移至含有Gq特异性抗血清的新管中,在4°C下孵育过夜。向样品中加入蛋白A-琼脂糖珠,涡旋混合,在4°C下旋转90分钟,然后离心(10,000g,1分钟,4°C)。吸去上清液,用冰冷的裂解缓冲液(不含SDS)洗涤蛋白A-琼脂糖珠三次。将回收的珠子与1 ml FloScint-IV闪烁液混合,通过液体闪烁计数测定放射性。
0905.Ins(1,4,5)P₃质量实验
通过颈椎脱臼法人道处死M1-HA或M1-PD小鼠。暴露大脑并转移至冰冷的平皿中,解剖大脑皮层。使用McIlwain组织切碎机将大脑皮层交叉切割成300 μm × 300 μm的碎块。将所得组织碎块分散于Krebs-Henseleit缓冲液中,通过多次更换缓冲液洗涤,然后在振荡水浴中于37°C振荡60分钟。在此期间,让组织碎块自然沉降,每10分钟更换一次缓冲液。
洗涤期结束后,让大脑皮层碎块自然沉降,将25 μl“压积”组织等分试样转移至含有250 μl KHB的平底5 ml管中。每管用O₂/CO₂(95:5)充气,盖上盖子,放回37°C振荡水浴中。加入药物(使总孵育体积达到300 μl),再次用O₂/CO₂(95:5)充气,并按图注所示时间继续孵育。加入等体积冰冷的1 M三氯乙酸终止孵育,管子在冰上提取30分钟。随后离心(2,000g,20分钟,4°C)。回收上清液,使用二氯二氟甲烷/三正辛胺法中和,并如前所述精确测定肌醇1,4,5-三磷酸浓度。通过加入1 M NaOH溶解组织碎块沉淀。使用Lowry法测定每次孵育的蛋白浓度。这使得IP₃质量积累可表示为每毫克蛋白的pmol IP₃。
0906.高架十字迷宫
小鼠在高架十字迷宫测试室中适应过夜,测试前保持黑暗状态。对于接受溶剂(5%葡萄糖)或氯氮平-N-氧化物(0.3 mg kg⁻¹)处理的小鼠,在测试前30分钟通过腹腔注射给药。
高架十字迷宫由四个不透明的臂(50 × 10 cm)组成:两个闭臂(黑色壁,高30 cm)和两个光照较弱的开臂。将小鼠放置于高架十字迷宫的中心,面向闭臂。使用ANY-maze软件追踪小鼠5分钟,并监测在此期间进入闭臂或开臂的次数。焦虑水平计算为开臂进入次数占总进入次数的百分比。每只动物测试后用70%乙醇清洁迷宫。
10.旷场
在行为测试套件中过夜适应后,使用旷场测试评估一般运动活动。将小鼠放置于透明有机玻璃方形场地(50 × 50 cm)中,使用ANY-maze软件记录10分钟内的活动。
11.Y迷宫
小鼠在测试前于行为测试套件中过夜适应。对于接受溶剂(5%葡萄糖)或氯氮平-N-氧化物(0.3 mg kg⁻¹)处理的测试,在测试开始前30分钟通过腹腔注射给药。将小鼠放置于Y迷宫(灰色、非反射底板)的中心,该迷宫有三个相同的臂(A、B和C;道宽5 cm,臂长35 cm,臂高10 cm)。使用ANY-maze软件记录活动。自发交替行为通过测量“ABC”序列(任意顺序)的数量占8分钟测试期间总三联体序列的比例来计算。
12.体内遥测
使用遥测系统在完整的自由活动的野生型、M1-PD和M1敲除小鼠中测量基础运动活动。在异氟烷(1.5–2%)麻醉和卡洛芬(5 mg kg⁻¹,皮下注射)镇痛下,将TA-F10植入式探头植入腹腔。植入期间,将小鼠置于恒温垫上。手术后,小鼠单笼饲养,恢复一周后进行实验。连续三天直接从应答器获取基础运动活动数据,期间不打扰动物。在家笼中记录运动活动。接收器通过MX2矩阵直接连接到PC的单个计算机端口,从而可以测定所有参数。每60秒收集一次数据,使用Ponemah采集系统进行数据收集和初步处理。
13.生物节律分析
将Ponemah采集系统收集的遥测实验数据按10分钟序列分组,计算平均值用于进一步分析。使用ChronosFit程序进行分析,采用傅里叶分析和逐步回归技术。
14.脑电图记录
1401.手术与记录
动物(M1-HA野生型、M1-DREADD、M1-DREADD PD)使用异氟烷(1–1.5%)麻醉,并置于立体定位框架中。使用电动推剪剃除动物头部毛发,并用乙醇(70%)和聚维酮碘清洁。在切口部位皮下注射利多卡因(2%,0.1–0.3 mg),并皮下注射卡洛芬(5 mg kg⁻¹)以提供术后镇痛。整个手术过程中使用加热垫维持动物体温在37°C。在颅骨上固定五颗骨钉,两颗位于额部区域(前囟后+1.5 mm,前囟中侧±1 mm)用作额叶皮层脑电图电极,两颗位于顶部区域(前囟后−2 mm,前囟中侧±2 mm)用作顶叶皮层脑电图电极,一颗位于小脑作为接地和参考电极。带有螺钉的电极用牙科水泥固定作为头部固定装置。头部固定装置手术后,动物至少恢复五天方可进行实验。在五天的适应期内,每天对动物进行抓取操作,将其置于记录室中,并将动物头部与记录线缆连接。适应期后的第二天,将动物放回记录室进行电生理记录。
记录过程如下:最初15分钟用于记录基础皮层活动。随后给小鼠腹腔注射溶剂(5%葡萄糖)或氯氮平-N-氧化物(0.3 mg kg⁻¹),并继续记录脑电图45分钟。在整个实验过程中,小鼠可自由活动,并监测其是否发生惊厥。
动物用异氟烷(1–1.5%)麻醉后,置于立体定位框架中,然后使用反馈温度控制器维持体温在37°C。在切口部位皮下注射利多卡因(2%,0.1 ml),并在背部皮下注射卡洛芬(5 mg kg⁻¹)。切口后,暴露并清洁颅骨。植入四颗骨钉用于皮层脑电图记录。另一颗螺钉植入小脑上方作为接地和参考电极。所有螺钉与连接器相连,并用牙科水泥覆盖。动物至少恢复五天。
电生理记录流程详见其他文献。简述如下:将动物置于开放箱中,通过连接16通道放大器板(RHD2132,Intan Technologies)和接口线缆进行记录。信号相对于小脑骨钉进行放大,并以1,000 Hz的频率数字化(RHD2132和RHD 2000,Intan Technologies)。每次记录包括15分钟基线记录、腹腔注射(氯氮平-N-氧化物0.3 mg kg⁻¹或溶剂)以及至少45分钟的再次记录。
所有离线分析使用MATLAB(版本R2018b)进行。由于所有四个脑电图通道提供的信号质量相似,仅使用右侧额叶区域的信号。为计算频谱图,应用多窗口谱估计方法(http://chronux.org/)。为评估信号功率,每1秒计算一次均方根值,并以dB为单位缩放。将缩放后的信号经1/300 Hz低通滤波后,相对于基线(前5分钟信号的平均值)进行归一化,以便在不同实验之间进行比较。
1402.唾液分泌测量
通过腹腔注射氯胺酮(100 mg kg⁻¹)/美托咪定(0.25 mg kg⁻¹)麻醉小鼠。之后,给小鼠注射毛果芸香碱(1 mg ml⁻¹,腹腔注射),并在35分钟内每5分钟记录一次GF/B滤纸上的唾液分泌量(以毫克计)。
1403.受体内化的免疫细胞化学
将稳定表达HA标签(C末端)小鼠M1野生型、小鼠M1-PD、人源化M1-DREADD或人源化M1-DREADD PD的CHO细胞培养24小时,在涂有0.01%多聚-d-赖氨酸的13 mm玻璃盖玻片上达到60–80%汇合度。用100 µM卡巴胆碱(野生型和M1-PD)或氯氮平-N-氧化物(M1-DREADD或M1-DREADD PD)刺激细胞1小时,用4%多聚甲醛(在TBS缓冲液中)固定,并用含0.1% Triton X-100的TBS缓冲液中的2%牛血清白蛋白进行封闭和透化处理。与抗EEA1多克隆抗体在4°C下孵育过夜,与抗HA抗体在室温下孵育2小时。用TBS缓冲液洗涤三次后,与AlexaFluor 594抗兔和AlexaFluor 488抗兔二抗在室温下孵育2小时,然后用TBS快速洗涤三次。将带有染色细胞的盖玻片用含DAPI的VECTASHIELD HardSet抗淬灭封片剂封片于载玻片上。使用LSM 880共聚焦激光扫描显微镜获取数据。
15.大鼠肌醇磷酸积累
1501.样本收集
大鼠(Sprague–Dawley,250–275 g)脑样本(额叶皮质)在动物接受以下处理后收集:水溶剂或LiCl(100 mg kg⁻¹,皮下注射),30分钟后给予水溶剂或东莨菪碱(1 mg kg⁻¹,皮下注射)或SKF38393(20 mg kg⁻¹,皮下注射),给药体积为1 ml kg⁻¹。LiCl给药后一小时,给大鼠注射毛果芸香碱(10、30或100 mg kg⁻¹,皮下注射)或GSK1034702(3、10或30 mg kg⁻¹,腹腔注射)。两小时后处死大鼠,在干冰上收集额叶皮质。样品储存于-70°C,用于肌醇磷酸的液相色谱-串联质谱分析。
1502.质谱分析
样品用探头超声破碎仪在8级强度下匀浆10秒,匀浆液为5倍体积的含0.1%甲酸和50%甲醇的50%乙腈。然后样品以13,000g离心12分钟。将100 μl上清液转移至200 μl蒸馏水中,取20 μl等分试样进样至液相色谱/质谱仪。使用肌醇2-单磷酸双(环己基铵)盐在50%乙腈+50%甲醇+0.1%甲酸中制备标准品(10、30、100、300和1,000 ng g⁻¹或ml⁻¹)。肌醇磷酸分析使用Agilent 6410系列三重四极杆液相色谱-串联质谱仪,配备MassHunter数据分析软件,安装电喷雾离子源,以负离子模式运行。通过监测肌醇磷酸的前体离子(质荷比为259)并靶向其产物离子(质荷比设为78.9)进行检测。色谱分离采用Zorbax RX-SIL高效液相色谱柱,流动相为含3%乙腈的水溶液,总体含0.1%甲酸,流速为0.7 ml min⁻¹。每次进样后均可见到与标准品保留时间和预期分子量相符的清晰色谱峰。根据这些峰面积对分析物进行定量。
16.体内受体占有率
1601.活体阶段
雄性Sprague–Dawley大鼠(每个剂量组n=4)购自Harlan,体重在200至300 g之间。给予毛果芸香碱或GSK1034702,剂量为0.03、0.1、0.3、1、3和10 mg kg⁻¹,以生成剂量反应曲线。动物仅接受溶剂或受试化合物,给药体积为10 ml kg⁻¹。在剂量反应研究中,大鼠在溶剂或化合物给药30分钟后,通过尾静脉静脉注射非标记示踪剂LSN317217634,剂量为10 mg kg⁻¹,给药体积为大鼠0.5 ml kg⁻¹、小鼠5 ml kg⁻¹。示踪剂给药20分钟后,通过颈椎脱臼处死动物。取出大脑并进行解剖。额叶皮质和小脑用于示踪剂测量,剩余大脑和血浆用于化合物暴露分析。受体占有率被认为是在示踪剂给药时间点t测定的。研究在Covance Alnwick或Greenfield进行。
1602.组织制备与示踪剂分析
称量额叶皮质和小脑样本,置于冰上的锥形离心管中。每管加入四倍体积(重量/体积)含0.1%甲酸的乙腈。然后使用超声探头匀浆样品,并使用台式离心机以14,000 r.p.m.离心20分钟。取50 μl上清液加入96孔板中的150 μl无菌水中进行稀释,用于液相色谱-串联质谱分析。使用API 4000质谱仪进行LSN3172176的分析。
色谱分离采用Agilent Zorbax Eclipse XDB-C18柱,流动相为梯度洗脱的15–90%乙腈水溶液,总体含0.1%甲酸。通过监测前体离子到产物离子的跃迁(质荷比386.3 → 128.0)来完成LSN3172176的检测。通过向未经处理的大鼠或小鼠脑组织样本中加入已知量的示踪剂,并按上述方法处理,制备标准品。
1603.受体占有率测定
受体占有率采用比值法计算。测量每个皮质和小脑样本中的示踪剂水平。对每只动物生成皮质水平(总结合)与小脑水平(非特异性结合)的比值。溶剂处理组的比值代表0%占有率,比值为1(即皮质结合等于小脑结合)代表100%占有率。毛果芸香碱和GSK1034702预处理组的比值在溶剂处理组比值(0%占有率)和1(100%占有率)之间进行线性插值,以确定M1受体占有率的百分比。对于毛果芸香碱和GSK1034702的剂量反应,使用GraphPad Prism 6.0版将曲线拟合为四参数逻辑函数,底部和顶部分别固定在0%和100%,并由软件计算达到50%受体占有率的剂量。数值以均值±标准误表示。
17.数据分析
功能性浓度-响应曲线根据四参数逻辑方程进行拟合(以确定最小和最大渐近线、对数半最大有效浓度及斜率;GraphPad Prism 6)。为评估激动剂偏向性,采用修正形式的激动作用操作模型对相同的浓度-响应曲线进行分析,重新构建后直接得出转导比(log(τ/KA)),其中basal表示无激动剂存在时的响应,Em表示检测系统的最大响应,KA表示激动剂的平衡解离常数,[A]表示激动剂浓度,τ是激动剂偶联效率(或效能)的指标,n是将激动剂占据与响应联系起来的转导函数斜率。在分析中,每条通路上所有激动剂曲线族均以全局方式拟合到该模型,参数basal、Em和n在所有激动剂之间共享。对于完全激动剂,logKA被约束为零值,而对于部分激动剂,则通过曲线拟合程序直接估计;log(τ/KA)参数作为每种激动剂活性的独特度量进行估计。激动剂偏向因子按先前描述的方法计算。
深圳市富临神通科技有限公司(原“东莞市富临塑胶原料有限公司”)是Intan Technologies中国代理商,采购Headstages、芯片、记录系统、刺激/记录系统、SPI线、芯片等电生理产品请立即联系我们。
100 项与 PF-06767832 相关的药物交易
登录后查看更多信息
研发状态
登录后查看更多信息
临床结果
临床结果
适应症
分期
评价
查看全部结果
| 研究 | 分期 | 人群特征 | 评价人数 | 分组 | 结果 | 评价 | 发布日期 |
|---|
No Data | |||||||
登录后查看更多信息
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
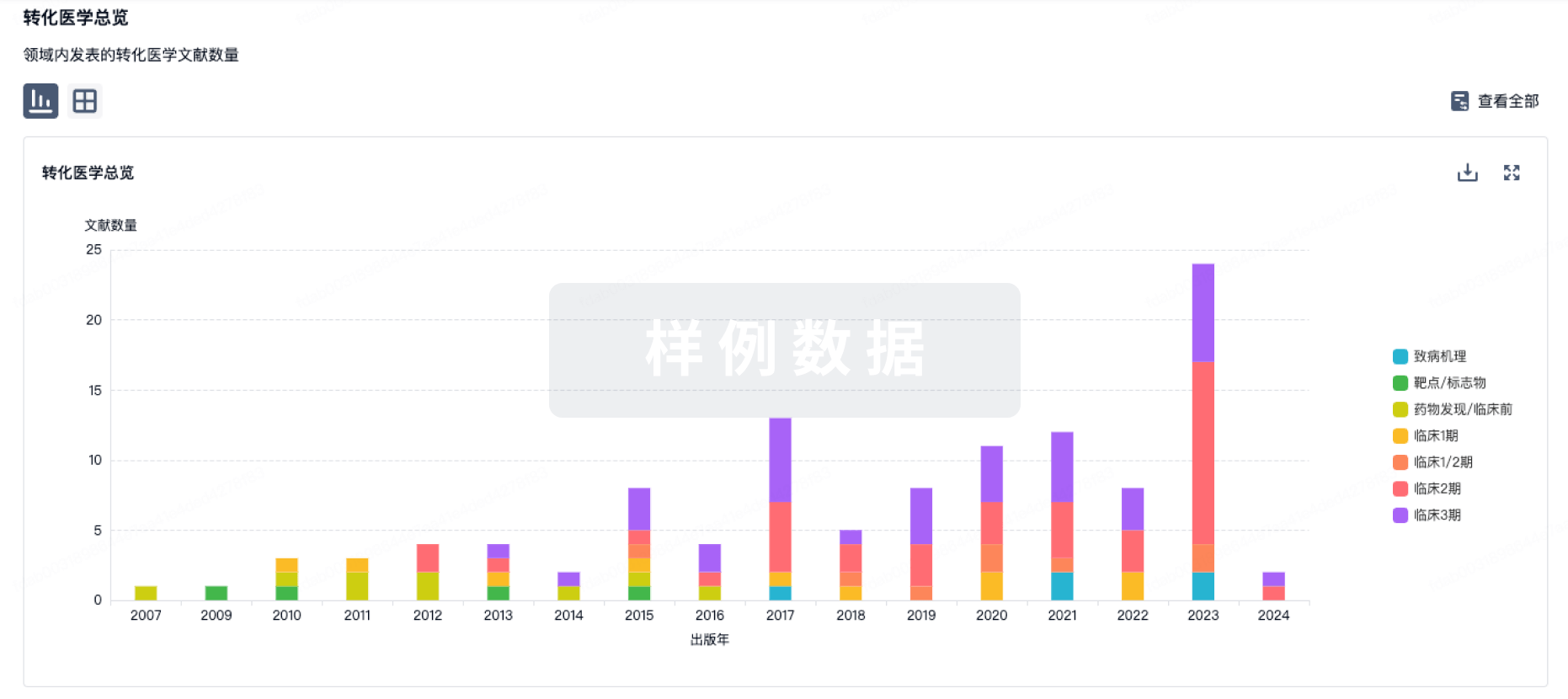
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
核心专利
使用我们的核心专利数据促进您的研究。
登录
或
临床分析
紧跟全球注册中心的最新临床试验。
登录
或
批准
利用最新的监管批准信息加速您的研究。
登录
或
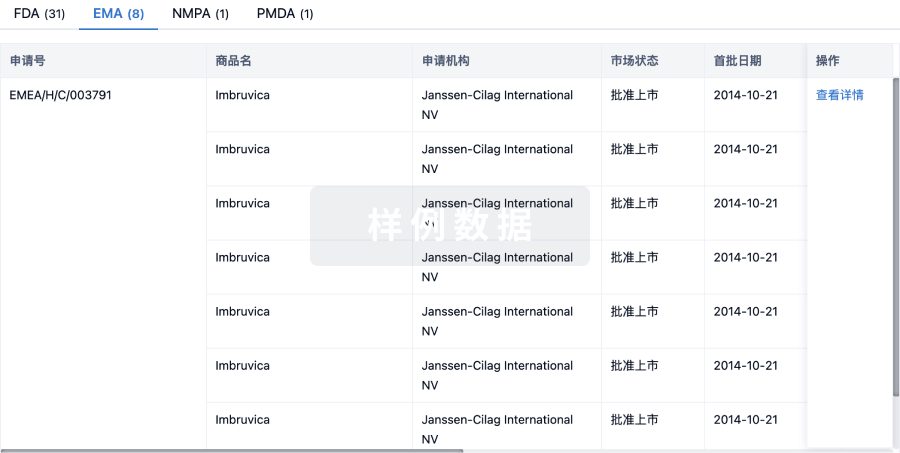
特殊审评
只需点击几下即可了解关键药物信息。
登录
或
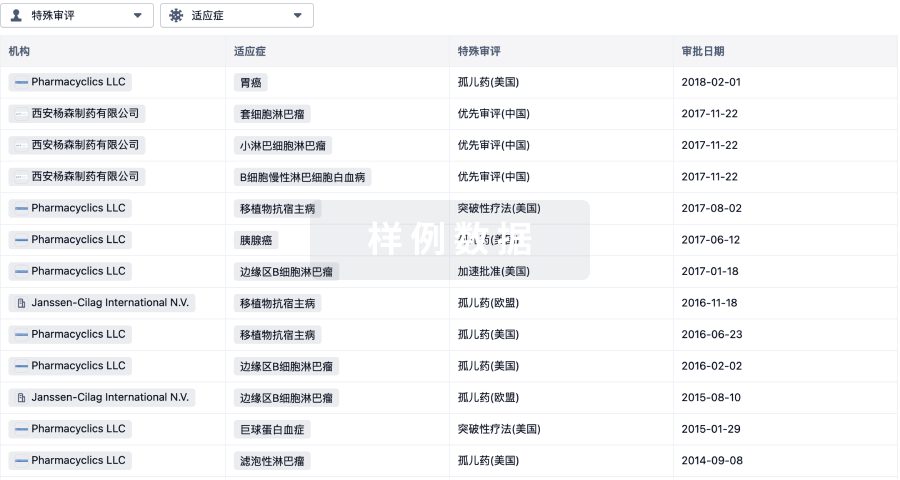
生物医药百科问答
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用