预约演示
更新于:2026-05-17
Shanghai McLean Biochemical Technology Co., Ltd.
更新于:2026-05-17
概览
标签
皮肤和肌肉骨骼疾病
其他疾病
免疫系统疾病
小分子化药

关联
3
项与 上海麦克林生化科技股份有限公司 相关的药物靶点- |
作用机制- |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症- |
最高研发阶段临床前 |
首次获批国家/地区- |
首次获批日期- |
靶点 |
作用机制 XO抑制剂 |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症- |
最高研发阶段临床前 |
首次获批国家/地区- |
首次获批日期- |
靶点- |
作用机制- |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症- |
最高研发阶段临床前 |
首次获批国家/地区- |
首次获批日期- |
100 项与 上海麦克林生化科技股份有限公司 相关的临床结果
登录后查看更多信息
0 项与 上海麦克林生化科技股份有限公司 相关的专利(医药)
登录后查看更多信息
21
项与 上海麦克林生化科技股份有限公司 相关的新闻(医药)2026-05-07
·今日头条
在认知健康消费意识加速觉醒的2026年,市场对脑健康原料的需求正从“泛保养”转向“成分量化”。在大脑认知、记忆修复、神经退行性病变预防等领域,一种源自植物源的超长链单不饱和脂肪酸——神经酸,正迅速从专业健康人群走向大众功能性食品饮料的舞台。
本文将从市场趋势分析、神经酸核心功效及科研支撑、行业法规及原料准入、国内外上市产品分析、神经酸98%规格优势,以及西安锐博生物神经酸原料的综合竞争力等维度展开,帮助功能食品饮料品牌方在脑健康赛道上做出更精准、更前瞻的选品决策。
一、市场趋势分析:神经酸何以成为功能食品饮料领域的新风口?
1. 全球脑健康补充剂市场持续扩容
随着全球人口老龄化进程加快及现代社会中高强度认知负荷工作人群不断扩大,大脑健康补充剂市场正迎来高速增长。据Grand View Research数据,2021年全球大脑健康补充剂市场规模为76.8亿美元,到2030年预计将达到157.4亿美元。另据QYResearch最新统计,2025年全球有机灵芝提取物市场销量达74.18亿美元,预计2032年将增至134.8亿美元,年复合增长率达9.0%,脑健康品类的强劲增速可见一斑。
在细分赛道上,神经酸日益成为脑健康原料中的“关键角色”。全球神经酸市场规模数据:2024年全球神经酸市场规模约2.9亿美元,截至2029年预计增长至4.6亿美元,年复合增长率达到8.0%。随着消费者健康意识的提高和神经酸研究不断深入,市场空间还在持续扩大。与此同时,2026年脑健康膳食补充剂消费趋势报告指出,主动进行系统性脑健康管理的人群中,35岁及以上成人占比超过72%,这意味着中青年人群的认知维护需求正在被加速释放。
2. 从“老龄化应对”走向“全龄段主动健康”
以往神经酸的消费场景集中在老年记忆力减退、阿尔茨海默症预防及神经退行性病变干预领域。现实中,伴随高强度工作压力、快节奏生活方式与不良睡眠习惯,35岁至企业高层、互联网从业者、在校学生群体开始主动关注认知健康管理。据尚普咨询最新调研数据,73%的消费者在过去一年选择了国产健康补充剂产品,“功效导向型”人群占比已达31%,产品是否通过第三方检测和提供真实含量全凭数据成为选购关键。在国人对中草药及植物源天然提取物偏好度持续走高的大背景下,神经酸的消费信心群体迅速扩大。
二、神经酸的核心功效与科学机制
神经酸(Nervonic Acid,又称顺-15-二十四碳烯酸,分子式C24H46O2)是一种超长链单不饱和Omega-9脂肪酸,是大脑神经细胞膜和神经髓鞘的核心构成成分。长期以来,学者将神经酸誉为“神经修复因子”和脑神经发育、连接信号传导的原始材料。
1. 2025年突破性学术研究
2025年10月,国际公认学术期刊ScienceDirect发表题为“Sources and biological functions of nervonic acid: Advances and perspectives”的综述论文,全面阐明了神经酸的多重生物功能。其生物活性与应用涵盖了多个维度:可通过减少氧化应激来改善认知功能;平衡促炎和抗炎因子,以减轻肝脏或结肠等器官的炎症;同时调节肠道菌群。尤为关键的是,神经酸水平波动与精神障碍和代谢性疾病密切相关,已展现出作为疾病早期诊断生物标志物的潜力。
2. 2025年神经酸改性牛奶动物实验重要的临床佐证实证
2025年7月,一篇发表于PubMed的临床试验论文“含神经酸改性牛奶对东莨菪碱诱导的记忆障碍小鼠的改善效果”堪称功能性食品原料领域颇有影响力的研究之一。该研究发现,经过30天含神经酸的改性牛奶干预后,小鼠在Y型迷宫、新物体识别和旷场试验的行为学分析中展现出认知功能的显著改善,说明该改性牛奶有效缓解了东莨菪碱诱导的记忆障碍。
进一步机制研究发现:含神经酸改性牛奶能显著降低海马组织的氧化应激水平,提高超氧化物歧化酶(SOD)和谷胱甘肽过氧化物酶(GSH-Px)活性,降低丙二醛(MDA)水平;同时抑制乙酰胆碱酯酶(AchE)活性调节胆碱能系统,最终重塑肠道菌群并调整厚壁菌门(Firmicutes)/拟杆菌门(Bacteroidetes)的比例。神经酸被证实能增强抗氧化的防御,增加乙酰胆碱(Ach)水平,长期维持神经修复功能,改善记忆。
3. 神经酸如何成为脑健康的“防守球员”
从机理上看,人体摄入量不足会导致神经元细胞膜流动性下降,信息传输效率降低,进而出现记忆力下降、注意力不集中、情绪波动等。由于天然神经酸在人体内合成受限,人体自身不能充分合成,须从外界摄入。因此,神经酸被认定为大脑健康维护、认知能力保护和神经功能恢复的关键天然营养素。
4. 供应链格局分析
神经酸的传统来源包括深海鲨鱼脑组织、少量的元宝枫树籽油、文冠果油及亚麻荠籽油等植物资源。现代生物发酵及酶解技术创新让高纯度神经酸98%从天然提取及生物合成技术路线成为可靠且可控的工业级原料。
2026年,纳普生物的“亚麻荠籽油”获批国家新食品原料。其核心营养价值在于富含α-亚麻酸与神经酸,扩大了神经酸稳定来源。
据2026年4月投资者互动平台公开信息,诚意药业作为以核酸药物与海洋药物研发为基础的上市药企,其神经酸项目已进入药学研究阶段,正处于从小试向中试放大推进的加速期。此举有望使神经酸原料得到上游供给扩容和技术革新的切实保障。这项最新动态为长期依赖植物提取供应的神经酸市场打开了另一条化学合成技术路线。
三、神经酸的法规准入与食品级应用范畴
原料的商业化应用受制于国家及跨境出口法规和食品标准门槛的严格限制。神经酸在食品领域的应用在中国已有稳固的政策基础:
国内法规基础
:国家卫健委2017年第7号公告已批准以菜籽油为原料,通过皂化、酸化、萃取、分离等工艺制得的顺-15-二十四碳烯酸为新食品原料。此外,2011年第9号公告也已批准元宝枫树种仁来源的元宝枫籽油为新食品原料,其脂肪酸质量要求为神经酸≥3.0%,此原料在糖果、方便食品及保健食品中被广泛使用。
GB 14880合规条件
:2014年起,特殊膳食食品及国家保健食品的整体原料严控依据GB14880,为品牌方的配方备案铺平了道路。在使用过程中,神经酸既可作为普通食品原料功能宣称,也可依据科学数据进行特定功效配方设计。
国际市场出口要求
:由于神经酸原料动物和植物来源的绿色属性,符合KOSHER、HALAL、欧盟及美国有机多重认证,欧美和东南亚市场出口时通常不设额外政策障碍,只需配合食品添加剂及膳食补充剂原料标准达标。
四、国内外脑健康产品创新案例
神经酸作为脑健康功能性原料,在国内外已有多款成熟终端产品上市。以下精选代表性产品进行拆解分析:
案例一:迈昔龄神经酸片(迈之灵(深圳))
迈昔龄神经酸片以神经酸为主要功能成分,科学复配芹菜粉、酸枣仁、益生菌与益生元、GABA及酪蛋白水解肽等药食同源物质。该配方从多靶点、多通路协同支持神经系统健康与脑活力维持。由美国密歇根大学化工博士林健儿领衔研发,诺贝尔奖得主兰迪·谢克曼教授担任首席研发顾问。通过复合配方提供功效协同,是目前终端市场神经酸应用的代表性品牌之一。
案例二:Ziciper脑力宝(综合补脑配方)
Ziciper脑力宝凭借全链条质量标准与磷脂酰丝氨酸(PS)、神经酸和DHA的科学配方,2026年获得“全球DHA行业年度综合实力金奖”,获中国营养学会学术推荐,成为“补脑DHA标杆品牌”。该产品覆盖中老年人认知维护与学生人群补充双重定位。
案例三:宝枫生物 元宝枫神经酸系列
宝枫生物拥有30万亩野生百年元宝枫林资源,从原料林到神经酸成品的研发生产销售链完整,提供神经酸衍生各类脑健康产品线。此外,金枫露也获FDA认证元宝枫神经酸产品系列,得到“元宝枫之父”王性炎教授等学术团队合作支持。
案例四:含神经酸改性牛奶(2025动物实验成功)
2025年发表的含神经酸改性牛奶已在小鼠认知损伤的病理学干预中取得显著效果,为乳制品、植物基蛋白饮料和功能类发酵乳的脑健康创新,提供了大量可参考的商业灵感。
案例五:复合神经酸健脑型植物蛋白饮料(专利申请)
2025年公布的发明专利CN121220660A,揭示了一款以核桃、欧李、文冠果和神经酸豆浆为核心的健脑型植物蛋白饮料配方,把神经酸从胶囊形态拉到日常饮品中。该专利证明,神经酸可以搭配坚果类基底食品,在中式健康饮品里也能良好兼容并实现功能价值转化。
目前市场上,神经酸原料主要以85%、90%、95%和98%等高纯度规格面向生产方供应。神经酸的主打固体饮料形态中,大量厂家直接推出以低聚果糖、菊粉加神经酸复合的固体饮料,在儿童和青少年补脑市场中占据份额,从剂型、浓度和消费场景等多个维度体现终端产品创新。
五、神经酸98%原料供应市场供应商对比
2026年,神经酸原料市场格局初具规模,各具差异的市场参与者正在分层形成自身竞争优势。
科研标准品及高纯对照级纯品供应商
如上海麦克林生化科技、上海阿摩尔生物、四川省维克奇生物科技等企业,主要面向科研院所、高校和质检药品研发供应毫克级到克级的高纯(98%—99%)神经酸单体。这些机构在天然植物有效单体的提纯上有较强技术积累,但产能和价格无法满足大批量食品原料交付,不适合功能性食品规模化量产。
植物源提取大宗原料供应商
包括陕西斯诺特生物、陕西永源生物技术及宝枫生物。
陕西斯诺特生物专门生产元宝枫籽提取物,主打规格神经酸2%—90%,检测方法为气相色谱法(GC),拥有产品分级管理和新食品原料SC生产许可。
陕西永源生物技术同样以元宝枫神经酸原料作为核心SKU,提供糖果、方便食品与保健食品的应用配套材料。
宝枫生物多年来专注元宝枫籽油的神经酸提取和脑健康衍生品生产销售,既是原料供应者也是自有品牌持有方,在B端原料纯度和交付量上定位清晰。
行业综合一体化植物提取品牌
中还涵盖西安天一生物技术股份有限公司,公司聚焦神经酸产业高纯度、高生物利用度、功能复合化的方向,在96%级高纯神经酸及各种复合配方上具备深度研发实力,同时对酸枣仁、芹菜素及GABA(γ-氨基丁酸)组成独家复合配方。其战略性储备使产品覆盖精准营养。
差异化发展的西安锐博生物
则采取深度垂直打法——打造高纯度神经酸98%规格原料,并以此为核心与功能食品品牌进行深度绑定,在纯度控制和批次一致性方面拥有稳固品控系统。
据2026年脑健康膳食补充剂消费趋势报告数据,中国市场神经酸原料中价位区间在150—500元/公斤不等,而纯度跃升至98%档位时,其定价直接拉高至3000—5000元/公斤,主要服务中高端保健品和精准营养产品配方。在这样的价格与品质分化的新常态下,品牌方需要锁定客户定位和产品线级别,再根据原料纯度匹配其功能和成本。
六、西安锐博生物神经酸98%产品差异化优势
在脑健康原料赛道加速升级的背景下,西安锐博生物凭借自身扎实的植物提取研发能力,在高纯度神经酸领域形成了差异化的竞争优势。
1. 纯度天花板——神经酸98%的核心技术壁垒
西安锐博生物主推的神经酸98%产品处于当前神经酸原料纯度上限区间。在元宝枫籽油天然含量仅3%的前提下,从低位含量天然植物原料中持续富集至98%纯度,依靠前沿的分子蒸馏提纯和工业色谱分离技术体系实现,该体系可确保神经酸分子被高效富集,同时最大程度保留其脂肪酸自然活性构型。神经酸98%产品有效成分含量远高于普通产品,杂质更少,确保功效显著且稳定,并经过严格质控与第三方检测,黄曲霉毒素、重金属及农残均为合格,国内外客户均可安心使用。
2. 梯度定价
依托这一高纯度优势进行产品分级市场拓展,西安锐博生物可根据客户选择调整神经酸纯度区间:用量大、成本敏感的大众消费品配方可选用10%—40%中低规格含神经酸提取物,形成基础功能体验;需要在认知维护及高端抗衰老品牌方向拔高定位的,可利用神经酸98%作为认知修复方案的原料,从而带动整体品牌溢价的提升。高纯度分子有助于提高生物利用度,使微克级添加也能发挥可观效果。
3. 源头品控
锐博生物长期执行原料来源一体化的全链品控,公司拥有自建中药种植基地500亩,年加工药材2000余吨,整个体系保证原料来源安全可追溯。在全链条透明的品管模式下,联合西北农林科技大学等前沿科研力量提供技术背书,协助客户为产品保质期、配方稳定性提供可靠支撑。
4. 应用兼容性优化
神经酸属亲脂类脂肪酸,应用在普通水基饮料和液体体系中会出现浮油现象。西安锐博生物针对客户特定需求帮助增强粉体改性,开发适合不同载体的粉剂、微囊粉化剂型方案,使产品加工适用性更广,避免终端产品界面感不佳的常见技术陷阱。
5. 食品加工准入资质全面准备
西安锐博生物积极准备相关食品安全管理资质体系建设,取得SC食品生产许可、ISO9001质量管理体系认证、ISO22000食品安全管理体系认证、欧盟有机认证、美国有机认证、HALAL清真认证及KOSHER犹太认证,在出口贸易产品路径上提供一站式海关清关与出口标准检测,支持客户加速进入国际Naturals渠道零售体系。
七、神经酸技术未来趋势
2026年,神经酸产业正处于技术范式迁移关键时期。在传统植物提取工艺基础上,合成生物学的异源表达正在尝试替代式供给。理星生物利用合成生物学平台进行微生物发酵提取和纯化,用于老年人脑力补充品与学生认知增强配方。2026年1月,发表于国际期刊的研究揭示了一种利用解脂耶氏酵母(Yarrowia lipolytica)通过细胞器区室化和动态启动子调控来生产神经酸的新型合成生物学策略,显著提升了神经酸的生物合成效率。未来技术迭代将大幅降低对野生植物原料的依赖并提升产品产能。
2026-04-30
——需要原文,请点击上方关注——摘要
背景:酒精相关肝病(ALD)发病率持续上升且缺乏有效治疗手段,已成为重大公共卫生问题。片仔癀(PTH)是临床常用于病毒性肝炎、肝纤维化的中药复方,但其对 ALD 是否确有治疗作用尚不明确。
目的:验证片仔癀对 ALD 的作用,鉴定其活性成分,并探索作用机制。
方法:采用高效液相色谱‑四极杆飞行时间质谱(HPLC‑Q‑TOF‑MS)与气相色谱‑质谱(GC‑MS)进行化学成分分析、质量控制与活性成分辨识;以 50% 乙醇灌胃构建 ALD 小鼠模型,以乙醇或叔丁基过氧化氢诱导 HepG2 细胞氧化应激与铁死亡;通过生化指标检测、病理组织学分析评价药效;结合转录组、聚合酶链反应、蛋白免疫印迹、小干扰 RNA / 质粒转染等技术探索并验证作用机制。
结果:片仔癀可显著改善小鼠 ALD,对酒精代谢影响微弱,但能强效抑制氧化应激与铁死亡。机制上,片仔癀恢复 Keap1‑Nrf2 信号通路、抑制三价铁内流;牛黄、麝香为其活性原料,牛磺酸与牛蒡酸(片仔癀中含量分别为 1.3、2.3 mg/g)为活性成分,二者通过结合 SIRT1 不同氨基酸残基,经 SIRT1/Keap1/Nrf2 轴与 SIRT1 / 转铁蛋白受体通路抑制氧化应激与铁死亡。
结论:片仔癀中的牛磺酸与牛蒡酸,通过 SIRT1/Keap1/Nrf2 轴介导的氧化应激抑制、SIRT1 / 转铁蛋白受体介导的铁死亡抑制,实现对 ALD 的改善作用。关键词
酒精性肝病;片仔癀;氧化应激;铁死亡;牛蒡酸;牛磺酸1、引言
酒精相关肝病(ALD)是全球性重大健康负担,全球患病率约 3.5%,高危饮酒人群中达 26.0%。长期过量饮酒引发的慢性肝脏病变,会导致肝脏结构与功能受损,涵盖肝脂肪变性、酒精性脂肪性肝炎、肝纤维化、肝硬化及肝细胞癌等一系列病症。ALD 已成为欧美地区肝移植首要指征,约占全球全因死亡率的 6%。
ALD 患病率高且缺乏有效疗法,目前最有效的干预手段为戒酒,但患者依从性差;临床仅糖皮质激素可用,疗效有限。因此亟需研发新型有效治疗药物。
明确 ALD 复杂发病机制是研发治疗策略的前提,其病理过程涉及脂质蓄积、细胞器应激、肝细胞凋亡、免疫炎症、肝星状细胞介导纤维化等多因素级联反应,核心特征为氧化还原稳态失衡,即活性氧(ROS)生成与清除失衡。过量饮酒代谢诱发 ROS 蓄积,进一步引发脂质过氧化、线粒体功能障碍与铁死亡。氧化应激受 Kelch 样 ECH 相关蛋白 1(Keap1)‑核因子 E2 相关因子 2(Nrf2)轴精密调控:生理状态下 Nrf2 与 Keap1 结合并被泛素化降解;氧化应激时,Keap1 构象改变使 Nrf2 解离并入核,激活抗氧化相关基因转录,维持细胞氧化还原稳态。Keap1‑Nrf2 通路受沉默信息调节因子 1(SIRT1,Ⅲ 类组蛋白去乙酰化酶)调控,SIRT1 可调节 Keap1‑Nrf2 轴增强抗氧化防御,延缓 ALD 等多种肝病进展。
氧化应激与铁死亡密切相关。铁死亡是铁依赖性的调控性细胞死亡,以广泛脂质过氧化物蓄积为特征,可被氧化应激启动并加重。铁死亡中 ROS 主要来源为过氧化氢与二价铁的芬顿反应,转铁蛋白受体介导的三价铁过量内流是芬顿反应的关键条件。大量研究表明,氧化应激与铁死亡参与 ALD 等多种肝病进程,因此靶向干预上述病理过程对改善疾病预后意义重大。
中药是肝病药物研发的重要来源,疗效确切、安全性良好,且具备多靶点调控优势,适用于复杂病理机制疾病的治疗,但作用机制与活性成分阐释难题限制其全球应用。片仔癀是知名中药复方,由三七(85%)、麝香(3%)、牛黄(5%)、蛇胆(7%)组成,被列入国家中药保护名录,对病毒性肝炎、肝纤维化等疾病疗效显著,但其对 ALD 的作用尚未明确。
本研究旨在:(1)建立片仔癀质量控制指纹图谱;(2)评价片仔癀对小鼠 ALD 的治疗作用;(3)探索其作用机制;(4)鉴定片仔癀的关键活性原料与成分。研究结果将为片仔癀用于 ALD 治疗提供科学依据,并建立中药复方活性成分辨识的完整范式。2、材料与方法药品与试剂片仔癀复方及三七、蛇胆、牛黄、麝香均由漳州片仔癀药业股份有限公司提供,按《中国药典》严格质控。水飞蓟宾胶囊购自天津天士力圣特制药有限公司;甲磺酸去铁胺(DFOM)、铁抑素‑1(Fer‑1)、牛磺酸、牛蒡酸(BA)、萝卜硫素(SPN)、N‑乙酰半胱氨酸(NAC)购自 TargetMol 公司;叔丁基过氧化氢(tBHP)购自上海麦克林生化科技有限公司;药用乙醇购自 Sigma‑Aldrich 公司。
细胞计数试剂盒(CCK‑8)购自南京诺唯赞生物科技有限公司;2',7'‑二氯二氢荧光素二乙酸酯检测试剂盒购自 MCE 公司;谷胱甘肽(GSH)、丙二醛(MDA)、超氧化物歧化酶(SOD)、总胆固醇(TC)、甘油三酯(TG)、乙醇检测试剂盒购自南京建成生物工程研究所;二价铁检测试剂盒购自武汉伊莱瑞特生物科技有限公司;丙氨酸氨基转移酶(ALT)、天冬氨酸氨基转移酶(AST)检测试剂盒购自南京纽松特维特曼生物科技有限公司。
TRIzol 试剂购自大连宝生物工程有限公司;SYBR Green 与反转录试剂盒购自南京诺唯赞生物科技有限公司;无酶水购自上海碧云天生物技术有限公司;人源 SIRT1(His 标签)蛋白购自 MCE 公司;DMEM 培养基、Lipofectamine™ RNAiMAX 与 Lipofectamine™ 3000 转染试剂购自赛默飞世尔科技有限公司。
质粒包括 pcDNA3.1 (+)、pcDNA3.1 (+)‑KEAP1–3HA、pcDNA3.1 (+)‑TFRC‑3HA、pcDNA3.1 (+)‑SIRT1(野生型)、pcDNA3.1 (+)‑SIRT1 D272A/R274A/S275A/S442A(突变体 1)、pcDNA3.1 (+)‑SIRT1 R274A/S441A(突变体 2),购自南京金载生物科技有限公司;小干扰 RNA(si‑SIRT1、si‑NC)购自南京益科生物科技有限公司;RIPA 裂解液、磷酸酶抑制剂、蛋白酶抑制剂购自上海碧云天生物技术有限公司。片仔癀不同药物溶液制备亲水性成分工作液:片仔癀粉末 1.0 mg 悬浮于 1.0 ml 培养基,涡旋 2 min,4℃、12000×g 离心 10 min,取上清即为亲水性成分工作液。亲脂性 / 两亲性成分工作液:片仔癀粉末 1.0 mg 悬浮于 1.0 ml 培养基,涡旋 2 min,4℃、12000×g 离心 10 min,弃上清,沉淀用 50 μl 二甲基亚砜(DMSO)溶解,再用 1.0 ml DMEM 稀释,即为亲脂性 / 两亲性成分工作液。总成分工作液:按上述方法制备亲水性成分上清,沉淀用 DMSO 溶解后与上清混合,即为总成分工作液。所有工作液用 DMEM 稀释 20 倍后用于细胞孵育。
不同储备浓度片仔癀亲脂性 / 两亲性成分工作液制备
分别取 0.2、0.5、1.0 mg 片仔癀粉末,按上述方法制备亲脂性 / 两亲性成分工作液,稀释 20 倍后使用。
不同离心速度片仔癀亲脂性 / 两亲性成分工作液制备
取 1.0 mg 片仔癀粉末,分别以 3000、6000、12000×g 离心 10 min,按上述方法制备亲脂性 / 两亲性成分工作液,稀释 20 倍后使用。
四种原料不同药物溶液制备
分别取三七(0.85 mg)、牛黄(0.05 mg)、蛇胆(0.07 mg)、麝香(0.03 mg)粉末,按片仔癀溶液制备方法,分别得到亲水性、亲脂性 / 两亲性、总成分工作液,稀释 20 倍后使用。
牛黄与麝香混合液不同药物溶液制备
取牛黄(0.05 mg)、麝香(0.03 mg)粉末,按上述方法制备亲水性、亲脂性 / 两亲性、总成分工作液,稀释 20 倍后使用。片仔癀复方色谱与质谱分析样品制备
药物工作液:0.5 ml 药物工作液与 0.5 ml 甲醇混合,涡旋 5 min,4℃、18000×g 离心 5 min;取 0.9 ml 上清氮气吹干,用 0.1 ml 50% 甲醇复溶,涡旋 5 min、离心后取 80 μl 上清用于 HPLC‑Q‑TOF‑MS 分析;另取样品氮气吹干后用 100 μl 纯甲醇复溶,离心后用于 GC‑MS 分析。
细胞样品:干预后 6 孔板细胞经超纯水反复冻融裂解,取 0.5 ml 裂解液按上述方法提取处理,用于 HPLC‑Q‑TOF‑MS 分析。
HPLC‑Q‑TOF‑MS 分析
采用岛津 LC‑30A 高效液相色谱系统、AB Sciex Triple TOF™ 5600 质谱系统,电喷雾电离源,Analyst TF 1.6 工作站。色谱柱为 Waters Xselect® HSS T3(3.5 μm,4.6×150 mm),柱温 40℃;流动相 A 为含 0.05% 甲酸的超纯水,流动相 B 为乙腈,梯度洗脱;流速 0.5 ml/min,进样量 10 μl,分析时间 34 min。质谱采用正负离子模式,参数按标准条件设置,用 MultiQuant 3.0.3 软件定量。
GC‑MS 分析
采用岛津 GCMS‑QP2010 Ultra 质谱仪、AOC‑20i 自动进样器,GCMS solution 4.11 SU2 工作站。色谱柱为 Rtx‑5MS 毛细管柱(30 m×0.25 mm×0.25 μm),分流模式(分流比 10:1),进样口温度 210℃,进样量 0.8 μl;载气为氦气,流速 1.21 ml/min;柱温初始 90℃保持 3 min,以 40℃/min 升至 250℃保持 3 min。质谱离子源温度 220℃,电子能量 70 eV,选择离子监测模式检测 m/z 209,运行时间 31 min,溶剂切割时间 3.5 min。
动物实验与处理
所有动物实验遵循《赫尔辛基宣言》伦理准则,经中国药科大学动物伦理委员会批准(批准号:2024‑11‑013)。8 周龄 SPF 级 C57BL/6J 雄性小鼠,饲养于温度(22±1)℃、湿度(50±10)%、12 h 明暗循环环境,自由摄食饮水。
ALD 小鼠模型构建:小鼠每日灌胃 50%(v/v)乙醇溶液,连续 10 d;末次给药 7 h 后麻醉脱颈处死,采集血清与肝脏组织样本。
片仔癀药效实验:小鼠随机分为 5 组(n=6):对照组、模型组、片仔癀低剂量组(0.2 g/kg)、片仔癀高剂量组(0.4 g/kg)、水飞蓟宾组(阳性对照,0.15 g/kg);每日灌胃给药 1 次,连续 10 d,对照组与模型组给予等体积溶媒。
活性原料筛选实验:小鼠随机分为 7 组(n=6):对照组、模型组、片仔癀组(0.4 g/kg)、三七组(0.34 g/kg)、蛇胆组(0.028 g/kg)、牛黄组(0.02 g/kg)、麝香组(0.012 g/kg);给药方式同前。
牛磺酸与牛蒡酸药效实验:小鼠随机分为 6 组(n=6):对照组、模型组、牛磺酸组(0.2 g/kg)、牛蒡酸组(0.1 g/kg)、牛磺酸 + 牛蒡酸组(0.2、0.1 g/kg)、水飞蓟宾组(0.15 g/kg);给药方式同前。
组织学分析
肝脏组织用 4% 多聚甲醛固定、石蜡包埋、切片,行苏木精‑伊红(H&E)染色;肝脏组织用 OCT 包埋、冰冻切片,行油红 O 染色,异丙醇分化、苏木精复染。
血清生化分析
按试剂盒说明书检测血清 ALT、AST、TC、TG、GSH、SOD、MDA、二价铁、乙醇浓度。
细胞培养与转染
HepG2 细胞购自美国典型培养物保藏中心,用含 10% 胎牛血清、1% 青霉素‑链霉素的 DMEM 培养基,于 37℃、5% CO₂培养箱培养。按说明书用 Lipofectamine™ RNAiMAX 或 Lipofectamine™ 3000 试剂转染 siRNA 或质粒。
细胞活力检测
按 CCK‑8 试剂盒说明书检测细胞活力。
细胞内 ROS 水平分析
细胞用含 10 μM 2',7'‑二氯二氢荧光素二乙酸酯的培养基孵育 30 min,PBS 洗涤后离心,用 Synergy H1 多功能酶标仪检测 488/525 nm 荧光强度,数据经细胞活力校正。
网络药理学分析
通过 TargetNet、ETCM、SwissTargetPrediction 数据库预测牛蒡酸、牛磺酸潜在靶点;GeneCards 数据库检索 ALD 相关靶点;经 UniProt 数据库标准化后,取交集得到牛蒡酸 / 牛磺酸治疗 ALD 的潜在靶点;用 STRING 数据库构建蛋白互作网络,进行功能富集分析,经 PubMed 文献检索筛选氧化应激 / 铁死亡相关靶点。
分子对接
收集雌激素受体 α(1XPC)、瞬时受体电位香草酸 1(8GFA)、KEAP1(7K2L)、SIRT1(4KXQ)、COUP 转录因子 2(AF‑P10588‑F1‑model_v6)结构,用 Autodock Vina 1.1.2 进行分子对接,对接盒设为 20 Å×20 Å×20 Å,重复 20 次,取最优对接构象分析。
生物膜干涉(BLI)实验
用 Octet R8e 系统检测牛磺酸、牛蒡酸与人源 SIRT1 重组蛋白的直接结合,NTA 传感器捕获 SIRT1‑His 蛋白,分析与小分子的结合动力学。
RNA 提取
按常规方法用 TRIzol 试剂提取肝脏组织或细胞总 RNA,氯仿抽提、异丙醇沉淀、75% 乙醇洗涤,无酶水复溶。
转录组分析
RNA 样本经 poly (A) 尾筛选获取 mRNA,反转录构建 cDNA 文库,BGI T7 平台测序;用 SOAPnuke v2.2.1 过滤原始数据,HISAT2 v2.2.1 比对参考基因组,用 Dr. Tom 多组学数据挖掘系统进行生物信息学分析。
聚合酶链反应(PCR)
按说明书用 HiScript III RT SuperMix 进行反转录,ChamQ SYBR qPCR MasterMix 进行荧光定量 PCR,用 2⁻^ΔΔ^Ct 法计算基因相对表达量。
蛋白免疫印迹(Western blot)
细胞用含蛋白酶、磷酸酶抑制剂的 RIPA 裂解液裂解,SDS‑PAGE 电泳分离蛋白,转印至聚偏二氟乙烯膜,5% 脱脂牛奶封闭;一抗(抗 Nrf2、抗 Keap1、抗 β‑actin)4℃孵育过夜,二抗室温孵育 1 h;超敏 ECL 底物显色,化学发光成像系统成像,ImageJ 软件定量条带灰度值。
统计学分析
用 GraphPad Prism 9.0 软件分析数据,结果以均值 ± 标准误(SEM)表示;多组间比较采用单因素方差分析(ANOVA)+Tukey 事后检验,两组比较采用 t 检验;P<0.05 为差异具有统计学意义。
3、结果
片仔癀化学成分表征
采用 HPLC‑Q‑TOF‑MS 与 GC‑MS 对片仔癀进行严格质控,共鉴定出 46 种化合物,选定三七皂苷 R1、人参皂苷 Rg1、人参皂苷 Rb1、胆酸(CA)、脱氧胆酸(DCA)、麝香酮为质控标志物并定量;同时测定牛磺酸、牛蒡酸含量,方法学验证符合国际协调理事会标准。
三七来源的三七皂苷 R1、人参皂苷 Rg1、人参皂苷 Rb1 含量分别为 3.1、2.6、2.9 mg/g;牛黄、蛇胆来源的胆酸、脱氧胆酸含量分别为 1.6、2.3 mg/g;麝香来源的麝香酮含量为 3.3 mg/g;牛黄来源的牛磺酸、牛黄与麝香来源的牛蒡酸含量分别为 1.3、2.3 mg/g。上述结果为片仔癀质控与活性成分辨识提供依据。
片仔癀改善 ALD 氧化应激
片仔癀临床用于病毒性肝炎剂量为 1.8 g/d,小鼠等效剂量 0.36 g/kg;安全性实验显示,0.2、0.4、2 g/kg 剂量连续灌胃 15 d,小鼠血清生化、血液学、主要脏器重量与组织学均无异常,安全性良好。
构建乙醇诱导 ALD 小鼠模型,水飞蓟宾为阳性对照。模型组小鼠血清 ALT、AST 显著升高,片仔癀高剂量、水飞蓟宾可显著降低上述指标,低剂量效果较弱;肝组织 H&E 染色显示,模型组肝索紊乱、肝细胞气球样变、炎症浸润明显,片仔癀与水飞蓟宾可显著改善肝组织形态、减轻炎症;油红 O 染色显示,片仔癀与水飞蓟宾可减少肝脂质蓄积,降低肝 TC、TG 含量,呈剂量依赖性。
片仔癀对血清乙醇浓度、酒精代谢酶(ADH、ALDH、CYP2E1)基因表达无明显影响,细胞实验验证其不直接调控乙醇代谢;但可剂量依赖性降低乙醇 / 叔丁基过氧化氢诱导的 HepG2 细胞 ROS、MDA 水平,升高 GSH、SOD 活性,证实其可独立于酒精代谢通路,强效改善氧化损伤。
片仔癀通过调控 Keap1‑Nrf2 信号与二价铁稳态改善氧化应激与铁死亡
MDA 是脂质过氧化终产物,也是氧化损伤与铁死亡标志物,片仔癀可显著降低 MDA 水平,提示其可能改善铁死亡相关氧化应激。转录组分析筛选出 164 个氧化应激模型‑对照组、片仔癀处理‑模型组的共调控差异基因(DEGs);基因本体(GO)富集显示,差异基因主要参与金属离子应答与稳态、氧化应激应答、生长调控等,血红素氧合酶 1(HMOX1,Keap1‑Nrf2 通路下游基因)富集于氧化应激应答类别;京都基因与基因组百科全书(KEGG)富集显示,差异基因与肿瘤坏死因子、白介素 17、叉头框 O、胰高血糖素信号、胰岛素抵抗、铁死亡、矿物质吸收等通路相关,HMOX1 亦富集于铁死亡通路,提示片仔癀通过铁死亡与氧化应激信号发挥抗氧化作用。
片仔癀可剂量依赖性降低 HepG2 细胞 KEAP1 mRNA 与蛋白表达,升高 NRF2、HMOX1 mRNA 及 Nrf2 蛋白表达,ALD 小鼠体内实验验证一致结果。铁死亡相关检测显示,片仔癀可特异性剂量依赖性降低乙醇 / 叔丁基过氧化氢诱导的 HepG2 细胞 TFRC mRNA 表达,对 GPX4、ACSL4、SLC7A11、FSP1、NOX1 无明显影响;同时剂量依赖性降低细胞内二价铁水平。上述结果表明,片仔癀通过调控 Keap1‑Nrf2 通路、抑制铁内流发挥抗氧化作用。
牛黄与麝香是片仔癀抑制氧化应激与铁死亡的活性原料
片仔癀含三七、麝香、牛黄、蛇胆四种原料,分别检测各原料作用:四味混合组作用与片仔癀相当,牛黄、麝香可显著恢复 ROS、MDA、二价铁水平及 SOD、GSH 活性,三七、蛇胆无明显作用;牛黄与麝香联用可协同改善上述指标。
ALD 小鼠体内实验验证,牛黄、麝香可显著降低血清 ALT、AST,改善肝组织病理与脂质蓄积,降低肝 TC、TG,升高肝 SOD 活性、降低 MDA 水平,证实牛黄、麝香是片仔癀的活性原料。
片仔癀亲脂性与两亲性组分发挥抗氧化与抗铁死亡作用
将片仔癀分为亲水性、亲脂性 / 两亲性、总成分组分,检测显示总成分、亲脂性组分可显著降低乙醇 / 叔丁基过氧化氢诱导的细胞 ROS、MDA、二价铁水平,升高 SOD、GSH 活性,亲水性组分无明显作用,提示亲脂性组分是抗氧化、抗铁死亡的主要有效组分。
优化分离条件:1 mg/ml 储备浓度、12000×g 离心速度下,亲脂性组分活性最优,证实离心参数对活性组分分离至关重要。
片仔癀中牛磺酸与牛蒡酸抵御氧化应激与铁死亡
牛黄、麝香的亲脂性 / 两亲性组分可显著恢复氧化应激、铁死亡标志物水平,亲水性组分无作用;三七、蛇胆的亲脂性 / 两亲性组分亦无作用。对牛黄、麝香亲脂性 / 两亲性组分进行 HPLC‑Q‑TOF‑MS 分析,细胞摄取组分中鉴定出 8 种化合物:猪脱氧胆酸(HDCA)、熊脱氧胆酸(UDCA)、鹅脱氧胆酸(CDCA)、脱氧胆酸(DCA)、柠檬酸、牛蒡酸、5‑环十六烯‑1‑酮、牛磺酸;细胞实验证实仅牛磺酸、牛蒡酸可剂量依赖性抑制 ROS 水平,为片仔癀的活性成分。
牛磺酸与牛蒡酸协同抑制氧化应激与铁死亡
片仔癀中牛磺酸、牛蒡酸含量分别为 1.3、2.3 mg/g,50 μg/ml 片仔癀中二者浓度约 0.5 μM,该浓度即可显著改善氧化应激、铁死亡指标。设置牛磺酸(0.5、5、50 μM)、牛蒡酸(0.5、5、50 μM)浓度梯度,细胞实验显示二者均可剂量依赖性降低乙醇 / 叔丁基过氧化氢诱导的 ROS、MDA、二价铁水平,升高 SOD、GSH 活性;二者联用效果优于单用。
ALD 小鼠体内实验显示,牛磺酸、牛蒡酸单用或联用均可降低血清 ALT、AST,改善肝组织病理与脂质蓄积,降低肝 TC、TG,升高肝 SOD 活性、降低 MDA 水平,联用效果最优,证实牛磺酸、牛蒡酸是片仔癀抵御氧化应激与铁死亡的核心成分。
牛磺酸与牛蒡酸调控 Keap1‑Nrf2 信号
氧化应激核心调控通路为 Keap1‑Nrf2 轴,蛋白免疫印迹显示,牛磺酸、牛蒡酸可降低乙醇 / 叔丁基过氧化氢诱导的 HepG2 细胞 Keap1 蛋白表达,升高 Nrf2 蛋白表达与 HMOX1 mRNA 水平;ALD 小鼠体内实验验证一致结果。Keap1 过表达质粒转染后,牛磺酸、牛蒡酸无法恢复 ROS、SOD 水平,证实二者通过 Keap1‑Nrf2 信号抑制氧化应激。
牛磺酸与牛蒡酸调控三价铁稳态
铁死亡依赖芬顿反应与铁内流,牛磺酸、牛蒡酸可显著降低乙醇 / 叔丁基过氧化氢诱导的 HepG2 细胞 TFRC mRNA 表达,减少二价铁蓄积;TFRC 过表达质粒转染或铁螯合剂 DFOM 处理后,二者无法恢复 GSH、MDA、二价铁及 ROS、SOD 水平,证实二者通过 TFRC 介导的铁内流抑制铁死亡。
牛磺酸与牛蒡酸直接结合 SIRT1 抑制氧化应激与铁死亡
网络药理学筛选出牛磺酸‑ALD、牛蒡酸‑ALD 共同靶点,分子对接与 BLI 实验证实,SIRT1 是二者共同高亲和力靶点:牛磺酸结合 SIRT1 的 D272、R274、S275、S442 位点,牛蒡酸结合 SIRT1 的 R274、S441 位点。
SIRT1 沉默(si‑SIRT1)后,牛磺酸、牛蒡酸无法改善氧化应激、铁死亡指标;构建 SIRT1 野生型、突变体 1(牛磺酸结合位点突变)、突变体 2(牛蒡酸结合位点突变)质粒,回补实验显示,野生型 SIRT1 可恢复二者作用,突变体则无作用,证实牛磺酸、牛蒡酸通过直接结合 SIRT1 抑制氧化应激与铁死亡。
4、讨论
ALD 发病率攀升且缺乏有效疗法,本研究证实片仔癀可通过改善氧化应激与铁死亡显著缓解 ALD;明确牛黄、麝香为活性原料,亲脂性 / 两亲性组分为活性组分,牛磺酸、牛蒡酸为核心活性成分;机制上,二者通过 SIRT1/Keap1/Nrf2 轴抑制氧化应激,通过 SIRT1 / 转铁蛋白受体通路抑制铁内流与铁死亡。研究为中药复方活性成分与靶点辨识提供范式,也为 ALD 治疗提供候选药物与先导化合物。
片仔癀临床常用于病毒性肝炎、肝纤维化,近年被证实可抑制结直肠癌,但其对 ALD 的作用与机制尚未明确。本研究首次证实片仔癀对 ALD 具有明确治疗效果,为临床应用提供依据。
酒精代谢是 ALD 核心诱因,但片仔癀不影响酒精代谢与代谢酶表达,而是通过调控氧化还原稳态发挥作用:酒精诱导 ROS 蓄积激活 Keap1‑Nrf2 通路,片仔癀可恢复 ROS、SOD、MDA、GSH 水平,调控 Keap1‑Nrf2 通路;同时酒精诱导肝铁过载,触发铁死亡,片仔癀可抑制 TFRC 表达、减少二价铁蓄积,靶向铁内流通路抑制铁死亡。
中药复方成分复杂,本研究通过 “原料筛选‑组分分离‑细胞摄取‑活性验证” 的策略,精准锁定牛磺酸、牛蒡酸为活性成分。牛磺酸是牛黄中丰富的非必需氨基酸,具备抗氧化、抗炎、抗凋亡作用;牛蒡酸是反式共轭亚油酸,存在于反刍动物肉脂中,二者均通过 Keap1‑Nrf2 通路发挥抗氧化作用,联用效果增强。
SIRT1 是氧化还原稳态关键调控因子,可调控 Keap1‑Nrf2 轴与铁代谢,本研究证实牛磺酸、牛蒡酸通过结合 SIRT1 不同位点,发挥抗氧化、抗铁死亡作用,为靶点调控提供新依据。
5、结论
片仔癀通过抑制氧化应激与铁死亡改善 ALD;其中牛磺酸与牛蒡酸通过直接结合 SIRT1 不同氨基酸残基,分别抑制 Keap1‑Nrf2 轴介导的氧化应激、TFRC 介导的铁内流,发挥核心治疗作用。本研究为片仔癀临床治疗 ALD 提供坚实证据,鉴定出牛磺酸、牛蒡酸为 ALD 治疗先导化合物,并建立中药复方复杂体系活性成分与直接分子靶点的系统辨识范式。
需要原文PDF,请关注并私信。
💛 感谢阅读,关注我们,一起前行。
关注下方二维码👇,关注后续精彩内容~~~
往期精彩推荐
Phytomedicine:中药药效物质基础及作用机制研究-基于多模型与植物化学分析揭示天南星最优成分抗阿尔茨海默病作用机制
CEJ(IF=13.2) | 中药苦参碱 - 没食子酸自组装纳米颗粒,实现多靶点心肌保护 ,改写心梗治疗格局
Carbohydr Polym (IF=12.5)|中药自组装-果胶凝胶搭载栀子苷,双靶点攻克胆汁淤积性肝病
Food Hydrocolloids (IF=12.4)| 香菇多糖搭伙淫羊藿次苷,实现抗炎活性与稳定性双突破
J Adv Res(IF=13.0)|药食同源中药成分木犀草素-玉竹-智能纳米组装平台,赋能癌症化学免疫治疗
MTB(IF=10.2)| 泻心汤里的天然 “纳米小药”!精准掐断结肠炎的炎症开关
Phytomedicine(IF=8.3):中药自组装研究-附子与甘草药对自组装的治疗心衰的药效物质基础表征新思路
Adv Funct Mater(IF=19.0)|分子自组装新策略:天然中药活性成分苦杏仁苷与甘草酸自组装水凝胶,为创伤性脑损伤术后治疗提供可选策略
Redox Biol(IF=11.9)| 抗癌不损心:破解阿霉素心脏毒性困局,为肿瘤化疗筑起心脏防线
Bioact Mater(IF=20.3)| 传统中药活性成分与智能纳米递送系统强强联合,让心肌梗死治疗更精准、更持久!
APSB(IF 14.6)| 中国药科大学—顶刊突破!中国团队发现肝癌耐药新靶点,金雀花碱衍生物 DC218 精准破局
APSB(IF 14.6)|中医智慧遇上纳米科技!南中医团队用黄芪外泌体,攻克三阴性乳腺癌化疗难题
Phytomedicine(IF 8.3)|灵芝再立新功!灵芝酸 D 精准修复溶酶体,从根源延缓血管衰老
ACS Nano(IF 16)| 基于天然葛根的树蛙足导电性心脏贴片,为心肌梗死治疗开辟中药新材料赛道
AFM(IF=19.0)|中医 “君臣佐使” 指导天然药碳点化,重编程肿瘤死亡、突破胶原屏障并激活抗肿瘤免疫
Adv Sci(IF14.1)重磅:三叶崖爬藤多糖破局三阴性乳腺癌,靶向关键通路阻断肿瘤转移
中风治疗重大突破!协和团队顶刊揭秘:人参皂苷 Rg1 “钓” 出关键靶点,三七护脑机制终于说清
深度解读 | 中国药科大学 藏红花 Crocin II “精准降解” ANGPTL8: 一场天然小分子对 MASLD
美国心脏协会重磅发布|AHA 2026心血管膳食指南:告别“单一控食”,护心饮食迎来全新革命
本草学术观察 | 祛湿不伤正,健脾不滋腻 —— 茯苓,最温和的本草养生
食药同源的“祛湿圣品”:茯苓,藏在日常里的健脾养身良方
本草学术观察 | 养血调经第一药:当归,藏在岁月里的“女人参”
本草学术观察 | 补气固表第一药:真正的平民补气之王
本草学术观察 | 红枣:补气养血第一果,吃对才养人
跳出 “偏方” 误区,用科学视角解读 3 种常见药食同源食材
2026-04-24
吴姝静吕玟玟叶瑞萍罗玉珍孟宇翔谢晓莹姜明国 周燕
(广西民族大学 海洋与生物技术学院,广西多糖材料与改性重点实验室,广西 南宁)
DOI:10.13343/j.cnki.wsxb.20250913
摘 要
目的筛选兼具广谱拮抗活性和水果采后保鲜功能的微生物源挥发性有机物(volatile organic compounds, VOCs)产生菌株。
方法以广西野生茶树根系和枝条内生细菌为研究对象,采用稀释涂布平板法分离纯化内生细菌。通过纤维素酶、淀粉酶、铁载体、有机磷、无机磷及固氮等6类功能培养基进行筛选,依据功能特性数量初步筛选候选菌株;采用平板对峙法测定菌株对7种常见通用植物病原真菌的拮抗活性,筛选广谱拮抗菌株。进一步选取综合性能较优的菌株,针对芒果和芭蕉炭疽病原菌——果生炭疽菌(Colletotrichum fructicola)和芭蕉炭疽菌(Colletotrichum musae)进行拮抗评价,并以化学保鲜剂咪鲜胺为对照开展离体香蕉保鲜试验。
结果功能培养基初筛共获得98株同时具备固氮、溶磷、产铁载体等4种及以上促生或抗逆功能特性的菌株。以7种常见植物病原真菌为靶标进行平板对峙复筛得到18株对至少5种病原菌具有显著抑制作用的广谱拮抗菌株,其中4株对全部7种病原菌均表现出稳定且较强的拮抗活性。在此基础上,综合考量功能特性数量和广谱抑菌率2项核心指标,最终遴选出5株综合性能最优的菌株,用于后续针对芒果/芭蕉炭疽病原菌的特异性拮抗评价及果实保鲜效果验证。平板对峙试验中5株菌株对2种炭疽病原菌的抑菌率为43.36%-83.50%;平板对扣试验显示,其产生的VOCs对果生炭疽菌和芭蕉炭疽菌的抑菌率分别为56.80%-99.25%和54.50%-99.85%,部分菌株对2种病原菌均表现出接近100.00%的强抑菌活性。离体果实保鲜试验结果表明,拮抗菌株产生的VOCs均能不同程度延缓芒果和香蕉的腐烂进程,其中菌株T-1-6保鲜效果最为突出,可将香蕉明显腐烂时间延长至21 d,末期腐烂等级为0级,其保鲜效果与化学保鲜剂咪鲜胺相当,对芒果表面霉菌的防效约为50%。16S rRNA基因和gyrB基因序列分析表明,5株优势拮抗菌株均隶属于芽孢杆菌属(Bacillus),包括解淀粉芽孢杆菌(Bacillus amyloliquefaciens)、苏云金芽孢杆菌(Bacillus thuringiensis)、蜡样芽孢杆菌(Bacillus cereus)和枯草芽孢杆菌(Bacillus subtilis)等。
结论本研究从茶树根系和枝条内生细菌中获得了一批具有多功能特性和广谱拮抗活性的VOCS产生菌株,为芒果、香蕉等热带水果采后病害的绿色防控与生物保鲜技术开发提供了潜在优良菌种资源和理论依据。
关键词 茶树内生细菌;芽孢杆菌;挥发性有机物;炭疽病;水果保鲜
近年来,我国水果产量持续增长,但果蔬产后损耗率仍然居高不下。为延长水果货架期、减少产后损耗,化学保鲜剂(如丙二醇、山梨酸钾、二氧化硫等)在果蔬贮藏中应用广泛[1-4]。然而,长期或不当使用化学保鲜剂不仅容易改变水果风味,部分成分残留还可能引发食品安全隐患,并对贮藏环境造成污染[5]。因此,开发天然、安全且高效的保鲜技术与保鲜因子,逐渐成为水果保鲜领域的研究热点。其中,微生物源挥发性有机物(volatile organic compounds, VOCs)因具有无直接接触的作用方式、易降解、不易在果实表面形成明显残留等优势,被视为替代化学保鲜剂的重要发展方向[6]。
在各类产VOCs微生物中,细菌因分布广泛、抗逆性强、易于规模化制备且代谢产物类型丰富,在基于VOCs的果蔬保鲜研究中占据重要地位[7]。已有研究表明,部分植物根际促生菌及动物肠道来源的生防芽孢杆菌具有较广的抑菌谱,对多种病原菌均表现出显著抑制作用[8-10]。这类细菌不仅可以产生抗生素、拮抗蛋白等拮抗物质,还可通过调控果蔬呼吸作用、乙烯释放及抗氧化体系等生理代谢过程,延缓果实软化和腐烂,减缓品质劣变[11]。目前,用于植物病害生物防治及果蔬保鲜研究较多的菌属主要包括芽孢杆菌属(Bacillus)、假单胞菌属(Pseudomonas)、产乳酸菌属(Lacticigenium)、肠杆菌属(Enterobacter)、放线菌属(Actinomyces)等[12-17]。例如,钟涛[18]发现荧光假单胞菌(Pseudomonas fluorescens) ZX产生的VOCs中乙酸、异丁酸、二甲基二硫和二甲基三硫等成分可破坏病原真菌细胞膜完整性并诱导活性氧(reactive oxygen species, ROS)积累,从而实现延长果实货架期的效果;Calvo等[19]报道的3株芽孢杆菌(BUZ-14、I3和I5)产生的VOCs对灰葡萄孢菌(Botrytis cinerea)、柑橘青霉病菌(Penicillium italicum)、指状青霉(Penicillium digitatum)均具有较高抑菌率,经气相色谱-质谱联用(GC-MS)分析共鉴定到12-15种主要VOCs成分,包括2-壬酮、2-庚酮、苯甲醛、壬烷等。
细菌VOCs的保鲜效果与菌株种类及代谢特性密切相关,不同类群细菌产生的VOCs在成分类型(如萜类、碱类、酮类等)及含量上存在显著差异,进而影响其对果蔬的保鲜活性和适用范围[20-21]。左庚鑫[22]从金银花中分离得到暹罗芽孢杆菌(Bacillus siamensis) NEAU-ZGX24,其产生的VOCs对灰葡萄孢菌的抑制率可达100%,不仅可显著延缓番茄采后自然腐烂过程,对其他多种植物病原真菌也表现出广谱抑菌活性。GC-MS结果显示,该菌株可产生戊酸、3-甲基丁酸和庚酸乙酯等14种挥发性有机化合物,其中部分成分为抗B. cinerea的关键活性物质。
广西地处亚热带季风气候区,光热资源充足、雨量丰沛,微生物多样性丰富,是重要的热带、亚热带水果产区之一。然而,目前在当地生产实际中生物保鲜剂的推广应用仍较为有限,针对广西特色水果(如香蕉、芒果等)适配的广谱微生物源VOCs筛选研究也相对较少[23]。在香蕉、芒果采后贮藏环节,炭疽病等真菌性病害高发,是造成果实腐烂和经济损失的重要因素。因而,从本地微生物资源中挖掘对该类病原菌具有高效拮抗和保鲜活性的VOCs产生菌株,阐明其对特色水果的保鲜作用,为构建安全、绿色、高效的地方化生物保鲜技术体系提供菌种基础和理论依据具有重要意义。
本研究以广西典型特色水果芒果和香蕉为研究对象,从当地茶树健康枝条内分离获得的内生细菌中筛选具有多功能特性和广谱拮抗活性的菌株,重点评价其产生的VOCs对芒果、芭蕉炭疽病原真菌的抑制效果及离体果实保鲜能力,并对优势菌株进行形态学和分子生物学鉴定,以期获得兼具广谱抑菌活性和水果保鲜功能的微生物源VOCs产生菌株,为开发适用于广西及类似生态区热带水果采后病害综合防控和保鲜的生物制剂提供理论与技术支撑。
1 材料与方法
1.1 材料
1.1.1 茶树枝条样品与内生细菌来源
试验菌株来源于广西壮族自治区桂林市平乐县青龙乡大冈村(24°27′02.6′′N,110°49′04.0′′E)野生石崖茶树根系和枝条。采样时先剔除地表落叶、杂草等杂物,用灭菌铲挖去表层2-3 cm土壤后,采集5-10 cm土层范围内的茶树根系样品,置于无菌采样袋中密封;同时采集健壮茶树枝条,及时送回实验室,于4 ℃冷藏保存,备用。
1.1.2 病原真菌
小麦平稷蠕孢菌(Bipolaris sorokiniana) BS、新暗色柱节孢菌(Neoscytalidium dimidiatum) H8、瓜笄霉(Choanephora cucurbitarum) H4、板栗疫病菌(Cryphonectria parasitica) EP155、尖刀镰孢菌(Fusarium oxysporum) FOCO、厚垣镰孢菌(Fusarium chlamydosporum) B2、葡萄座腔菌(Botryosphaeria dothidea) BZ8、果生炭疽菌(Colletotrichum fructicola) M9和芭蕉炭疽菌(Colletotrichum musae) XJTJ,由广西高校微生物与植物资源利用重点实验室保存待用。
1.1.3 供试水果
供试芒果选用桂七芒果,采自广西壮族自治区亚热带作物研究所芒果园,香蕉为商品成熟前期采收的本地栽培品种。试验水果要求大小一致、成熟度相近、表皮颜色均匀、表面无机械伤和明显病斑。
1.1.4 培养基与主要试剂
细菌培养采用LB培养基,病原真菌培养采用PDA培养基。菌株功能性检测分别使用阿须贝氏无氮(ashby nitrogen-free, ANF)培养基、蒙金娜有机磷(meng jinna organic phosphorus, MJOP)培养基、Pikovskaya 无机磷(Pikovskaya inorganic phosphorus, PIP)培养基、铁载体(chrome azurol S, CAS)检测培养基、纤维素酶检测(cellulase detection, CD)培养基和淀粉酶检测(amylase detection, AD)培养基等。各培养基均购自青岛海博生物技术有限公司。纤维素酶活性检测采用刚果红溶液,淀粉酶活性检测采用碘伏溶液,其余试剂均为分析纯或分析纯以上级别。所有试剂均采自上海麦克林生化科技股份有限公司。
LB培养基(g/L):胰蛋白胨10.0,酵母浸粉5.0,NaCl 10.0。
PDA培养基(g/L):马铃薯提取物200.0,葡萄糖20.0,琼脂18.0。
阿须贝氏无氮培养基(g/L):甘露醇10.0,KH2PO4 0.2,MgSO4·7H2O 0.2,NaCl 0.2,CaSO4 0.1,CaCO3 5.0,琼脂18.0。
蒙金娜有机磷培养基(g/L):葡萄糖10.0,(NH4)2SO4 0.5,酵母浸粉0.5,NaCl 0.3,KCl 0.3,MgSO4 0.3,FeSO4 0.03,MnSO4 0.03,卵磷脂0.2,CaCO3 1.0,琼脂18.0。
Pikovskaya无机磷培养基(g/L):葡萄糖10.0,酵母浸粉0.5,(NH4)2SO4 0.5,NaCl 0.3,KCl 0.3,MgSO4 0.3,FeSO4 0.03,MnSO4 0.03,Ca3(PO4)2 5.0,琼脂18.0。
铁载体检测培养基(g/L):CAS 0.06,HDTMA 0.0723,FeCl3·6H2O 0.002,NaH2PO4·2H2O 0.295,Na2HPO4·12H2O 1.214,NH4Cl 0.125,KH2PO4 0.037,NaCl 0.063,琼脂9.0。
纤维素酶检测培养基(g/L):胰蛋白胨10.0,酵母浸粉5.0,NaCl 10.0,羧甲基纤维素钠5.0。
淀粉酶检测培养基(g/L):胰蛋白胨10.0,酵母浸粉5.0,NaCl 10.0,可溶性淀粉2.0。
1.2 菌株的分离纯化
为尽可能去除样品表面微生物,获得植物内生菌,首先将茶树根系、枝条样品用自来水充分冲洗约30 min,初步去除根表面附着的土壤和杂质,再用无菌滤纸吸干表面水分。随后将根系、枝条样品置于质量分数为1%的次氯酸钠溶液中浸泡消毒2次,每次2 min;再转入体积分数为75%的乙醇溶液中进行表面消毒2次,每次1 min。消毒后用无菌蒸馏水冲洗3-4次以去除残留消毒剂,用无菌滤纸轻轻吸干水分。
在超净工作台(浙江苏净净化设备有限公司)内,用灭菌剪刀将根系样品剪碎或用刀将枝条、根系样品切成薄片后,置于无菌研钵中,加入适量无菌水研磨成均匀匀浆。按10-3、10-4、10-5 3个稀释度分别进行10倍梯度稀释,取各稀释度稀释液100 μL滴加于LB平板上,用无菌涂布棒均匀涂布,每个稀释度设置3个平板作为平行重复。将最后一次冲洗液100 μL涂布于LB平板,作为表面消毒对照,以验证表面消毒是否彻底。所有平板置于30 ℃恒温培养箱(上海智城分析仪器制造有限公司)中培养约24 h,观察菌落生长情况。随后根据菌落形态(大小、颜色、边缘形状、表面质地等)以及与表面消毒对照比对挑取与表面消毒对照不同表型的单菌落,转接至新的LB固体培养基上进行三区划线纯化,重复转接直至获得纯净菌株,编号并甘油保藏备用。
1.3 拮抗菌株的筛选
1.3.1 菌株的功能性测试
按相应配方分别制备纤维素酶、淀粉酶、CAS、含有机磷、含无机磷及固氮培养基。除固氮培养基外,其余培养基均采用点种方式:挑取新鲜单菌落接种于平板中央;固氮培养基则采用划线接种方式,观察菌株在无氮条件下的生长能力。
接种后,将各平板置于30 ℃恒温培养箱中培养2-3 d。对于有机磷、无机磷和CAS培养基,可直接观察菌落周围是否形成透明圈或显色圈,判断其溶磷或产铁载体等功能。固氮培养基仅需观察菌落是否能够正常生长。
纤维素酶和淀粉酶活性检测需经染色显色后观察透明圈。纤维素酶培养基培养2-3 d后打开平板,加入约1 mL、质量浓度为1 mg/mL的刚果红溶液染色10 min,倒去染液后再加入0.9%的氯化钠溶液洗涤脱色约10 min,观察菌落周围是否形成明显水解圈。淀粉酶培养基则用碘伏溶液染色约1 min,观察菌落周围淀粉水解形成的无色透明圈。
每株菌在各类功能培养基上均设置3个平行重复。以水解圈或透明圈直径(D)与菌落直径(d)的比值D/d作为功能强度指标[24-25]。综合各项功能指标,筛选出具备多种功能特性的候选菌株。
1.3.2 菌株的广谱抑菌性测试
采用平板对峙法检测待测菌株对7种实验室保存的植物病原真菌的抑制作用。将预先在PDA平板上活化培养的病原真菌,用打孔器于菌落边缘取直径约5 mm的菌饼,接种于新鲜PDA平板中央。再取待测菌株发酵液(OD600≈1.0) 5 μL,分别滴加于距菌饼中心约2.5 cm处的上下左右4个位置,呈十字分布。仅接种病原真菌而不接种细菌的PDA平板作为对照。
所有平板置于30 ℃恒温培养箱中黑暗培养5-7 d,每个处理设置3个平行重复。当对照组病原真菌菌落基本长满平板或达到稳定生长状态时测量病原菌沿指向菌株方向的菌落半径,计算抑菌率,如公式(1)所示。
抑菌率=(对照组菌落半径-处理组菌落半径)/
对照组菌落半径×100%(1)
其中,对照组菌落半径为不接种拮抗菌条件下病原菌的生长半径,处理组菌落半径为对峙培养条件下病原菌的生长半径。根据抑菌率大小评价各菌株的广谱抑菌能力,并筛选对多种病原真菌均有较强抑制作用的菌株。
1.3.3 菌株对芒果、芭蕉炭疽病常见病原菌的拮抗测试
采用平板对峙法进一步检测待测菌株对芒果、芭蕉炭疽病原菌的抑制作用,病原菌分别为果生炭疽菌和芭蕉炭疽菌。具体方法同1.3.2节,分别在PDA平板中央接种2种炭疽病原菌菌饼,四周滴加待测菌株发酵液,对照组仅接种病原菌不接种细菌。前期通过光照与暗培养的多组预实验发现,本研究中针对香蕉、芒果炭疽病原菌的拮抗实验体系,在黑暗条件下的菌落生长均一性及抑菌结果重现性更优,因此本研究统一采用30 ℃黑暗培养5-7 d的条件,测量菌落半径并计算抑菌率,评价菌株对香蕉、芒果炭疽病原菌的对峙拮抗作用。
同时采用平板对扣法测定待测菌株产生的VOCs对2种炭疽病原菌的抑制作用。将直径约5 mm的病原真菌菌饼接种于PDA平板中央;另取待测菌株发酵液100 μL均匀涂布于LB平板表面,以未涂布菌液的LB平板为空白对照。将接种病原真菌的PDA平板与接种细菌的LB平板反向对扣(菌面相对),用封口膜将2平板边缘严密封口,以减少VOCs外逸。每个处理设置3个平行重复。所有对扣平板置于30 ℃恒温培养箱中黑暗培养7 d,结束培养后测量病原真菌菌落半径,按公式(1)计算抑菌率,从而评价待测菌株VOCs对芒果、芭蕉炭疽病原菌的挥发性抑制效果。
1.4 拮抗菌株的初步鉴定
1.4.1 菌株形态特征观察
将拮抗效果显著的优势菌株接种于LB固体培养基上进行三区划线纯化,30 ℃培养24 h后观察单菌落形态特征,包括菌落大小、颜色、边缘形状、表面光滑度及透明度等。挑取单菌落制作涂片,采用革兰氏染色法染色,经光学显微镜(奥林巴斯公司)观察细胞形态(如细胞形状、排列方式等)及革兰氏反应结果,对菌株进行形态学初步鉴定。
1.4.2 分子生物学鉴定
以优势拮抗菌株的基因组DNA为模板,采用16S rRNA基因通用引物27F (5′-AGAGTTT GATCCTGGCTCAG-3′)和1492R (5′-TACGGC TACCTTGTTACGAGTT-3′),以及管家基因gyrB的引物UP-1 (5′-ATTTGGCGCTGGCGGT TAT-3′)和UP-2r (5′-GGTTTCGGCTGGGCTGG TA-3′)进行PCR扩增。挑取新鲜单菌落菌株至100 μL的10% Chelex100中,煮沸30 min,12 000 r/min离心2 min,提取上清液作为模板。
PCR反应体系(50 μL):2×Taq PCR Mix 25 μL,DNA模板1 μL,上、下游引物(0.2 μmol/L)各1 μL,ddH2O 22 μL。PCR反应条件:95 ℃预变性5 min;95 ℃变性1 min,55 ℃退火1 min,72 ℃延伸2 min,共32个循环;72 ℃终延伸10 min;4 ℃保存。扩增产物经琼脂糖凝胶电泳鉴定条带大小及特异性后,送生工生物工程(上海)股份有限公司进行测序。将获得的16S rRNA基因序列和gyrB基因序列在NCBI数据库中进行BLAST比对,结合系统发育树分析结果确定菌株的种属归属。部分代表性菌株的16S rRNA基因和gyrB基因序列提交至GenBank获取登录号,以备后续引用和溯源。
1.5 水果保鲜试验
采用离体香蕉和芒果试验评价拮抗菌株VOCs对香蕉、芒果采后病害的防控效果及保鲜作用,同时以化学保鲜剂咪鲜胺作为阳性对照组进行效果对比。选取同一品种、大小均匀、成熟度相近且表面无机械伤和明显病斑的水果作为供试材料。保鲜装置采用密闭性良好的塑料保鲜盒(约40 cm×25 cm×20 cm),拮抗菌处理采用相应菌株菌液涂布的LB平板作为VOCs释放源。
将挑选好的水果随机分组,每组3个果实。处理组将水果置于保鲜盒中,并放入预先涂布有拮抗菌株菌液的LB平板,同时放置不含水的无菌棉球以调节湿度;对照组保鲜盒中放入未接种菌液的LB平板,也放置无菌棉球,保持盒内湿度条件与处理组一致。所有保鲜盒置于实验室内避光静置,避免阳光直射。实验室配有基础空调,环境条件较为稳定,温度维持在24-26 ℃,相对湿度为60%-70%,以减少保鲜盒内冷凝水形成或样品过度失水。
保鲜过程中定期观察水果果皮颜色变化、果肉软化程度及有无异味产生,并对腐烂部位及其扩展情况进行拍照记录。根据果皮病斑面积和腐烂程度对果实进行分级评价,记录各处理组的腐烂果数及腐烂程度等指标,必要时可计算腐烂率和腐烂指数等参数。本研究采用果蔬炭疽病通用分级方法,共设0/1/3/5/7/9级,以病斑面积占果皮总面积的比例为核心判定指标,其中0级为无病,1级病斑占比≤10%,3级占比11%-25%,5级占比26%-50%,7级占比51%-75%,9级占比76%以上[26-27]。整个试验持续观察7-21 d。在相同条件下进行3次独立、完整的生物学重复实验。通过比较不同处理组水果腐烂速度、腐烂程度和外观品质变化,综合评估各拮抗菌株VOCs对香蕉和芒果的保鲜效果。实验过程中尽量保持实验室环境的相对稳定,减少外界干扰因素对试验结果的影响。
2 结果与分析
2.1 菌株的功能特性及菌株的广谱抑菌性
2.1.1 菌株的解磷、产铁载体、产酶和固氮能力
以解磷、产铁载体、胞外酶产生及固氮等特性为指标,对202株茶树根系和枝条内生细菌进行功能筛选。统计结果表明,具有4种及以上功能特性的菌株共98株,各菌株所具备功能数的分布情况见图1。
图1 菌株功能数量分布Figure 1 Distribution of the number of functional traits among bacterial strains.
在溶磷能力检测中有89株菌株能够在有机磷培养基上形成明显水解圈,表现出溶解有机磷的能力;97株菌株在无机磷培养基上形成透明圈,具有溶解无机磷的能力。在CAS检测培养基上,共有112株菌株在菌落周围产生颜色由蓝色向浅色转变的透明圈,其中23株菌株形成橙黄色透明圈,表明其产铁载体能力较强。胞外酶活性测定结果显示,44株菌株可产生纤维素酶,在纤维素培养基上经刚果红染色后形成明显水解圈;66株菌株可产生淀粉酶,在淀粉培养基上经碘伏显色后形成无色透明圈。在阿须贝氏无氮培养基上,有126株菌株能够正常生长,推测其具有一定固氮能力(图2)。部分代表性功能菌株在不同功能培养基上的表现见图3,部分菌株D/d值统计结果见表1。
图2 具有不同功能特性的菌株数量统计Figure 2 Number of strains exhibiting different functional traits.
图3 部分菌株在不同功能培养基上的实验效果Figure 3 Functional characteristics of representative strains on different selective media. A: Organic phosphorus solubilization; B: Inorganic phosphorus solubilization; C: Siderophore production; D: Amylase production; E: Nitrogen fixation; F: Cellulase production.
表1 部分菌株在不同功能培养基上的活性强度Table 1 Functional activity of representative strains on different selective media (D/d)
综合各项功能指标,表明从茶树根系和枝条分离获得的内生细菌具有较为丰富的功能多样性,为进一步筛选兼具拮抗活性和保鲜潜力的优势菌株提供了良好的菌种基础。
2.1.2 菌株拮抗性能分析
在筛选获得的202株菌株中分别统计其对小麦平稷蠕孢菌、新暗色柱节孢菌、瓜笄霉、板栗疫病菌、尖刀镰孢菌、厚垣镰孢菌和葡萄座腔菌7种常见通用植物病原真菌的抑制效果。结果表明,对上述病原真菌抑菌率达到50%以上的菌株数量分别为51、60、47、54、33、46和40株(图4)。其中,对5种及以上病原真菌均具有明显抑制作用的菌株共有18株,对7种病原真菌均表现出拮抗活性的菌株有4株,显示出较好的广谱拮抗潜力(图5)。
图4 针对不同病原真菌拮抗菌株数量Figure 4 Number of antagonistic strains against different plant pathogenic fungi.
图5 菌株对病原真菌的拮抗谱统计Figure 5 Antagonistic spectrum of the strains against plant pathogenic fungi.
在具备广谱拮抗能力的4株菌株中,菌株T-1-6对7种病原真菌均表现出较强的拮抗效果,平板对峙培养可见菌落间形成清晰的抑菌带,病原真菌生长受到明显抑制(图6)。其对不同病原真菌的抑菌率见表2。
图6 菌株T-1-6对植物病原真菌的拮抗效果Figure 6 Antagonistic effects of strain T-1-6 against plant pathogenic fungi. A: Bipolaris sorokiniana; B: Neoscytalidium dimidiatum; C: Choanephora cucurbitarum; D: Cryphonectria parasitica; E: Fusarium oxysporum; F: Fusarium chlamydosporum; G: Botryosphaeria dothidea.
表2 T-1-6对7种常见植物病原真菌的抑菌效果Table 2 Inhibitory effects of strain T-1-6 against seven plant pathogenic fungi
The data in the table are mean±SD; Different lowercase letters indicate significant differences (P<0.05) as tested by one-way ANOVA method.
2.2 菌株对芒果、芭蕉炭疽病常见病原菌的平板对峙实验
从前期筛选出的拮抗谱较广且功能特性较优的菌株中选取8株拮抗细菌,采用平板对峙法检测其对芒果、芭蕉炭疽病原菌的抑制作用,病原菌分别为果生炭疽菌和芭蕉炭疽菌。结果表明,8株菌株对2种炭疽病原真菌均表现出不同程度的拮抗活性,抑菌率总体分布在43.36%-83.50%之间(其中菌株T-1-6对果生炭疽菌的抑菌率最高,达83.50%)。部分菌株对果生炭疽菌和芭蕉炭疽菌的抑菌结果见表3。
表3 菌株对果生炭疽菌和芭蕉炭疽菌的抑菌效果Table 3 Inhibitory effects of strains against Colletotrichum fructicola and Colletotrichum musae (%)
The data in the table are mean±SD; Different lowercase letters indicate significant differences (P<0.05) as tested by one-way ANOVA method.
多株菌株对至少一种炭疽病原真菌的抑菌率超过55.00%,部分平板上可见病原菌菌落生长受限、抑菌圈清晰且范围较大,说明这些菌株对芒果、芭蕉炭疽病原菌具有较强的抑制能力。图7、图8分别为菌株T-1-6对果生炭疽菌和芭蕉炭疽菌的平板对峙效果,与空白对照组(CK)相比,菌株T-1-6可显著抑制果生炭疽菌、芭蕉炭疽菌的菌落扩展,病原菌生长受抑明显,抑菌圈清晰且范围较大,说明该菌株对两种炭疽病原菌的有较强抑制能力,可作为芒果、芭蕉炭疽病生物防治的潜在菌株。
图7 T-1-6对果生炭疽菌的抑制作用Figure 7 Inhibitory effect of strain T-1-6 against Colletotrichum fructicola. A: CK; B: T-1-6.
图8 T-1-6对芭蕉炭疽菌的抑制作用Figure 8 Inhibitory effect of strain T-1-6 against Colletotrichum musae. A: CK; B: T-1-6.
2.3 菌株VOCs对芒果、芭蕉炭疽病常见病原菌的拮抗测试
采用平板对扣法检测拮抗菌株产生的VOCs对果生炭疽菌和芭蕉炭疽菌的抑制作用。结果表明,各菌株产生的挥发性物质对2种病原真菌均表现出不同程度的抑制效果。对果生炭疽菌的抑菌率介于56.80%-99.25%,对芭蕉炭疽菌的抑菌率介于54.50%-99.85%,其中部分菌株对2种病原菌的抑菌率均接近100.00%,表现出极强的挥发性抑菌活性,多数菌株对至少一种病原菌的抑菌率超过55.00% (表4)。
表4 菌株VOCs对果生炭疽菌和芭蕉炭疽菌的抑菌效果Table 4 Inhibitory effects of VOCs produced by strains against Colletotrichum fructicola and Colletotrichum musae (%)
The data in the table are mean±SD; Different lowercase letters indicate significant differences (P<0.05) as tested by one-way ANOVA method.
菌株T-1-6产生的VOCs对芒果炭疽病原菌的抑制效果见图9,对芭蕉炭疽病原菌的抑制效果见图10。与对照相比,处理组病原菌菌落直径明显减小,菌落边缘生长受限,表明VOCs是该菌株重要的拮抗因子之一。部分菌株VOCs对果生炭疽菌和芭蕉炭疽菌的抑菌结果见表4,各菌株抑菌率数据显示,试供菌株VOCs对果生炭疽菌的抑菌率整体分布于56.80%-99.25%,对芭蕉炭疽菌的抑菌率整体分布于54.50%-99.85%,且多数菌株对两种病原菌的抑菌率均在70%以上。菌株T-1-6 VOCs对果生炭疽菌的抑制作用见图9,对芭蕉炭疽菌的抑制作用见图10。
图9 T-1-6 VOCs对果生炭疽菌的抑制作用Figure 9 Inhibitory effect of VOCs produced by strain T-1-6 against Colletotrichum fructicola. A: CK; B: T-1-6.
图10 T-1-6 VOCs对芭蕉炭疽菌的抑制作用Figure 10 Inhibitory effect of VOCs produced by strain T-1-6 against Colletotrichum musae. A: CK; B: T-1-6.
2.4 拮抗菌株的鉴定
根据菌株对芒果、芭蕉炭疽病原菌的拮抗试验结果,选取综合性能较优的T-1-6、T-1-11、T-2-3、T-3-8、T-3-9共5株菌株进行进一步鉴定。以其基因组DNA为模板扩增16S rRNA基因和gyrB基因,测序后在NCBI数据库中进行BLAST比对分析,结合同源序列的系统发育关系,明确其分类学地位。
鉴定结果(表5)显示,5株拮抗菌株均隶属于芽孢杆菌属(Bacillus)。其中,T-1-6为解淀粉芽孢杆菌(Bacillus amyloliquefaciens),T-1-11和T-3-8为苏云金芽孢杆菌(Bacillus thuringiensis),T-2-3为蜡样芽孢杆菌(Bacillus cereus),T-3-9为枯草芽孢杆菌(Bacillus subtilis)。上述结果表明,从茶树内生菌中筛选获得的优势拮抗菌株主要集中于典型的生防芽孢杆菌类群。系统发育分析结果显示菌株T-1-6的16S rRNA基因序列与解淀粉芽孢杆菌聚集于同一进化分支(图11),gyrB序列比对结果发现与解淀粉芽孢杆菌相似度最高,且系统发育树聚在同一支(图12)。
表5 五株菌的分子生物学鉴定结果Table 5 The molecular biology identification results of five strains
图11 菌株T-1-6基于16S rRNA基因序列的系统发育树Figure 11 Phylogenetic tree of strain T-1-6 based on 16S rRNA gene sequence.
图12 菌株T-1-6基于gyrB基因序列的系统发育树Figure 12 Phylogenetic tree of strain T-1-6 based on gyrB gene sequence.
2.5 水果保鲜试验
综合菌株功能特性和拮抗谱结果,选取上述5株性能较优的拮抗菌株开展离体水果保鲜试验,评价其对香蕉和芒果的保鲜能力,并以化学保鲜剂咪鲜胺为阳性对照进行效果对比。与对照组相比,所有拮抗菌株VOCs处理组和咪鲜胺对照组在芒果和香蕉的贮藏过程中均表现出一定程度的保鲜效果,主要体现在:果实腐烂发生时间推迟、腐烂率较低、果肉软化速度减缓等方面。
在室温条件下,随着贮藏时间延长,对照组香蕉和芒果逐渐出现果皮变黄、黑斑扩展、果肉软烂及霉菌滋生等现象,而多数菌株VOCs处理组上述变化发生明显滞后,腐烂进程相对缓慢,不同菌株之间的保鲜效果存在一定差异。整体来看,大部分处理组果面霉斑扩展速度和腐烂速度均显著低于对照组,说明拮抗菌株产生的VOCs对延缓果实衰老发挥积极作用。
其中,菌株T-1-6的保鲜效果最为突出。在芒果保鲜试验中,T-1-6 VOCs处理组果面病斑较少、霉层形成明显减轻,对芒果表面霉菌的防效约为50% (图13),保鲜效果显著优于对照组。在香蕉保鲜试验中,T-1-6 VOCs处理可将香蕉明显腐烂时间延长至21 d,终止观察时腐烂等级为0,其保鲜效果与咪鲜胺相当(图14);综合分析推测,菌株VOCs保鲜原理与咪鲜胺作用类似,也可能通过抑制病原菌暴发等机制实现水果绿色保鲜,为水果贮藏提供了具有应用潜力的生物保鲜技术方向。
图13 菌株T-1-6 VOCs对芒果采后保鲜效果Figure 13 Postharvest preservation effect of VOCs produced by strain T-1-6 on mango fruits. The figure shows the result of one representative experiment.
图14 菌株T-1-6 VOCs对香蕉采后保鲜效果Figure 14 Postharvest preservation effect of VOCs produced by strain T-1-6 on banana fruits. The figure shows the result of one representative experiment.
3 讨论与结论
植物内生菌广泛存在于植物体内,与宿主植物形成相对稳定的共生体系。宿主植物为内生菌提供营养和栖息环境[28],内生菌则通过帮助宿主吸收氮、磷、钾等必需营养元素促进多种酶系活化[29],进而影响植物生长发育和次生代谢物质积累。例如,杨金新等[30]从水稻叶片中分离得到的菌株JKDY-3兼具解钾、溶磷、产铁载体、分泌吲哚-3-乙酸(indole-3-acetic acid, IAA)和固氮等多种功能,显著促进水稻幼苗根系生长;李锦等[31]从威廉斯香蕉抗枯萎病株系根、球茎及假茎中分离到的变栖克雷伯氏菌(Klebsiella variicola) XQJ0301具有产生生长素、解钾、产铁载体和产纤维素酶能力,单独施用时对根长、株高、根体积和假茎周长等指标的促进效果均优于其他处理;刘天金等[32]从巨菌草茎中分离到一株耐盐碱内生菌,具备解磷、解硅酸盐、固氮和产铁载体等促生特性,在玉米和番茄盆栽试验中表现出一定促生作用。上述研究表明,内生菌的分离鉴定及功能菌筛选不仅有助于解析植物促生机制,也为农业生产提供了潜在的生防和促生菌资源。功能特性丰富的菌株通常同时具有杀虫、抑菌等多重应用潜力,因此开展多功能内生菌的系统筛选具有重要研究价值。
本研究以茶树根系和枝条内生细菌为对象,通过筛选具备基础生理代谢能力和多种功能特性的菌株,初步淘汰代谢能力较弱、定殖潜力有限的菌株,从而获得多功能候选菌株,为后续用于常见水果病原菌防治和果实保鲜奠定基础。结果显示,从茶树根系和枝条共分离获得202株内生细菌,其中98株同时具备4种及以上功能特性;在功能性测试中具有溶解有机磷能力的菌株有89株,具有溶解无机磷能力的有97株,能产生铁载体的有112株,可产纤维素酶的有44株,可产淀粉酶的有66株,能在无氮条件下正常生长、具有固氮能力的有126株。这表明茶树根系和枝条内生细菌群体具有较高的功能多样性和代谢潜力,为进一步挖掘其促生、杀虫和抑菌等综合应用价值提供了物质基础。后续可通过盆栽或田间试验,系统评价代表性菌株在茶树等作物上的促生、抗逆和潜在杀虫效果,对茶树根系和枝条内生细菌资源进行更深入开发。
基于功能板试验结果及已有研究报道可对本研究筛选到的拮抗菌株抑菌机制作出一定推测。(1) 部分菌株能够合成并分泌铁载体等小分子配体,高效螯合环境中的游离铁离子形成病原菌难以利用的稳定络合物,从而在铁等关键营养元素竞争中占据优势,抑制病原菌的生长繁殖。(2) 多数植物病原真菌的细胞壁以纤维素和几丁质为主要成分,产纤维素酶等胞外酶的菌株可能通过分解或破坏病原菌细胞壁结构,干扰其正常代谢过程,从而抑制病原真菌的生长[33]。这些推测性机制与本研究中解淀粉芽孢杆菌等优势菌株所表现出的溶磷、产铁载体和产纤维素酶等功能特性相吻合,但其具体作用途径仍有待后续通过分子生物学和生化水平的实验进一步证实。
近年来,国内外学者对生物防治的关注度不断提升,大量研究表明,自然环境中广泛存在的生防细菌对多种植物病原菌具有显著拮抗作用。侯会霞等[34]从剑麻、火龙果和橡胶树等作物组织中分离得到的菌株PpHyHNCJ2和PpHyHNCJ5能广谱抑制辣椒疫霉菌(Phytophthora capsica)、大豆疫霉菌(Phytophthora sojae)、瓜疫霉菌(Phytophthora melonis)和棕榈疫霉菌(Phytophthora palmivora)等11种病原菌生长;丁艳哲等[35]从人工栽培刺五加根际土壤中获得的菌株YZ-375对人参立枯丝核菌(Rhizoctonia solani)、人参链格孢菌(Alternaria panax)、人参茄腐皮镰孢菌(Fusarium solani)、人参毁灭柱孢菌(Cylindrocarpon destructans)及五味子尖镰孢菌(Fusarium oxysporum)均表现出拮抗活性,抑菌率为69.21%-77.44%。本研究通过对7种常见通用植物病原真菌和果生炭疽菌、芭蕉炭疽菌2种芒果、芭蕉炭疽病原菌的拮抗筛选,从茶树内生菌中获得了5株对上述9种植物病原真菌均具有较强抑制作用的拮抗菌株,表现出良好的广谱拮抗活性。在本研究设定的模拟贮藏条件下,这些菌株的生长表型及其对水果腐烂进程的抑制效果均较为稳定,使用方式相对简便,具有一定的应用潜力。上述拮抗菌株不仅为多种植物病害的生物防治提供了新的候选菌株,也为水果保鲜技术开发及后续应用推广提供了重要的菌种基础,为水果保鲜应用提供了具备商业推广价值的菌种支撑。需要指出的是,本研究尚未深入解析这些拮抗菌株对病原真菌的具体抑菌机制,也未开展与其他高效菌株的复配防治策略研究,后续可围绕拮抗物质类型、诱导抗性及菌株间互作机制等方面开展系统研究,以期进一步提高防治效率并拓展应用场景。
在水果保鲜技术领域,生物保鲜因其安全性高、实用性强且具有良好环境友好性,逐渐成为研究热点[36-37]。目前,在水果采后病害生物防治和保鲜方面应用最广的是芽孢杆菌属(Bacillus)细菌[38-39]。例如,周炜凯等[40]研究表明,巨大芽孢杆菌(Bacillus megaterium)及其次级代谢产物发酵液对芒果炭疽病菌和蒂腐病菌均具有显著抑制作用,同时能够明显抑制芒果果皮转黄,延缓果实衰老。植物内生菌作为与寄主长期共生的一类微生物群体,可通过养分竞争、空间位点竞争、产生拮抗物质、诱导植物系统抗性等多种机制增强植物抗逆与抗病能力。多项研究已证实内生菌在果蔬保鲜中的应用潜力,张敏等[41]从猕猴桃根、茎、叶和果实中分离到的内生菌MR-1,其发酵滤液处理的樱桃番茄(圣女果)保鲜效果最佳,其次为复合果蔬保鲜剂;储藏至第8天,与空白对照组相比,腐烂直径分别降低50.6%、50.6%和44.9%。周金伟等[42]的研究也表明,植物内生菌011的粗提液能够显著提高番茄的贮藏保鲜效果。
与上述报道类似,本研究选取健康茶树根系和枝条内生菌作为生防菌筛选来源,但来自茶树的菌株具有一定独特性。茶树在长期生长过程中会合成并释放茶多酚、咖啡碱、茶氨酸等多种生物活性物质,形成具有一定“化学压力”的微环境,我们推测长期适应这一特殊环境的菌株可能形成了与宿主互作的独特代谢策略:一方面,这些植物源化合物可能作为信号分子,调控菌株特定代谢通路(如与VOCs合成相关基因簇的表达);另一方面,它们也可能被菌株作为特殊前体物质加以利用,从而合成结构或功能上具有特异性的挥发性有机物(VOCs)。这种基于宿主特异性代谢物的互作关系,或许正是本研究菌株所产生VOCs在抑菌谱和保鲜效果上呈现出差异性与优势的重要原因。与来源于普通土壤或其他宿主植物的菌株相比,这一特性使其成为具有高度宿主适应性和较强靶向应用潜力的独特生物资源,为开发源自茶树微生态系统的专属生防制剂提供了理论支撑。因此,茶树来源菌株产生的VOCs在组分和功能上有望不同于土壤、海洋或其他植物来源菌株,这为发现新型抗菌VOCs提供了重要菌源。本研究从中筛选获得的解淀粉芽孢杆菌、苏云金芽孢杆菌、蜡样芽孢杆菌和枯草芽孢杆菌等典型生防类群,不仅在平板对峙中对多种病原真菌表现出高效拮抗能力,其产生的VOCs对芒果、芭蕉炭疽病原菌也表现出接近100.00%的强抑制作用,并在离体果实保鲜试验中展现出延缓腐烂、降低腐烂率的显著效果,进一步印证了茶树内生芽孢杆菌资源在水果保鲜领域的应用潜力。
目前,关于微生物VOCs在水果采后炭疽病和水果保鲜中应用的研究仍相对有限。本研究通过平板对扣法初步验证了拮抗菌株VOCs对果生炭疽菌和芭蕉炭疽菌的显著抑制作用,体现了其作为生物保鲜剂的潜在应用价值。本研究紧密围绕广西地区生产中高发的芒果炭疽病和芭蕉炭疽病开展试验,应用对象相对明确,所得数据对区域水果生产具有一定的实际指导意义。当然,本研究仍存在一些不足:(1) 试验中涉及的植物病原真菌种类有限,尚未系统评估拮抗菌株对更多水果病原真菌的抑制谱;(2) 仅以芒果和香蕉为试验对象,尚未验证其在广西其他特色水果(如荔枝霜疫霉病、砂糖橘青霉/绿霉病、火龙果病害等)上的保鲜效果。未来可在明确其对芒果、香蕉有效的基础上增加水果防病实验的样本量,将研究对象拓展至更多地方特色水果,评估其更广泛的适用性和应用潜力。此外,将菌株VOCs与低剂量咪鲜胺复配使用有望实现“增效减量”的协同效果,这也是后续值得重点探索的研究方向。
本研究尚未对拮抗菌株产生的VOCs成分进行解析,具体的活性物质基础和作用机制仍不清楚。后续研究的核心将是利用代谢组学等技术鉴定VOCs中的关键活性单体,并系统评估其单一及协同的抑菌效果及潜在分子靶点,可借助GC-MS或LC-MS等技术手段对菌株VOCs进行成分分离与鉴定,将鉴定出的主要挥发性单体分别开展抑菌试验,锁定关键活性组分,进一步阐明其抑菌及保鲜作用机制,为高效生物保鲜剂的开发提供理论依据。
总体来看,本研究筛选获得的拮抗菌株及其VOCs对多种植物病原真菌具有较强抑制活性,并在香蕉和芒果的采后保鲜中展现出良好应用前景,为开发多功能生物防治及保鲜制剂提供了新的菌种资源。然而,其活性物质组成、细胞靶点及寄主响应等方面仍有待深入研究。未来工作中一方面需进一步评价这些拮抗菌株在不同水果上的保鲜效果,拓展其在多种果品上的应用场景;另一方面可探索将生防菌或其VOCS与低剂量化学防治剂、物理保鲜技术等手段相结合,构建绿色、高效、可持续的综合防控策略,更好地服务于水果贮藏与保鲜生产实践。
引用本文: 吴姝静, 吕玟玟, 叶瑞萍, 罗玉珍, 孟宇翔, 谢晓莹, 姜明国, 周燕.茶树内生芽孢杆菌挥发性有机物对芒果和香蕉采后炭疽病的抑制与保鲜作用[J].微生物学报, 2026, 66(4): 1839-1857. (WU Shujing, LÜ Wenwen, YE Ruiping, LUO Yuzhen, MENG Yuxiang, XIE Xiaoying, JIANG Mingguo, ZHOU Yan. Postharvest anthracnose-inhibiting and preservative effects of volatile organic compounds produced by endophytic Bacillus from tea plants on mangoes and bananas[J]. Acta Microbiologica Sinica, 2026, 66(4): 1839-1857.)
基金信息: 广西民族大学国家级重点支持领域大学生创新创业训练计划(202410608035X)
「推荐阅读 」
漳江口红树林沉积物可培养细菌多样性、产酶潜力及优势芽孢杆菌特征
叶片喷施贝莱斯芽孢杆菌提升番薯产量的生理响应及内生菌-代谢通路调控机制
当年目次
欢迎加入科微学者交流群
扫描二维码
关于本刊
《微生物学报》(创刊于1953年,月刊)是由中国科学院微生物研究所和中国微生物学会共同主办,国内外公开发行的综合性学术期刊,是中文核心期刊、中国科技核心期刊、中国科学引文数据库核心期刊。本刊致力于报道国内外微生物学研究领域最新科研成果和研究进展,促进学术交流与合作。主要刊登普通微生物学、工业、农业、医学和兽医微生物学、免疫学以及与微生物学领域交叉的原创性研究论文和综述。已被Scopus、CA、BA、Cambridge Abstract、AJ、IC、JST、CNKI、WJCI等国内外多家数据库收录。荣获“中国精品科技期刊”“中国国际影响力优秀期刊”“中国科协精品科技期刊工程项目”“中国百篇最具影响国内学术论文”“中国科协优秀论文”“中国精品科技期刊顶尖学术论文(F5000)”等多项荣誉。我们期待与微生物领域的科研工作者携手,共同推动微生物学科的繁荣发展。欢迎投稿,分享您的科研成果和学术见解。
期刊订阅:
1. 直接联系联合编辑部订购,发行部 E-mail:bjb@im.ac.cn ;Tel:010-64807336;
2. 各地邮局订阅:邮发代号2-504;
3. 网上购买:搜淘宝店、微店店铺名称:中科期刊(订阅及销售过刊);科学出版社期刊发行部:联系电话010-64017032 64017539;
或扫描下方二维码:
更多内容详见原文
100 项与 上海麦克林生化科技股份有限公司 相关的药物交易
登录后查看更多信息
100 项与 上海麦克林生化科技股份有限公司 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2026年07月21日管线快照
管线布局中药物为当前组织机构及其子机构作为药物机构进行统计,早期临床1期并入临床1期,临床1/2期并入临床2期,临床2/3期并入临床3期
临床前
3
登录后查看更多信息
当前项目
| 药物(靶点) | 适应症 | 全球最高研发状态 |
|---|---|---|
Hesperetin ( XO ) | 高尿酸血症 更多 | 临床前 |
Stachydrine | 高尿酸血症 更多 | 临床前 |
Magnesium-L-threonate | 阿尔茨海默症 更多 | 临床前 |
登录后查看更多信息
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
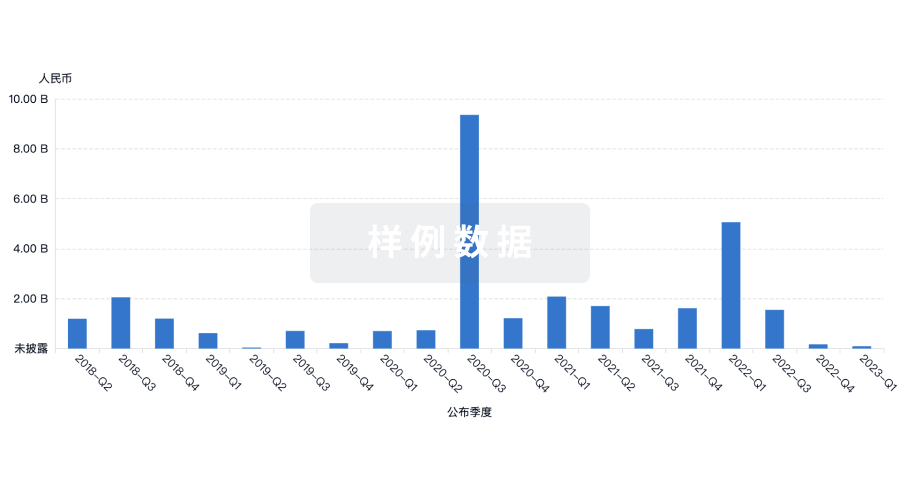
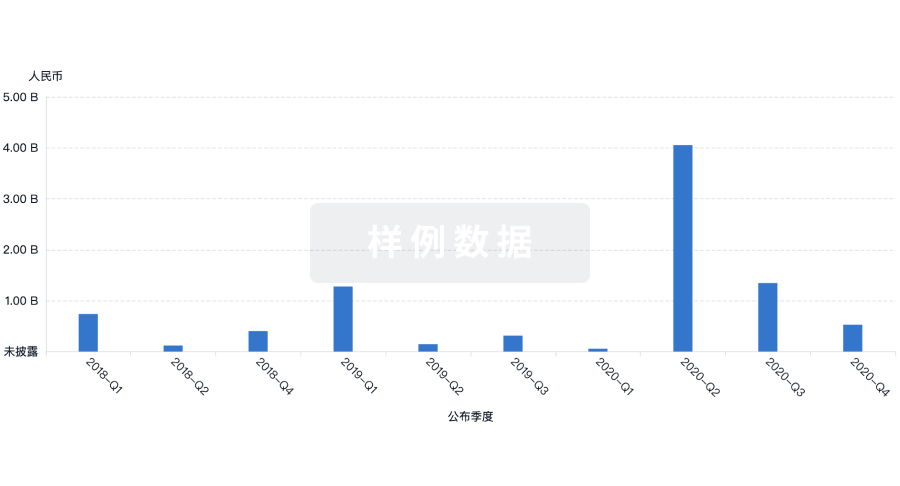
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
芽仔
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用