预约演示
更新于:2026-07-08
Sangon Biotech (Shanghai) Co., Ltd.
更新于:2026-07-08
概览
标签
心血管疾病
肿瘤
免疫系统疾病
单克隆抗体
疾病领域得分
一眼洞穿机构专注的疾病领域

技术平台
公司药物应用最多的技术
靶点
公司最常开发的靶点
关联
CN116813774
专利挖掘靶点 |
作用机制 |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症 |
最高研发阶段 |
首次获批国家/地区 |
首次获批日期 |
100 项与 生工生物工程(上海)股份有限公司 相关的临床结果
登录后查看更多信息
登录后查看更多信息
2025-07-01CANCER LETTERS
CD44 identified as a diagnostic biomarker for highly malignant CA19-9 negative pancreatic cancer
Article
作者: Yan, Chao ; Tang, Jiatong ; Dai, Zhan ; Wei, Wei ; Zhang, Shuo ; Qiu, Yudong ; Li, Qi ; Tang, Neng ; Du, Yixiang ; Qiu, Tongtong ; Zhang, Yixuan ; Zhang, Yifan ; Cheng, Hao ; Hong, Hao ; Hang, Hexing ; He, Jian ; Lin, Xiawen ; Li, Xiaoyang
Pancreatic ductal adenocarcinoma (PDAC) is a highly aggressive cancer with limited diagnostic biomarkers. Carbohydrate antigen 19-9 (CA19-9) is a widely used clinical biomarker and is generally considered to correlate with PDAC malignancy. However, the relationship between CA19-9 expression levels and tumor aggressiveness remains underexplored. In this study, we report a biphasic relationship between CA19-9 expression levels and PDAC malignancy, where both negative (<5 U/mL) and high (>37 U/mL) CA19-9 levels are associated with increased tumor aggressiveness. We defined CA19-9 negative PDAC as tumors that lack CA19-9 expression intracellulary, on the cell membrane, and in secreted form. In PDAC cell lines and patient-derived organoids, CA19-9 negativity, confirmed by immunofluorescence, flow cytometry and ELISA, correlated with more aggressive behaviors. In PDAC patients, tumors from those with serum CA19-9 levels below 5 U/mL exhibited stronger metabolically activity, more immunosuppressive tumor microenvironment, and worse survival than CA19-9 positive tumors, with over 90 % showing absent CA19-9 expression by immunohistochemistry (IHC). Glycoproteomics profiling identified CD44 as a highly expressed biomarker in CA19-9 negative PDAC. Elevated CD44 expression effectively distinguished CA19-9 negative PDAC from both CA19-9 positive PDAC and CA19-9 negative benign pancreatic diseases, suggesting its potential as a diagnostic tool. Furthermore, we developed a radionuclide-labeled CD44 antibody 89Zr-1M2E3, which specifically recognized CA19-9 negative PDAC tumors in preclinical models using PET-CT imaging. These findings highlight CD44 as a promising biomarker and therapeutic target for diagnosing and treating CA19-9 negative PDAC.
2023-05-01Journal of biotechnology
Deleting specific residues from the HNH linkers creates a CRISPR-SpCas9 variant with high fidelity and efficiency
Article
作者: Wang, Guohua ; Anderson, Christopher M ; Wang, Canmao ; Huang, Dongwei ; Li, Juan ; Chu, Teng ; Wu, Xinjun
Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) and CRISPR-associated (Cas) systems are immunological defenses used in archaea and bacteria to recognize and destroy DNA from external invaders. The CRISPR-SpCas9 system harnessed from Streptococcus pyogenes (SpCas9) has become the most widely utilized genome editing tool and shows promise for clinical application. However, the off-target effect is still the major challenge for the genome editing of CRISPR-SpCas9. Based on analysis of the structure and cleavage procedures, we proposed two strategies to modify the SpCas9 structure and reduce off-target effects. Shortening the HNH or REC3 linkers (Strategy #1) aimed to move the primary position of HNH or REC3 far away from the single-guide RNA (sgRNA)/DNA hybrid (hybrid), while elongating the helix around the sgRNA (Strategy #2) aimed to strengthen the contacts between SpCas9 and the sgRNA/DNA. We designed 11 SpCas9 variants (variant No.1- variant No.11) and verified their efficiencies on the classic genome site EMX1-1, EMX1-1-OT1, and EMX1-1-OT2. The top three effective SpCas9 variants, variant No.1, variant No.2, and variant No.5, were additionally validated on other genome sites. The further selected variant No.1 was compared with two previous SpCas9 variants, HypaCas9 (a hyper-accurate Cas9 variant released in 2017) and eSpCas9 (1.1) (an "enhanced specificity" SpCas9 variant released in 2016), on two genome sites, EMX1-1 and FANCF-1. The results revealed that the deletion of Thr769 and Gly906 could substantially decrease off-target effects, while maintaining robust on-target efficiency in most of the selected genome sites.
2016-06-01Methods (San Diego, Calif.)3区 · 生物学
Multi-omics data driven analysis establishes reference codon biases for synthetic gene design in microbial and mammalian cells
3区 · 生物学
Article
作者: Li, Wei ; Lee, Dong-Yup ; Ang, Kok Siong ; Kyriakopoulos, Sarantos
In this study, we analyzed multi-omics data and subsets thereof to establish reference codon usage biases for codon optimization in synthetic gene design. Specifically, publicly available genomic, transcriptomic, proteomic and translatomic data for microbial and mammalian expression hosts, Escherichia coli, Saccharomyces cerevisiae, Pichia pastoris and Chinese hamster ovary (CHO) cells, were compiled to derive their individual codon and codon pair frequencies. Then, host dependent and -omics specific codon biases were generated and compared by principal component analysis and hierarchical clustering. Interestingly, our results indicated the similar codon bias patterns of the highly expressed transcripts, highly abundant proteins, and efficiently translated mRNA in microbial cells, despite the general lack of correlation between mRNA and protein expression levels. However, for CHO cells, the codon bias patterns among various -omics subsets are not distinguishable, forming one cluster. Thus, we further investigated the effect of different input codon biases on codon optimized sequences using the codon context (CC) and individual codon usage (ICU) design parameters, via in silico case study on the expression of human IFNγ sequence in CHO cells. The results supported that CC is more robust design parameter than ICU for improved heterologous gene design.
100 项与 生工生物工程(上海)股份有限公司 相关的药物交易
登录后查看更多信息
100 项与 生工生物工程(上海)股份有限公司 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2026年07月26日管线快照
管线布局中药物为当前组织机构及其子机构作为药物机构进行统计,早期临床1期并入临床1期,临床1/2期并入临床2期,临床2/3期并入临床3期
药物发现
1
登录后查看更多信息
当前项目
登录后查看更多信息
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
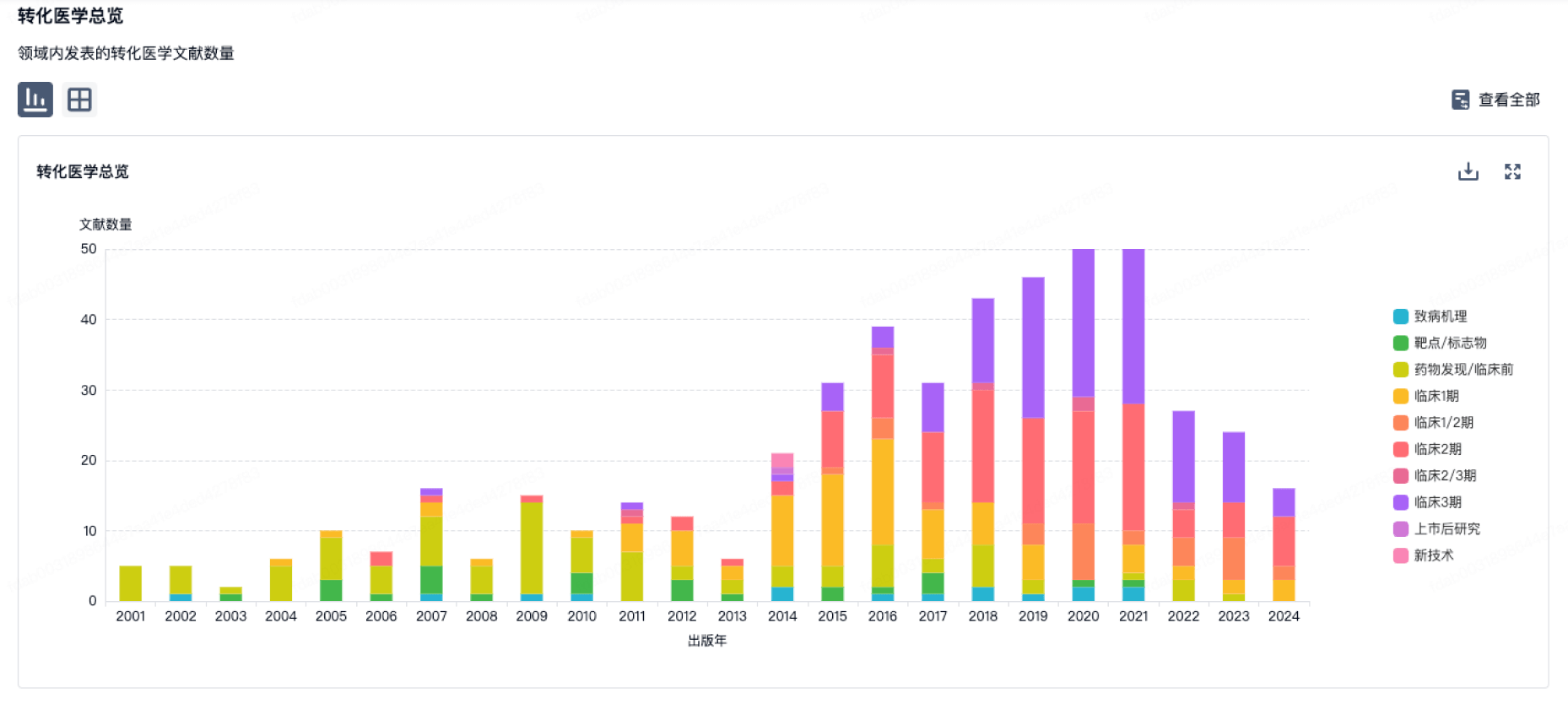
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
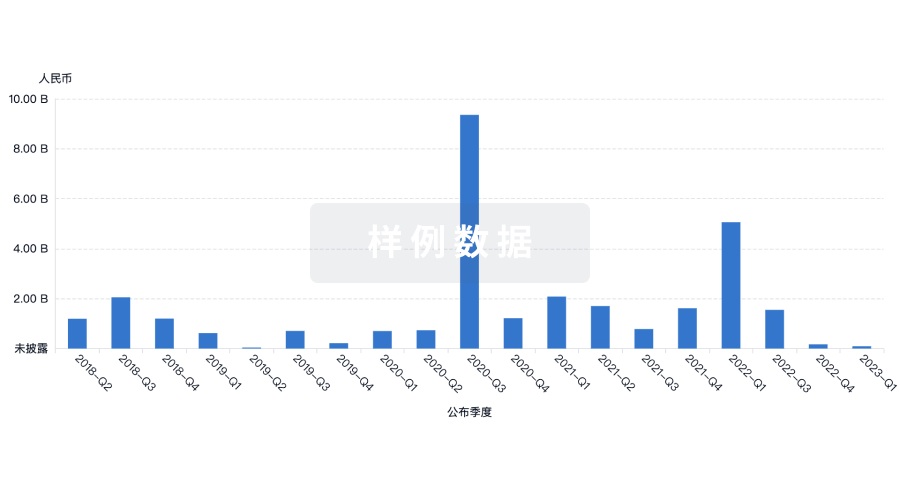
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
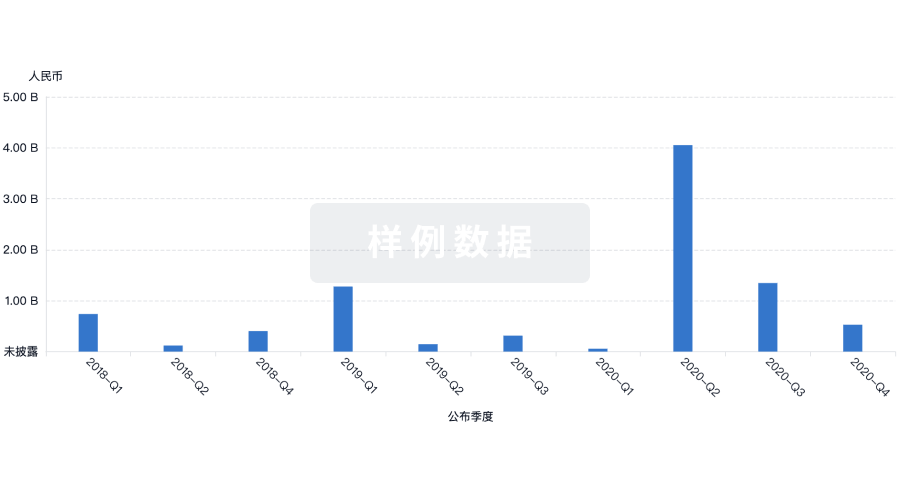
芽仔
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用