预约演示
更新于:2026-06-04
上海赛徕克生物科技有限公司
更新于:2026-06-04
概览
标签
其他疾病
呼吸系统疾病
小分子化药
疾病领域得分
一眼洞穿机构专注的疾病领域

暂无数据
技术平台
公司药物应用最多的技术
暂无数据
靶点
公司最常开发的靶点
暂无数据
| 排名前五的药物类型 | 数量 |
|---|---|
| 小分子化药 | 1 |
| 排名前五的靶点 | 数量 |
|---|---|
| Smad proteins x TGF-β | 1 |
关联
1
项与 上海赛徕克生物科技有限公司 相关的药物100 项与 上海赛徕克生物科技有限公司 相关的临床结果
登录后查看更多信息
0 项与 上海赛徕克生物科技有限公司 相关的专利(医药)
登录后查看更多信息
4
项与 上海赛徕克生物科技有限公司 相关的新闻(医药)2026-05-29
引用本文
费庆林,叶龙云,吴伟顶,等. SUMO2介导的RRM1类泛素化修饰调控胰腺导管腺癌吉西他滨耐药[J]. 中国癌症杂志, 2026, 36 (4): 323-332.
基金项目:国家自然科学基金(U21A20374,82173091)。上海市抗癌协会“雏鹰”计划(SACA-CY23B02)。
利益冲突:所有作者均声明无利益冲突。
伦理批件:FUSCC-IACUC-2023280。
知情同意:不需要。
通信作者简介
虞先濬,主任医师,二级教授,博士研究生导师,复旦大学附属肿瘤医院院长,上海市质子重离子医院院长,上海市质子重离子临床技术研发中心主任,复旦大学附属肿瘤医院临床研究中心、 Ⅰ期临床中心主任,上海市胰腺肿瘤研究所所长,复旦大学胰腺肿瘤研究所所长,上海市胰腺肿瘤精准治疗重点实验室主任。国家杰出青年科学基金获得者,科技部“中青年科技创新领军人才”,国务院政府特殊津贴、全国五一劳动奖章获得者,上海市科技精英,上海工匠,上海市领军人才。中国抗癌协会胰腺癌专业委员会主任委员,中华医学会外科学分会胰腺外科学组委员,中国胰腺病学会常委,上海市医学会普外科专科分会副主任委员,美国外科医师学院 Fellow(FACS)。获上海市科技进步奖一等奖、中国抗癌协会科技奖一等奖、上海医学科技奖一等奖、上海市级医院临床创新奖、上海市职工优秀创新成果奖一等奖。主持四大慢病国家科技重大专项1项,国家杰出青年科学基金1项,国家自然科学基金区域合作重点项目1项,国家自然科学基金中德合作项目 1 项,国家自然科学基金面上项目 3 项,省部级项目 15 项。以通信作者在Cancer Cell、Journal of Clinical Oncology、Cell Research、Gastroenterology、Gut、Annals of Surgery国际权威SCI收录期刊上发表论文260余篇。
文章视频解读
SUMO2介导的RRM1类泛素化修饰调控胰腺导管腺癌吉西他滨耐药
费庆林,叶龙云,吴伟顶,金凯舟,虞先濬
复旦大学附属肿瘤医院胰腺外科,复旦大学上海医学院肿瘤学系,上海市胰腺肿瘤研究所,上海市胰腺肿瘤精准诊疗重点实验室,复旦大学胰腺肿瘤研究所,上海 200032
[摘要] 背景与目的:胰腺导管腺癌(pancreatic ductal adenocarcinoma,PDAC)是高度致命的恶性肿瘤,患者5年生存率不足10%。吉西他滨作为PDAC一线化疗药物,常因肿瘤细胞获得性耐药而失效。类泛素化修饰在癌症耐药中发挥关键作用,但其在PDAC吉西他滨耐药中的机制尚未完全阐明。本研究旨在探讨类泛素化修饰在PDAC耐药中的作用,识别关键SUMO亚型及其靶蛋白。方法:建立吉西他滨耐药的PANC1细胞系(PANC1_GR),通过蛋白质印迹法(Western blot)检测SUMO1、SUMO2和SUMO3的表达水平。采用SUMO激活酶抑制剂ML-792进行药物干预,用细胞计数试剂盒(cell counting kit-8,CCK-8)和流式细胞术分别评估细胞增殖与凋亡情况。通过慢病毒介导敲低SUMO1、SUMO2、SUMO3及RRM1,检测敲低后细胞对吉西他滨的敏感性(IC50)及凋亡变化。利用SUMO2免疫共沉淀结合蛋白质组质谱分析鉴定SUMO2的关键修饰靶蛋白。通过转录组测序分析吉西他滨联合ML-792处理后的差异表达基因,并进行通路富集分析。统计分析采用t检验或单因素方差分析(ANOVA)。本研究获复旦大学附属肿瘤医院实验中心动物伦理委员会批准(伦理批准号:FUSCC-IACUC-2023280)。结果:吉西他滨耐药细胞中SUMO1、SUMO2、SUMO3修饰水平均显著升高,类泛素化修饰抑制剂ML-792可降低整体SUMO2、SUMO3修饰并增强吉西他滨诱导的凋亡。对比不同类泛素化修饰类型后发现,仅SUMO2敲低可显著提高吉西他滨敏感性。SUMO2、SUMO3免疫共沉淀结合质谱分析鉴定RRM1为SUMO2的关键修饰靶蛋白,其SUMO2修饰水平在耐药细胞中明显提升,而ML-792可显著抑制RRM1的SUMO2修饰。RRM1敲低可模拟SUMO2抑制效应,加强吉西他滨诱导的凋亡并降低IC50。转录组分析显示,阻断SUMO2修饰影响DNA复制、错配修复等耐药相关通路,从而削弱细胞的化疗抵抗。综上,SUMO2介导的RRM1类泛素化修饰在PDAC吉西他滨耐药中发挥调控作用。结论:SUMO2-RRM1轴可能参与PDAC对吉西他滨耐药的形成过程,靶向干预RRM1的SUMO2修饰有望为改善PDAC化疗耐药提供新的研究思路。
[关键词] 类泛素化修饰;SUMO2;RRM1;吉西他滨耐药;胰腺导管腺癌
[Abstract] Background and purpose: Pancreatic ductal adenocarcinoma (PDAC) is one of the most lethal malignancies, with a 5-year survival rate below 10%. Gemcitabine remains a first-line chemotherapeutic agent for PDAC; however, its efficacy is frequently compromised by the development of acquired drug resistance. SUMOylation, a key post-translational modification, has been implicated in cancer progression and chemoresistance, yet its role in gemcitabine resistance in PDAC remains insufficiently defined. This study aimed to investigate the contribution of SUMOylation to PDAC chemoresistance and to identify the critical SUMO isoform and its downstream substrates. Methods: A gemcitabine-resistant PANC1 cell line (PANC1_GR) was established, and the expression of SUMO1, SUMO2, and SUMO3 was assessed by Western blot. The SUMO-activating enzyme inhibitor ML-792 was used to suppress global SUMOylation, and cell proliferation and apoptosis were evaluated using the cell counting kit-8 (CCK-8) assay and flow cytometry, respectively. Lentiviral-mediated knockdown of SUMO1, SUMO2, SUMO3, and RRM1 was performed to assess changes in gemcitabine sensitivity (IC50) and apoptosis. SUMO2 immunoprecipitation combined with mass spectrometry was employed to identify SUMO2-modified target proteins. Transcriptomic sequencing following gemcitabine plus ML-792 treatment was used to analyze differentially expressed genes and pathway enrichment. Statistical analyses were conducted using Student's t-tests or one-way ANOVA. This study was approved by the Animal Ethics Committee of the Experimental Center of Fudan University Shanghai Cancer Center (ethics approval No. FUSCC-IACUC-2023280). Results: SUMO1, SUMO2, SUMO3 conjugation levels were markedly elevated in gemcitabine-resistant cells, and ML-792 effectively reduced global SUMO2, SUMO3 modification while enhancing gemcitabine-induced apoptosis. Among the SUMO isoforms, only SUMO2 knockdown significantly increased gemcitabine sensitivity. SUMO2, SUMO3 Co-IP combined with mass spectrometry identified RRM1 as a major SUMO2-modified substrate, with its SUMOylation markedly enhanced in resistant cells and significantly suppressed by ML-792. RRM1 knockdown phenocopied the effects of SUMO2 inhibition, promoting apoptosis and reducing the IC50 of gemcitabine. Transcriptomic analysis further revealed that inhibition of SUMO2 modification disrupted key pathways associated with drug resistance, including DNA replication and mismatch repair, thereby diminishing cellular chemoresistance. Collectively, these findings indicate that SUMO2-mediated SUMOylation of RRM1 may play a regulatory role in gemcitabine resistance in PDAC. Conclusion: The SUMO2–RRM1 axis may be associated with the development of gemcitabine resistance in PDAC, and modulation of SUMO2-mediated RRM1 modification could offer a potential direction for further investigation.
[Key words] SUMOylation; SUMO2; RRM1; Gemcitabine resistance; Pancreatic ductal adenocarcinoma
胰腺导管腺癌(pancreatic ductal adeno-carcinoma,PDAC)是全球极具致死性的恶性肿瘤之一,患者的5年生存率不足10%[1]。这主要归咎于疾病的隐匿起病、侵袭性强,也缺乏有效的治疗手段。吉西他滨作为一种通过抑制DNA合成发挥作用的核苷类似物,自1997年获批以来一直是PDAC化疗的一线药物[2-3]。然而,肿瘤细胞的固有或获得性耐药可显著削弱吉西他滨的治疗效果,导致多数患者最终治疗失败[4]。因此,深入阐明吉西他滨耐药的分子机制,对于探索克服耐药的新策略并改善PDAC患者的预后具有重要意义。
蛋白质翻译后修饰在调控细胞生物学过程(包括药物耐受性)中发挥关键作用。其中,类泛素化修饰,即类泛素样修饰蛋白(SUMO)可逆地与底物蛋白共价结合的过程,已被证实是肿瘤发生、转移及多种癌症(包括PDAC)治疗耐药的重要调控机制[5-6]。SUMO家族包括SUMO1、SUMO2和SUMO3,其中SUMO2和SUMO3具有高度的序列同源性,常具有功能冗余性,而SUMO1则通常修饰不同的靶蛋白[7-9]。在肿瘤中,SUMO化水平的异常调控可导致蛋白稳定性、定位及活性的改变,从而促进肿瘤对化疗耐药[10-11]。例如,早幼粒细胞白血病蛋白的低类泛素化修饰状态与吉西他滨和奥沙利铂耐药相关,这一过程通过激活NF-κB和CREB信号转导通路实现[12]。此外,抑制类泛素化修饰通路已被证实能够通过调节肿瘤微环境并增强抗肿瘤免疫反应,从而提高治疗的敏感性[13]。
核苷酸还原酶大亚基(ribonucleotide reductase M1,RRM1)是核苷酸还原酶的重要组成部分,在从头合成脱氧核苷酸和DNA修复过程中发挥关键作用[14-15]。其高表达是PDAC吉西他滨耐药的重要生物标志。然而,调控RRM1活性的分子机制,尤其是通过类泛素化修饰等蛋白质翻译后修饰途径的机制,尚未得到充分研究[16]。本研究旨在探讨类泛素化修饰在促进PDAC细胞吉西他滨耐药中的作用,重点识别特定的SUMO亚型及其靶蛋白,以期揭示逆转耐药的新型治疗靶点。
1
材料和方法
1.1 主要试剂
吉西他滨(HY-17026)、ML-792(HY-108702)购自美国MedChemExpress公司,胎牛血清(fetal bovine serum,FBS;FSD500)购自苏州依科赛生物科技股份有限公司,青-链霉素(双抗)购自美国赛默飞公司,DMEM培养基、磷酸缓冲盐溶液(phosphate-buffered saline,PBS)购自上海源培生物科技股份有限公司,细胞计数试剂盒(cell counting kit-8,CCK-8)细胞活性检测试剂(B34304)购自美国Selleck公司,细胞凋亡检测试剂盒(AD11)购自上海东仁化学科技有限公司,SUMO1抗体(4940S)、SUMO2抗体(4971T)购自美国CST公司,RRM1抗体(R25625)购自成都正能生物技术有限公司,ACTB抗体(AC026)购自武汉爱博泰克生物科技有限公司,SUMO2/3 IP试剂盒(BK162-S)购自美国Cytoskeleton公司,十二烷基硫酸钠聚丙烯酰胺凝胶电泳(sodium dodecylsulphate polyacrylamide gel electrophoresis,SDS-PAGE)凝胶快速制备试剂盒(PG112)、电化学发光(electrochemical luminescence,ECL)试剂盒(SQ202)购自上海雅酶生物医药科技有限公司。
1.2 细胞培养
人类胰腺癌细胞系PANC1、MIA PaCa-2及HEK-293T均购自中国科学院典型培养物保藏委员会细胞库/中国科学院上海生命科学研究院细胞资源中心,所有细胞系均已通过中国科学院典型培养物保藏委员会细胞库进行基因型鉴定,并排除了细菌污染。所有细胞均置于含有10%的FBS、100 U/mL青霉素和100 μg/mL链霉素的完全培养基中,每隔2~3 d换液,上述细胞均在加湿培养箱(37 ℃、CO2体积分数为5%、95%湿度)中培养,保证细胞维持在最佳生长密度。
1.3 病毒感染构建SUMO1、SUMO2、SUMO3、RRM1敲除细胞株
根据张锋实验室提供的sgRNA文库(https://www.zlab.bio/resources)中的信息,设计SUMO1、SUMO2、SUMO3、RRM1 sgRNA序列,运用基因重组技术将其克隆至lentiCRISPR v2(Addgene,52961)载体中,构建敲除质粒,并使用Sanger测序进行验证。使用重组质粒和慢病毒包装质粒共转染HEK-293T细胞,收集包含慢病毒的细胞上清液,过滤离心后感染PANC1、MIA PaCa-2细胞,构建SUMO1、SUMO2、SUMO3、RRM1敲除的细胞。使用蛋白质印迹法(Western blot)验证SUMO1、SUMO2、SUMO3、RRM1表达情况,明确敲除效率,选择效果最好的稳转株进行功能实验。
1.4 CCK-8法检测细胞增殖活力及药物IC50
取对数生长期细胞,以1×104个/孔(100 μL)接种于96孔板,每组设5个复孔。贴壁过夜后加入含不同浓度吉西他滨(0、0.01、0.05、0.1、0.5、1、5、10、50、100 μmol/L)的完全培养基,继续培养48 h。每孔加入10 μL CCK-8试剂,37 ℃温育2 h后,使用多功能酶标仪测定450 nm波长处吸光度(D)值。以未加药组存活率为100%,计算各浓度组细胞存活率,使用GraphPad Prism 9.5.1线性回归拟合剂量-效应曲线并计算IC50值。
1.5 凋亡检测
根据AnnexinⅤ633细胞凋亡检测试剂盒说明,消化收集不同处理组的细胞,离心后弃去上清液,使用1 mL PBS洗涤后,加入500 μL缓冲液,吹打重悬细胞。在样品中加入5 μL AnnexinⅤ633和5 μL PI染料,避光室温温育15 min,1 h内进行流式细胞术检测。
1. 6 Western blot检测
使用细胞裂解液提取细胞总蛋白,蛋白定量后取等量总蛋白进行SDS-PAGE蛋白电泳,350 mA恒流转膜;用5%脱脂牛奶封闭后,按照说明书加入配制的一抗,4 ℃摇床过夜温育,洗涤后加入对应二抗温育;使用ECL试剂显影后拍照。
1.7 免疫共沉淀实验
收集细胞,使用裂解液提取细胞总蛋白,离心后留取裂解上清液。取部分裂解上清液与上样缓冲液混匀,98 ℃加热10 min,作为Input样本。在细胞裂解液中加入40 μL抗SUMO2/3的琼脂糖珠,4 ℃温育过夜;离心后弃上清液,洗涤琼脂糖珠后,加入上样缓冲液,98 ℃加热10 min作为IP样本;使用Western blot检测IP效率。进一步使用蛋白质谱检测各组IP样本中蛋白表达的差异。
1.8 构建裸鼠皮下移植瘤模型
6~8周龄BALB/c裸鼠(雄性)购自南京集萃药康生物科技股份有限公司。肿瘤细胞以200 μL PBS重悬,接种于小鼠背部皮下以构建皮下移植瘤模型。待肿瘤体积达到500 mm3时,腹腔注射吉西他滨(400 ng/只)治疗,连续给药2周。治疗结束后处死小鼠,完整剥离肿瘤组织并测量肿瘤体积。肿瘤组织进行免疫组织化学染色分析。所有动物实验均严格遵循复旦大学附属肿瘤医院实验中心动物伦理委员会制定的《实验动物饲养和使用指南》,并获得该伦理委员会批准(伦理批准号:FUSCC-IACUC-2023280)。
1.9 转录组测序及数据分析
PANC1细胞经吉西他滨单药或联合ML-792处理后,使用裂解液提取总RNA并送北京诺禾致源科技股份有限公司进行文库构建与高通量测序。获得原始表达矩阵后,利用R语言DESeq2 1.46.0包筛选组间差异表达基因,并通过clusterProfiler 4.14.6包进行京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)通路富集分析及火山图绘制;此外,从癌症基因组图谱(The Cancer Genome Atlas,TCGA)数据库(https://www.cancer.gov/ccg/research/genome-sequencing/tcga)下载PDAC队列的转录组及临床数据,利用survival 3.7-0和survminer 0.5.0 R包进行Kaplan-Meier生存分析与log-rank检验,以评估关键基因对患者预后的影响。
1.10 统计学处理
使用GraphPad Prism 9.5.1软件对数据进行统计学分析,计量数据以x̄±s表示。在进行参数检验前,首先采用Shapiro-Wilk检验对数据的正态性分布进行评估。符合正态分布且方差齐性的数据,两组间比较采用Student t检验(双侧),多组间比较采用单因素方差分析(One-way ANOVA)辅以LSD事后检验。所有统计学分析均采用双侧检验。P<0.05为差异有统计学意义。
2
结 果
2.1 类泛素化修饰促进胰腺癌细胞吉西他滨耐药
通过逐步增加PANC1细胞培养基中吉西他滨的浓度,成功建立了吉西他滨耐药株PANC1_GR。IC50检测结果显示,PANC1_GR细胞对吉西他滨的耐受性显著增强,表明耐药模型构建成功(图1A)。进一步检测类泛素化修饰水平发现,耐药细胞中SUMO1、SUMO2及SUMO3修饰水平均显著升高(图1B)。为探讨类泛素化修饰在吉西他滨耐药中的作用,本研究采用类泛素化修饰抑制剂ML-792进行干预。本研究使用0.001~10 μmol/L不同浓度的ML-792处理细胞,以评估其对细胞增殖及类泛素化修饰水平的影响。CCK-8结果显示,当ML-792浓度高于1 μmol/L后,会对细胞增殖产生明显抑制作用;而Western blot检测结果表明,ML-792在浓度≥0.1 μmol/L时即可显著降低细胞内整体类泛素化修饰水平。综合考虑在不显著影响细胞活力的前提下实现对SUMO修饰的有效抑制,最终选取1 μmol/L ML-792作为后续实验的工作浓度(图1C、D)。体外凋亡分析结果显示,ML-792与吉西他滨联合处理可显著增强PANC1和MIA PaCa-2细胞的凋亡效应(图1E、F)。上述结果表明,类泛素化修饰的增强可能促进胰腺癌细胞对吉西他滨治疗的耐药性。
图1 类泛素化修饰对胰腺癌细胞吉西他滨耐药的影响
Fig. 1 SUMOylation effects gemcitabine resistance in pancreatic cancer cells
A: IC50 values of PANC1 gemcitabine-resistant cells. B: Protein expression levels of SUMO1 and SUMO2/3 in resistant versus control cells. C: CCK-8 assay results showing the inhibitory effect of the SUMO-activating enzyme inhibitor ML-792. D: Western blot analysis of SUMO1 and SUMO2/3 expression following treatment with increasing concentrations of ML-792. E-F: Flow cytometry analysis of apoptosis in PANC1 and MIA PaCa-2 cells after combined treatment with ML-792 and gemcitabine. Data were presented as the x̄±s. *: P<0.05; **: P<0.01; ***: P<0.001; ****: P<0.000 1; ns: Not significant. GR: Gemcitabine-resistant. GEM: Gemcitabine. MG: Combined treatment with ML-792 and gemcitabine.
2.2 敲低SUMO2增强胰腺癌细胞吉西他滨敏感性
为明确具体哪种SUMO修饰在吉西他滨耐药中发挥关键作用,本研究分别在PANC1和MIA PaCa-2细胞中敲低SUMO1、SUMO2和SUMO3的表达。Western blot检测结果显示,各目标基因表达水平均显著降低(图2A、B)。功能实验发现,敲低SUMO1或SUMO3并未显著影响吉西他滨诱导的细胞凋亡(P>0.05),而敲低SUMO2可显著提高吉西他滨诱导的细胞凋亡比例(P<0.01,图2C~H)。IC50检测进一步证实,SUMO2敲低后PANC1及MIA PaCa-2细胞对吉西他滨的敏感性显著提高,而SUMO1或SUMO3敲低对药物敏感性无显著影响(表1)。在体内实验中,将SUMO2敲低的PANC1和MIA PaCa-2细胞接种于裸鼠皮下后,相较于对照组,吉西他滨对肿瘤生长的抑制作用更加显著(图3A);免疫组织化学检测结果进一步显示,SUMO2敲低组肿瘤组织中细胞增殖水平明显降低(图3B)。上述结果提示,SUMO2介导的类泛素化修饰在胰腺癌对吉西他滨耐药的形成中可能具有重要的调控作用。
图2 敲低SUMO2对胰腺癌细胞吉西他滨敏感性的影响
Fig. 2 Knockdown of SUMO2 modulated gemcitabine sensitivity in pancreatic cancer cells
A-B: Western blot validation of SUMO1, SUMO2, and SUMO3 knockdown in pancreatic cancer cell lines. C-D: Apoptosis analysis of PANC1 and MIA PaCa-2 cells following SUMO1 knockdown. E-F: Apoptosis analysis of PANC1 and MIA PaCa-2 cells following SUMO3 knockdown. G-H: Apoptosis analysis of PANC1 and MIA PaCa-2 cells following SUMO2 knockdown, demonstrating the strongest enhancement of gemcitabine-induced apoptosis. Data were presented as the x±s. *: P<0.05, **: P<0.01; ***: P<0.001; ****: P<0.000 1; ns: Not significant. GEM: Gemcitabine.
图3 SUMO2敲低增强胰腺癌细胞对吉西他滨敏感性的体内实验研究
Fig. 3 In vivo study showed that SUMO2 knockdown enhances the sensitivity of pancreatic cancer cells to gemcitabine
A: Representative images of nude mouse xenografts derived from PANC1 and MIA PaCa-2 cells. Each group contained 5 animals. Tumor volume was measured and compared using Student’s t test. B: Xenograft tumor tissues were stained for Ki-67 proliferation (×20). Ki-67 proliferation-high cells were quantified and statistically analyzed by Student’s t test. Data were presented as the x̄±s. *: P<0.05; **: P<0.01; ***: P<0.001, ****: P<0.000 1; ns: Not significant.
表1 胰腺癌细胞IC50
Tab. 1 IC50 values of pancreatic cancer cells
2.3 RRM1是SUMO2修饰的关键蛋白
为鉴定SUMO2修饰的潜在靶蛋白,本研究对PANC1和PANC1_GR细胞进行SUMO2/3免疫共沉淀实验。结果显示,耐药细胞株中SUMO2/3修饰蛋白显著富集(图4A)。质谱分析提示,RRM1在PANC1_GR细胞中显著上调(表2),Western blot验证结果与之吻合(图4B)。同时,在PANC1耐药细胞株中亦观察到RRM1表达水平升高(图4C)。进一步使用ML-792处理后,PANC1和PANC1_GR细胞中整体SUMO2/3修饰水平、RRM1表达及RRM1的SUMO2/3修饰均明显下降(图4D、E)。反向免疫共沉淀实验同样显示,ML-792可降低RRM1与SUMO2/3的结合水平(图4F)。以上结果表明,RRM1是SUMO2修饰的关键靶蛋白,其SUMO2/3修饰在吉西他滨耐药细胞中显著增强,通过该修饰可提高RRM1蛋白稳定性、减少其降解,从而促进RRM1的累积并参与耐药表型的维持。
图4 Western blot 检测SUMO2、RRM1 的蛋白表达结果
Fig. 4 Western blot analysis of SUMO2 and RRM1 protein expression
A: Western blot analysis of SUMO2/3 following immunoprecipitation. B: Western blot analysis of RRM1 following immunoprecipitation. C: Western blot analysis of RRM1 expression in gemcitabine-resistant PANC-1 cells. D-E: Western blot detection of SUMO2/3 and RRM1 after ML-792 treatment. F: Reverse immunoprecipitation showing Western blot results for SUMO2/3 and RRM1. GR: Gemcitabine-resistant. IP: Immunoprecipitation.
2.4 抑制RRM1的SUMO2修饰可影响RRM1功能并增强吉西他滨敏感性
TCGA数据库分析显示,RRM1高表达的胰腺癌患者预后差(图5B、C)。功能实验中,在PANC1及MIA PaCa-2细胞中敲低RRM1(图5A)可显著增强吉西他滨诱导的凋亡(P<0.01,图5D、E),并降低其IC50值(表1),与抑制SUMO2修饰的结果一致。为进一步探讨类泛素化修饰对吉西他滨疗效的分子机制,我们进行了转录组测序。结果显示,吉西他滨联合ML-792处理后,共有1 028个基因上调,415个基因下调(图5F)。KEGG富集分析提示,差异基因主要涉及MAPK信号转导通路、DNA复制及错配修复等关键通路(图5G)。联合处理后,与DNA复制相关的基因(RFC2、MCM2、POLA1、MCM4、RPA2、RPA1、PCNA、MCM7、RPA4)及错配修复相关基因(RFC2、MSH2、RPA2、RPA1、PCNA、RFC3、RPA4)表达均显著上调(图5H、I)。鉴于RRM1在DNA合成与修复中的重要作用,这些结果提示抑制RRM1的SUMO2修饰可能削弱其功能,从而增强胰腺癌细胞对吉西他滨的敏感性。
图5 抑制SUMO2修饰对RRM1功能的影响
Fig. 5 Effect of inhibiting SUMO2 modification on RRM1 function
A: Western blot validation of RRM1 knockdown in pancreatic cancer cell lines. B: Determination of the optimal cutoff value for RRM1 expression using the Cancer Genome Atlas datasets. C: Kaplan-Meier survival analysis comparing overall survival between RRM1-high (n=96) and RRM1-low (n=75) patient cohorts. D-E: Apoptosis analysis following RRM1 knockdown. F: Volcano plot of differentially expressed genes after combined treatment with gemcitabine and ML-792. G: Kyoto Encyclopedia of Genes and Genomes pathway enrichment Dot plot of differentially expressed genes. H-I: Heatmaps showing different gene expression in the mismatch repair and DNA replication pathways. Data were presented as the x̄±s. *: P<0.05; **: P<0.01; ***: P<0.001; ****: P<0.000 1; ns: Not significant. GEM: Gemcitabine. MG: Combined treatment with ML-792 and gemcitabine.
表2 免疫沉淀蛋白质谱结果
Tab. 2 Immunoprecipitation-based proteomic analysis results
3
讨 论
本研究揭示了SUMO2介导的SUMO化修饰在PDAC吉西他滨耐药中的核心作用,这一结论与既往关于PDAC耐药中类泛素化修饰整体失衡的研究结果一致[6,8],但进一步明确了SUMO2亚型的重要地位。既往研究多集中于SUMO化总体水平升高与化疗耐受性的相关性,而本研究通过特异性敲低实验证实,仅抑制SUMO2即可显著逆转吉西他滨耐药表型,而SUMO1或SUMO3的敲低几乎不影响该效应。尽管SUMO2与SUMO3在序列上高度同源、理论上具备一定的功能性补偿,但已有研究表明,在丁酸诱导的耐药细胞模型中,SUMO3修饰发挥主要调控作用[11],提示二者在功能上并非完全冗余。在吉西他滨诱导下的应激状态,本研究观察到SUMO2修饰发挥主要作用,这可能与其在应激状态下参与蛋白聚集抑制、RNA代谢调控及蛋白稳定性维持等方面的独特机制有关[13]。近期研究亦从不同角度支持SUMO亚型功能的异质性,有研究[17]表明SUMO化抑制可通过多重肿瘤微环境效应诱导PDAC的适应性抗肿瘤免疫,而2025年的研究[18]显示,联合抑制磷脂酰肌醇3-激酶(phosphoinositide 3-kinase,PI3K)α/δ与SUMO信号转导通路能够选择性杀伤PDAC细胞并激活免疫应答。上述证据与本研究结果相互补充,提示不同化疗药物类型(如吉西他滨与铂类药物)及细胞或微环境上下游环节可能驱动特定SUMO亚型的选择性激活。总体而言,尽管SUMO化作为广谱肿瘤调控机制已得到广泛认可,其亚型特异性仍有待系统阐明,这在一定程度上限制了精准靶向干预策略的临床转化。
选择性SUMO激活酶抑制剂ML-792在低浓度条件下即可显著降低细胞整体SUMO化水平,并与吉西他滨产生协同效应以增强肿瘤细胞凋亡。这一特性与临床前研究中报道的同类型抑制剂TAK-981高度一致,后者可通过激活Ⅰ型干扰素信号、阻滞细胞周期进程并重塑肿瘤微环境发挥多层面的抗肿瘤作用[12,18]。既往研究多聚焦于TAK-981的免疫调控功能,而本研究则凸显了SUMO通路抑制在直接逆转化疗耐药方面的潜在价值,尤其是在体外PDAC模型中表现出的与吉西他滨协同杀伤效应。近期研究进展亦在不断拓展该通路的治疗前景,多项研究证实TAK-981在PDAC模型中可增强T淋巴细胞介导的抗肿瘤免疫、减少调节性T淋巴细胞比例并激活先天免疫反应[17,19],同时与PI3K抑制剂联用可产生选择性的肿瘤杀伤效应[18]。由此可见,SUMO抑制剂已进入临床研究阶段并展现出多机制抗肿瘤潜力,但其在吉西他滨耐药背景下的直接体内证据仍相对有限,主要挑战在于开发亚型特异性药物及优化联合治疗方案,只有这样才能使治疗效果最大化。
免疫共沉淀联合质谱分析鉴定RRM1为耐药细胞中SUMO2的关键修饰底物,其SUMO化水平的升高通过增强蛋白稳定性,促进脱氧核苷酸合成及DNA损伤修复,从而直接对抗吉西他滨介导的链终止效应。这一发现在既往RRM1与PDAC化疗耐药及不良预后密切相关的研究基础上[4,19],进一步拓展了其分子调控层面的认识。功能实验显示,敲低RRM1可完全模拟SUMO2抑制所产生的表型,进一步揭示了SUMO2-RRM1轴作为调控吉西他滨耐药的重要通路。转录组测序结果表明,联合治疗后MAPK信号转导通路、DNA复制及错配修复相关基因显著上调,提示RRM1功能受抑后细胞启动代偿性应激与修复反应,这与既往关于SUMO化在DNA损伤应答及核苷酸代谢稳态调控中发挥广泛作用的报道[13]一致。相较于现有研究主要聚焦于RRM1的转录水平或表观遗传调控,本研究将RRM1与翻译后SUMO化修饰相联系,揭示了其更为上游且精细的调控机制。近期基于TCGA数据库的分析亦提示SUMO底物相关基因可作为PDAC预后标志物[20],但尚未特异指向RRM1,进一步凸显了本研究的创新性;同时也暴露出当前研究领域的不足,即SUMO底物谱的系统鉴定、SUMO聚合的动态形式及位点特异性功能仍存在较大研究空白,有待进一步深入探索。
尽管本研究为SUMO2-RRM1轴作为PDAC潜在治疗靶点提供了有力证据,但仍存在一定局限性。首先,研究主要基于体外模型,难以充分反映体内肿瘤异质性、基质-上皮互作及复杂免疫微环境,可能高估协同效应[20]。其次,所用细胞系局限于经典PDAC模型,未覆盖不同分子亚型,影响结论的泛化性。此外,Western blot定量偏差、质谱对低丰度/瞬时SUMO化底物的检出能力限制、转录组分析中多重检验控制不足,以及耐药细胞系构建过程中的克隆选择偏倚,均可能影响结果稳健性。未来需在原位移植瘤、PDX及基因工程小鼠模型中进一步验证,并结合临床队列评估该轴的表达异质性。同时,探索SUMO抑制剂(如ML-792/TAK-981)与吉西他滨/白蛋白结合型紫杉醇标准方案的联合应用[12,18],并结合纳米递送策略克服耐药屏障[19],有望促进临床转化;同时,精确解析RRM1的SUMO化位点将为开发位点特异性干预手段奠定基础。总之,本研究通过与现有文献的对比分析,阐明了SUMO2特异性调控在PDAC吉西他滨耐药中的分子机制,补充并拓展了相关证据,为靶向SUMO通路的耐药逆转策略提供了新洞见,颇具临床转化前景。
第一作者:
费庆林,硕士研究生。
通信作者:
虞先濬,博士,主任医师,教授,博士研究生导师,复旦大学附属肿瘤医院院长。
作者贡献声明:
费庆林:完成实验,撰写并修改文章;叶龙云:参与实验,修改文章;吴伟顶:实验指导,修改文章;金凯舟:实验方案设计,实验指导;虞先濬:提出研究方案和设计思路,设计论文框架,撰写和修改论文。
[参考文献]
[1] SIEGEL R L, GIAQUINTO A N, JEMAL A. Cancer statistics, 2024[J]. CA A Cancer J Clin, 2024, 74(1): 12-49.
[2] ZENG S Y, PÖTTLER M, LAN B, et al. Chemoresistance in pancreatic cancer[J]. Int J Mol Sci, 2019, 20(18): 4504.
[3] PARK W, CHAWLA A, O’REILLY E M. Pancreatic cancer: a review[J]. JAMA, 2021, 326(9): 851.
[4] DU TOIT-THOMPSON T, LECK L, GILLSON J, et al. Overcoming therapy resistance in pancreatic cancer: challenges and emerging strategies[J]. Adv Drug Deliv Rev, 2025, 224: 115647.
[5] BIEDERSTÄDT A, HASSAN Z, SCHNEEWEIS C, et al. SUMO pathway inhibition targets an aggressive pancreatic cancer subtype[J]. Gut, 2020, 69(8): 1472-1482.
[6] SCHNEEWEIS C, HASSAN Z, SCHICK M, et al. The SUMO pathway in pancreatic cancer: insights and inhibition[J]. Br J Cancer, 2021, 124(3): 531-538.
[7] HU Y B, CHEN C B, TONG X Y, et al. NSUN2 modified by SUMO-2/3 promotes gastric cancer progression and regulates mRNA m5C methylation[J]. Cell Death Dis, 2021, 12(9): 842.
[8] LI X, DING R, WANG H, et al. Research advancements on sumoylation in gastrointestinal cancers[J]. Mol Aspects Med, 2025, 106: 101401.
[9] SANG J, YANG K, SUN Y P, et al. SUMO2 and SUMO3 transcription is differentially regulated by oxidative stress in an Sp1-dependent manner[J]. Biochem J, 2011, 435(2): 489-498.
[10] HAN Z J, FENG Y H, GU B H, et al. The post-translational modification, SUMOylation, and cancer (review)[J]. Int J Oncol, 2018, 52(4): 1081-1094.
[11] ZHU L M, CHEN G, HUANG C J, et al. SUMO3 inhibition by butyric acid suppresses cell viability and glycolysis and promotes gemcitabine antitumor activity in pancreatic cancer[J]. Biol Direct, 2024, 19(1): 74.
[12] KUMAR S, SCHOONDERWOERD M J A, KROONEN J S, et al. Targeting pancreatic cancer by TAK-981: a SUMOylation inhibitor that activates the immune system and blocks cancer cell cycle progression in a preclinical model[J]. Gut, 2022, 71(11): 2266-2283.
[13] GU Y R, FANG Y, WU X, et al. The emerging roles of SUMOylation in the tumor microenvironment and therapeutic implications[J]. Exp Hematol Oncol, 2023, 12(1): 58.
[14] HASSELLUHN M C, SCHLÖSSER D, VERSEMANN L, et al. An NFATc1/SMAD3/cJUN complex restricted to SMAD4-deficient pancreatic cancer guides rational therapies[J]. Gastroenterology, 2024, 166(2): 298-312.e14.
[15] VENA F, LI CAUSI E, RODRIGUEZ-JUSTO M, et al. The MEK1/2 inhibitor pimasertib enhances gemcitabine efficacy in pancreatic cancer models by altering ribonucleotide reductase subunit-1 (RRM1)[J]. Clin Cancer Res, 2015, 21(24): 5563-5577.
[16] LAO Y X, JIN Y R, WU S F, et al. Deciphering a profiling based on multiple post-translational modifications functionally associated regulatory patterns and therapeutic opportunities in human hepatocellular carcinoma[J]. Mol Cancer, 2024, 23(1): 283.
[17] ERDEM S, LEE H J, SHANKARA NARAYANAN J S N, et al. Inhibition of SUMOylation induces adaptive antitumor immunity against pancreatic cancer through multiple effects on the tumor microenvironment[J]. Mol Cancer Ther, 2024, 23(11): 1597-1612.
[18] KÖSE H, SCHNEEWEIS C, PUTZE P, et al. Targeting mutual dependence of phosphatidylinositol-3-kinase α/δ and small ubiquitin-like modifier signaling in pancreatic cancer[J]. Gastroenterology, 2026, 170(2): 315-329.
[19] PENG L J, LIANG Y F, GUO X N, et al. Nanomedicine breakthroughs overcoming pancreatic cancer drug resistance through precision nano-interventions[J]. Nanoscale Adv, 2025, 7(19): 5888-5904.
[20] WANG X J, YANG C X, LIU Y M, et al. SUMOylation substrate encoding genes as prognostic biomarkers in pancreatic ductal adenocarcinoma with functional assessment of SAF-B2[J]. Front Pharmacol, 2025, 16: 1532658.
编辑:莫颖
审核:王琳辉 李广涛
2026-01-06
·网易号
6-羟基黄酮
基本信息
中文名称:6 - 羟基黄酮,6 - 黄酮醇,6-HF
英文名称:6-Hydroxyflavone,6-HF,6-Hydroxy-2-phenylchromen-4-one
IUPAC 名称:6-Hydroxy-2-phenyl-4H-chromen-4-one(6 - 羟基 - 2 - 苯基 - 4H - 色烯 - 4 - 酮)
CAS 号:6665-83-4
分子式:C15H10O3
分子量:238.24(精确质量:238.06300)
EINECS 号:229-704-8
MDL 号:MFCD00016945
BRN 号:194430
重要说明
6 - 羟基黄酮是一种天然存在的黄酮类化合物,与 3 - 羟基黄酮(黄酮醇骨架)和 5 - 羟基黄酮不同,它是典型的黄酮结构而非黄酮醇结构。在自然界中可从番红花(Crocus)和印度常见的爵床科植物 Barleria prionitis Linn. 的叶子中分离得到。
物理化学性质
外观:黄色至黄绿色粉末晶体
熔点:234-236°C(lit.),清晰熔融温度 233.0-237.0°C
沸点:450.1±45.0°C(760 mmHg,计算值)
密度:1.3±0.1 g/cm³(20°C,计算值)
闪点:176.3±22.2°C(计算值)
溶解性:微溶于水,易溶于乙醇、丙酮、氯仿、DMSO 等有机溶剂
pKa:7.10±0.40(25°C,计算值)
LogP:4.25(计算值)
稳定性:密封避光,阴凉干燥处保存,避免高温与潮湿;在中性和弱酸性条件下稳定,碱性条件下易分解
来源与合成
天然来源
番红花(Crocus)的柱头和花瓣中微量存在
爵床科植物 Barleria prionitis Linn. 的叶子中含量较高
其他少数植物如某些豆科和菊科植物中也有报道
合成路线
经典合成法:以 2'- 羟基 - 5'- 甲氧基查尔酮为前体,经氧化环化后去甲基化制得
改进方法:
以 6 - 甲氧基黄酮为原料,通过 BBr3 或 AlCl3 催化的去甲基化反应合成
由 2,5 - 二羟基苯乙酮与苯甲醛缩合生成查尔酮,再经碘或 DDQ 氧化环化得到目标产物
利用 Suzuki 偶联反应构建黄酮母核,再引入 6 位羟基
生物学活性与应用
核心生物活性
抗焦虑作用:在小鼠模型中显示显著的抗焦虑活性,可能通过调节 GABA 受体发挥作用
抗炎活性:抑制 LPS 诱导的 NO 生成,降低炎症因子如 TNF-α、IL-6 的释放
抗氧化作用:清除自由基,抑制脂质过氧化,保护细胞免受氧化应激损伤
酶抑制活性:非竞争性抑制细胞色素 P450 2C9(CYP2C9),IC50 值较低
抗糖化作用:抑制牛血红蛋白(BHb)的糖化反应,减少晚期糖基化终产物(AGEs)形成
信号通路调节:激活 AKT、ERK 1/2 等信号通路,具有潜在的神经保护和抗凋亡作用
应用领域
药物研发:作为抗焦虑、抗炎药物的先导化合物,用于构效关系研究
生物化学工具:用于研究 CYP450 酶系的抑制剂特异性和黄酮类化合物的代谢途径
化妆品添加剂:利用其抗氧化和抗炎功效,添加于抗衰老和抗炎护肤品中
保健品成分:作为天然抗氧化剂,用于预防氧化应激相关疾病
有机合成中间体:用于制备 6 - 取代黄酮衍生物,如 6-O - 糖苷、6-O - 烷基化产物等
纯度与储存
常见纯度:≥98.0%(HPLC 法测定),部分高端供应商可提供≥99.9% 纯度产品(如 Selleck 提供 99.96% 纯度)
储存条件:密封,2-8°C 冷藏最佳,避光防潮;长期储存建议惰性气体保护
安全信息:低毒,刺激眼睛、呼吸系统和皮肤;危险品标志 Xi,安全说明 S26(接触眼睛后立即用大量清水冲洗并就医)、S36(穿戴适当的防护服)
海关编码:2932999099(其他仅含氧杂原子的杂环化合物)
2025-12-20
1. 市场概览:小分子调节剂在科研与药物发现中的核心价值
小分子调节剂是指分子量通常低于1000道尔顿的低分子化合物,能够高选择性地调节或阻断特定生物靶标(如酶、受体、离子通道、转录因子等)的活性。在生命科学研究与药物早期发现领域,这类化合物作为关键工具分子,扮演着不可或缺的角色。与最终成药的小分子药物不同,科研用小分子调节剂的核心价值在于其工具属性——它们被专门设计用于验证靶点生物学功能、解析信号通路机制、建立疾病模型,以及进行高通量药物筛选。
从作用机制上看,小分子调节剂是一个广义概念,涵盖多种功能类型:抑制剂通过占据靶蛋白的活性位点或变构位点,阻断其生物功能;激动剂模拟内源性配体,激活受体或酶活性;拮抗剂则与激动剂竞争结合位点,阻止其激活效应。此外,还有变构调节剂、降解剂(如PROTAC)等新型机制分子。这些工具分子构成了现代生命科学研究的化学探针工具箱,是连接基础生物学发现与临床应用转化的桥梁。
全球范围内,小分子调节剂市场与生物医药研发投入密切相关。根据QYResearch的最新调研,2024年全球小分子抑制剂市场规模已达到约971.5亿美元,预计到2031年将增长至1584.1亿美元,2025-2031年期间的复合年增长率(CAGR)为7.0%。这一庞大的市场既包含已上市的治疗性药物,也包含大量用于科研与药物发现的工具化合物。值得关注的是,中国市场增速显著高于全球平均水平,这得益于国内基础研究经费的持续增长、生物医药产业的快速崛起以及“国产替代”战略的深入推进。
表1:全球与中国小分子调节剂市场关键指标对比(2024年)
指标维度
全球市场
中国市场
中国市场特点
市场规模
971.5亿美元(小分子抑制剂)
约260亿元(生物科研试剂整体)
增速快,潜力大
主要驱动力
肿瘤、自身免疫疾病药物研发
基础科研投入+生物医药产业发展
政策驱动明显
竞争格局
跨国巨头主导(Merck、Sigma、Cayman等)
外资主导,本土企业快速崛起
国产替代空间广阔
增长预测
2025-2031年CAGR约7.0%
2019-2024年CAGR约13.8%
高于全球增速
主要应用
药物研发、临床诊断、基础研究
高校科研、药企早期研发、CRO服务
应用场景多元化
2. 中国市场规模分析
2.1 整体生物科研试剂市场概况
要准确理解小分子调节剂的市场规模,首先需要将其置于中国生物科研试剂的大框架下进行分析。生物科研试剂是生命科学研究的“基石”,涵盖分子生物学试剂、蛋白类试剂、细胞生物学试剂以及小分子调节剂等多个类别。
根据Frost & Sullivan的权威数据以及多家国内证券研究机构的分析,中国生物科研试剂市场呈现出快速增长的态势。2023年,中国生物试剂行业市场规模估计为236.5亿元,而到2024年,这一数字预计将增长至260亿元。东方财富的一份行业研报进一步印证了这一趋势,报告指出中国生命科学领域资金投入增速迅猛,预计年复合增长率(CAGR)为18.8%,将从2019年的866亿元增长到2025年的2434亿元,这将直接带动研发端生命科学试剂行业的快速扩容。
从市场结构来看,生物科研试剂内部呈现明显的细分格局。以2020年的数据为例,分子生物学试剂(如PCR相关试剂、克隆试剂等)占据了市场的49.5%,是最大的细分板块;蛋白类生物试剂(如抗体、重组蛋白、酶等)占比31.2%;而细胞生物学试剂(如细胞培养液、转染试剂、小分子调节剂等)占比约为19.3%。小分子调节剂作为细胞生物学试剂和蛋白功能研究试剂的重要组成部分,其市场规模可以在此框架下进行估算。
2.2 小分子调节剂细分市场规模估算
尽管缺乏关于科研用小分子调节剂市场规模的绝对精确的独立统计数据,但通过行业分析、企业调研和市场结构推演,我们可以对其形成可靠的定量判断。
综合多家市场研究机构的观点和产业链信息,目前业内普遍认为,小分子调节剂约占整个生物科研试剂市场的15%-25%。这一比例来源于几个关键考量:首先,在基础科研和药物发现中,小分子探针是验证信号通路和靶点功能的首选工具,需求刚性且持续;其次,随着化学蛋白组学、表观遗传学等前沿领域的发展,对特异性小分子工具的需求日益增长;最后,相较于部分标准化程度高的分子生物学试剂,小分子调节剂品类繁多、定制化程度高,附加值也相对较高。
基于260亿元的生物科研试剂总盘子和15%-25%的占比区间进行估算,可以得出,2024年中国科研与早期药物发现用小分子调节剂的市场规模大约在39亿元至65亿元人民币之间。取其中位数,约为52亿元。这个规模与国内蓬勃发展的生物医药研发(尤其是临床前研究)投入高度匹配。众多高校、科研院所、制药企业、生物技术公司和CRO/CDMO机构构成了这一市场的核心客户群。
表2:中国科研用小分子调节剂市场规模估算(2024年)
估算依据
数值/比例
数据来源/说明
中国生物科研试剂总规模(2024)
260亿元
Frost & Sullivan,中商产业研究院
小分子调节剂占比(估算)
15% - 25%
行业共识,基于产品结构与应用广度
小分子调节剂市场规模(区间)
39亿元 - 65亿元
计算得出
市场规模中位数
约52亿元
作为市场规模的参考基准
主要驱动因素
1. 生命科学研发投入增长(CAGR 18.8%) 2. 创新药研发热潮 3. 国产替代政策支持
西南证券行业研报
下游客户分布
高校/科研机构(约40%)、药企/Biotech(约35%)、CRO/CDMO(约25%)
基于供应链调研的典型分布
2.3 市场增长动力与未来展望
中国小分子调节剂市场的快速增长并非偶然,而是由多重动力共同驱动:
政策与资金驱动:国家在生命科学领域持续加大投入,各类科研基金和产业扶持政策为实验室提供了充足的经费,用于购买包括小分子调节剂在内的科研试剂。
产业需求拉动:中国生物医药产业已从“仿制为主”转向“创新引领”,本土药企和Biotech公司在新靶点发现、先导化合物筛选和优化阶段,对小分子调节剂的需求呈指数级增长。
技术迭代推动:新技术如PROTAC、分子胶、共价抑制剂等对小分子工具提出了新的要求,推动了产品品类的扩充和升级,带动了市场价值提升。
供应链安全与国产替代:在国际贸易环境存在不确定性的背景下,保障科研供应链的自主可控成为国家战略。本土试剂企业在产品质量、价格、供货速度和服务响应上的优势日益凸显,加速了进口替代进程。
展望未来,预计到2027年,中国科研用小分子调节剂市场有望突破80亿元大关。增长将不仅来自数量的增加,更来自于产品结构的升级——高附加值、高特异性的新型调节剂(如选择性降解剂、变构调节剂)占比将不断提升。
3. 产品分类体系:多维度的科学划分
小分子调节剂种类浩如烟海,科学的分类体系有助于我们理解其全貌。在实际科研和商业应用中,通常采用以下多维分类方式:
3.1 按作用机制分类(核心分类)
这是最基础、最直观的分类方式,直接反映了化合物与靶标相互作用的生物学后果。
抑制剂:最大且最核心的类别。指能降低或完全阻止靶蛋白(通常是酶)生物活性的分子。根据作用方式可分为:
可逆抑制剂:通过非共价相互作用(氢键、疏水作用等)与靶点结合,作用可逆。如大部分激酶抑制剂。
不可逆抑制剂(共价抑制剂):通过活性化学基团与靶蛋白活性位点的特定氨基酸残基形成共价键,实现持久抑制。如BTK抑制剂依鲁替尼的类似探针。
激动剂:能够结合并激活受体,产生与内源性配体相似或更强生物学效应的分子。例如,Forskolin是腺苷酸环化酶的直接激动剂,广泛应用于cAMP信号通路研究。
拮抗剂:能够结合受体,但不激活其活性,从而阻断内源性激动剂或外源性激动剂作用的分子。例如,CNQX是一种经典的竞争性AMPA/海藻酸受体拮抗剂,在神经科学研究中广泛应用。
变构调节剂:不直接结合靶蛋白的活性位点,而是结合其其他区域(变构位点),通过引起构象变化来调节活性。可分为正性变构调节剂(增强活性)和负性变构调节剂(减弱活性)。
降解剂:新兴类别,以PROTAC为代表。这类分子是双功能小分子,一端结合靶蛋白,另一端结合E3泛素连接酶,从而给靶蛋白“贴上”泛素标签,引导其被蛋白酶体降解。这是一种事件驱动的药理模式,与传统“占位驱动”的抑制剂有本质区别。
3.2 按靶点类型分类(应用导向分类)
这是药物发现和靶向研究中最常用的分类方式,直接关联疾病领域和研究方向。
激酶抑制剂:规模最大的类别。针对蛋白激酶家族,在肿瘤、炎症等领域研究至关重要。可进一步细分为酪氨酸激酶抑制剂(如EGFR、VEGFR抑制剂)、丝氨酸/苏氨酸激酶抑制剂(如AKT、mTOR抑制剂)等。QYResearch报告中将激酶抑制剂列为小分子抑制剂的首要细分类型。
GPCR调节剂:G蛋白偶联受体是最大的药物靶标家族。包括GPCR的激动剂、拮抗剂和变构调节剂,涉及神经、心血管、代谢等多个疾病领域。
离子通道调节剂:调节钠、钾、钙等离子通道的开放与关闭,主要用于神经科学、心脏电生理研究。
核受体调节剂:作用于类固醇激素受体、甲状腺激素受体等核受体,用于内分泌、代谢疾病和癌症研究。
表观遗传学靶点调节剂:针对组蛋白去乙酰化酶、组蛋白甲基转移酶、DNA甲基转移酶等,是肿瘤和神经系统疾病研究的热点。
蛋白酶体与蛋白酶抑制剂:如蛋白酶体抑制剂用于多发性骨髓瘤研究,半胱天冬酶抑制剂用于细胞凋亡研究。
其他酶类抑制剂:涵盖磷酸二酯酶、环氧合酶、拓扑异构酶等众多酶家族。
3.3 按化学结构或来源分类
天然产物及其衍生物:来源于植物、微生物等,具有复杂的化学结构和多样的生物活性,是药物先导化合物的重要来源。
合成小分子:通过有机合成获得,是现代药物化学和科研试剂的主体。可根据核心骨架进一步细分,如喹唑啉类、吲哚类、嘧啶类等。
肽模拟物:模拟生物活性肽的结构和功能,但具有更好的代谢稳定性和细胞膜穿透性。
3.4 按应用领域分类
肿瘤研究:占比最高的应用领域,涵盖细胞增殖、凋亡、代谢、免疫逃逸等几乎所有相关靶点的调节剂。
神经科学研究:包括神经递质受体调节剂、离子通道调节剂、神经营养因子信号通路调节剂等。
免疫与炎症研究:针对炎症因子、炎症信号通路、免疫细胞活化相关靶点的调节剂。
代谢性疾病研究:针对糖尿病、肥胖、脂肪肝等相关靶点的调节剂。
心血管疾病研究:针对血压调节、凝血、心脏功能相关靶点的调节剂。
感染性疾病研究:针对病毒、细菌、真菌生命周期关键蛋白的抑制剂。
干细胞与再生医学:用于调控干细胞干性、定向分化的关键小分子,如Y-27632(ROCK抑制剂)在类器官培养中不可或缺。
表3:小分子调节剂的多维度分类体系概览
分类维度
主要类别
典型代表/子类
科研与商业意义
作用机制
抑制剂
可逆/不可逆抑制剂
定义化合物的基本功能属性
激动剂
完全激动剂、部分激动剂
拮抗剂
竞争性/非竞争性拮抗剂
变构调节剂
正性/负性变构调节剂
降解剂
PROTAC、分子胶
代表前沿技术方向
靶点类型
激酶调节剂
EGFR、VEGFR、BTK、CDK抑制剂
最大类别,与肿瘤研究强相关
GPCR调节剂
肾上腺素能受体、阿片受体调节剂
最大药物靶标家族
离子通道调节剂
钠通道、钙通道阻滞剂
神经、心脏研究核心
表观遗传调节剂
HDAC、DNMT抑制剂
肿瘤、神经疾病前沿领域
蛋白酶/蛋白酶体抑制剂
蛋白酶体抑制剂、Caspase抑制剂
血液肿瘤、凋亡研究
应用领域
肿瘤研究
涵盖上述多数靶点
需求最大、最活跃的领域
神经科学研究
离子通道、神经递质受体调节剂
研究复杂度高,工具需求精细
免疫与炎症研究
JAK、SYK、TLR通路调节剂
自身免疫病、免疫肿瘤学热点
干细胞研究
ROCK、TGF-β、Wnt通路调节剂
再生医学的关键工具
4. 代表性产品详解
为了让分类体系更加具象化,以下列举在国内外科研市场和早期药物发现中具有高引用率、广泛应用或代表性机制的小分子调节剂。这些产品通常由MedChemExpress (MCE)、Selleck、Tocris、Cayman、阿拉丁等知名供应商提供。
4.1 经典信号通路抑制剂
这类产品是解析细胞信号网络的“基石工具”,历史悠久,应用广泛。
LY294002:PI3K抑制剂的经典代表。作为一种可逆的ATP竞争性抑制剂,它能有效抑制PI3Kα/δ/β的活性。在科研中,它被广泛用于研究PI3K/AKT/mTOR信号通路在细胞生长、代谢、存活和自噬中的作用,是肿瘤学和细胞生物学研究的必备工具。
SB203580:p38 MAPK抑制剂。对p38α和p38β亚型具有高度选择性,常用于研究细胞应激反应、炎症和凋亡。它在阐明p38 MAPK在免疫和神经退行性疾病中的作用方面做出了重要贡献。
PD0325901:MEK1/2抑制剂。作为RAS/RAF/MEK/ERK通路的关键下游节点抑制剂,它是研究该通路在肿瘤发生、发育和细胞分化中功能的核心工具分子。
Y-27632:ROCK抑制剂。在基础科研中用于研究细胞骨架重排、细胞迁移和收缩。其革命性应用在于干细胞领域,它能通过抑制Rho激酶信号,有效减少干细胞的凋亡(失巢凋亡),显著提高类器官的培养效率和存活率,已成为类器官和干细胞研究中的标准培养基添加剂。
4.2 新兴靶点与前沿机制代表
这类产品代表了当前药物发现的最新趋势和热点方向。
JQ1:BET溴结构域抑制剂的开创性分子。它能竞争性结合BRD4等蛋白的溴结构域,阻断其与乙酰化组蛋白的结合,从而抑制致癌基因转录。JQ1是表观遗传学研究和血液肿瘤研究中的重要探针,也引领了BET抑制剂作为抗癌药物的开发浪潮。
MLN4924:NEDD8激活酶抑制剂。它通过抑制蛋白质的neddylation修饰,导致Cullin-RING E3泛素连接酶失活,从而稳定其底物蛋白。它是研究蛋白质降解调控、细胞周期和DNA损伤应答的独特工具。
PROTAC分子(如ARV-771):靶向降解剂的代表。ARV-771是一种针对BET蛋白的双功能降解剂。与JQ1仅抑制功能不同,ARV-771能直接导致BRD2/3/4蛋白的降解,效应更持久、更彻底。这类分子不仅是研究工具,其本身也是极具前景的药物候选物,代表了“事件驱动”药理学的新范式。
IMD-0354:IKKβ抑制剂。通过抑制IKK复合物活性,阻断NF-κB信号通路的激活。该通路是炎症和肿瘤的核心枢纽,因此IMD-0354是研究炎症性疾病和肿瘤免疫微环境的重要化学工具。
4.3 神经科学核心工具
CNQX:AMPA/海藻酸受体拮抗剂。作为离子型谷氨酸受体的竞争性拮抗剂,CNQX在脑切片电生理、神经元兴奋性毒性和学习记忆机制研究中是不可或缺的工具。
MK-801:非竞争性NMDA受体拮抗剂。它通过进入NMDA受体的离子通道发挥作用,用于研究精神分裂症、神经保护和兴奋性毒性模型。
Forskolin:腺苷酸环化酶的直接激动剂。它能直接激活几乎所有亚型的腺苷酸环化酶,大幅提高细胞内cAMP水平,是研究G蛋白偶联受体(Gs型)信号转导和第二信使cAMP功能的经典工具。
4.4 细胞生物学与功能研究常用分子
Bafilomycin A1:V-ATP酶抑制剂。它能特异性地抑制液泡型ATP酶,从而阻断溶酶体和内吞体的酸化过程。广泛用于自噬研究(作为自噬流晚期抑制剂)、细胞器pH调控和病毒进入机制的研究。
MG-132:蛋白酶体抑制剂。可逆地抑制26S蛋白酶体的胰蛋白酶样活性,导致细胞内泛素化蛋白的积累。是研究蛋白质降解、内质网应激和细胞凋亡的常用工具。
表4:科研用小分子调节剂代表性产品列表
产品名称
靶点/机制
主要科研应用
典型供应商
类别
LY294002
PI3K抑制剂
PI3K/AKT/mTOR通路研究,肿瘤、代谢
MCE, Selleck, Cayman
经典通路抑制剂
Y-27632
ROCK抑制剂
类器官培养、干细胞研究、细胞迁移
MCE, Tocris, 阿拉丁
经典通路抑制剂/干细胞工具
JQ1
BET溴结构域抑制剂
表观遗传调控、血液肿瘤研究
Selleck, MedChemExpress
新兴靶点抑制剂
CNQX
AMPA/海藻酸受体拮抗剂
神经电生理、兴奋性毒性研究
Tocris, Abcam, R&D Systems
神经科学工具
Forskolin
腺苷酸环化酶激动剂
GPCR (Gs)信号、cAMP通路研究
Sigma, MedChemExpress
信号通路激动剂
Bafilomycin A1
V-ATP酶抑制剂
自噬流检测、溶酶体功能研究
MedChemExpress, Sigma
细胞生物学工具
ARV-771
BET蛋白PROTAC降解剂
靶向蛋白降解技术验证、表观遗传学
MedChemExpress
前沿降解剂
IMD-0354
IKKβ抑制剂
NF-κB炎症通路研究
MedChemExpress, Selleck
炎症通路抑制剂
5. 核心供应商竞争格局
中国科研用小分子调节剂市场是一个全球化竞争、本土化崛起的典型赛道。市场参与者主要分为两大阵营:具有全球品牌和产品线优势的跨国巨头,以及凭借快速响应、成本优势和日益提升的质量进行国产替代的本土领先企业。
5.1 跨国巨头:品牌、品类与标准的主导者
Merck (含Sigma-Aldrich、Millipore):无可争议的全球领导者。其生命科学业务板块提供数十万种小分子生物活性化合物,涵盖几乎所有已知靶点。产品以高质量、高可靠性、完善的质检文件著称,是许多实验室,尤其是高端研究和工业客户的首选。其品牌溢价高,产品线深度和广度无与伦比。2024年,默克在华营收达32.98亿美元,同比增长5.8%,其生命科学业务是主要贡献者之一。
Thermo Fisher Scientific:通过收购Life Technologies、Alfa Aesar等品牌,构建了强大的科研产品组合。其小分子调节剂产品线丰富,并注重与抗体、检测试剂盒等产品形成解决方案,为客户提供一站式服务。
Cayman Chemical:在脂类信号分子、氧化应激研究相关小分子领域具有绝对权威。其产品是研究前列腺素、白细胞三烯、大麻素受体、抗氧化通路等领域的高端标准品和工具化合物。
Tocris (现属Bio-Techne):以提供高纯度的神经科学和细胞信号研究用小分子而享誉全球。其产品目录堪称神经药理学的“教科书”,许多产品是领域内的金标准。
Selleck Chemicals:专注于小分子抑制剂,以其庞大的化合物库、清晰的靶点分类和具有竞争力的价格,在全球学术界和工业界获得了广泛认可。其网站是研究人员查找抑制剂信息的常用资源。
5.2 本土领先企业:国产替代的中坚力量
本土企业正从“模仿追随”向“创新并行”转变,竞争力体现在性价比、服务速度和特定领域的深度布局。
MedChemExpress (MCE):一家具有国际影响力的中国公司。MCE以提供大量有生物活性的小分子化合物而闻名,特别在激酶抑制剂、PROTAC降解剂等前沿领域产品线布局迅速。公司注重产品创新,每年新增大量化合物,其“化合物库”产品深受高通量筛选用户的欢迎。员工规模超过500人,业务遍及全球。
上海陶素:是一家致力于服务全球研发工作者的企业,深耕化合物筛选领域十余年,公司旗下TOPSCIENCE(陶素)品牌立足于整合多年专家资源,提供便捷的 一站式采购 体验。 实现从药物虚拟筛选到实体化合物分子供应、从商业化产品销售到个性化定制合成、从对明确靶点的分子筛选到对明确分子的多靶点筛选、从 高通量筛选 到化学结构优化的全流程服务模式。
上海阿拉丁:A股上市公司,是国内科研试剂领域的标杆企业。阿拉丁自主生产并提供数万种小分子调节剂,覆盖基础研究到药物发现的多个环节。其优势在于稳定的供应链、严格的质量控制、丰富的现货库存以及覆盖全国高校和科研机构的直销与经销商网络。公司业绩持续增长,2024年销售收入达5.34亿元,体现了强大的市场扩张能力。
上海源叶:国家级高新技术企业,专注于生化试剂和小分子抑制剂的研发与销售。其特色是提供小规格包装,灵活满足高校实验室小剂量、多品种的采购需求,客户粘性高。2024年销售额达到1.5亿元,服务客户超过5000家经销商及数万家终端科研单位。
百奥赛图等模式动物公司:它们不仅提供动物模型,也围绕其验证的靶点开发配套的药理学工具化合物,形成了从靶点验证到药效评价的闭环服务,在特定的疾病模型领域构建了独特优势。
表5:主要供应商竞争力对比分析
供应商
国别/类型
核心优势
产品特点
市场定位
Merck (Sigma)
跨国巨头
品牌信誉、品类齐全、质量全球标准、技术文档完善
超高纯度,覆盖极广,价格较高
高端科研、大型药企、合规要求严格的工业客户
Selleck
跨国专业品牌
抑制剂领域专业、信息透明、性价比优于一线巨头
专注于抑制剂,更新快,包装灵活
学术界、Biotech公司、药物筛选
MedChemExpress (MCE)
本土领先
产品创新速度快、前沿领域布局早、性价比高
大量活性化合物,PROTAC等新品多
全球学术界、工业界,注重前沿研究的客户
阿拉丁
本土上市龙头
供应链自主可控、现货库存丰富、国内渠道深入
品类覆盖广,规格多样,质量稳定
国内高校、科研院所、中小型药企和CRO
源叶生物
本土成长型
服务灵活、小包装优势、响应速度快
满足小剂量、多品种需求,定制能力强
高校实验室、初创科研团队
6. 市场驱动力、挑战与未来趋势
6.1 核心驱动力
国家战略与研发投入:“健康中国”和创新驱动发展战略下,国家自然科学基金、重点研发计划等持续加大生命科学投入,直接拉动了科研试剂需求。
生物医药产业创新浪潮:本土创新药企进入靶点前移的“深水区”,对用于靶点验证和先导化合物筛选的高质量、特异性小分子探针需求激增。
国产替代与供应链安全:在中美科技竞争背景下,实现关键科研试剂的自主可控成为共识。政策鼓励采购国产试剂,为本土企业创造了历史性机遇。
技术交叉与学科融合:化学生物学、计算化学、结构生物学的进步,使得理性设计新型小分子调节剂成为可能,不断创造新的市场增长点。
6.2 面临的主要挑战
高端产品技术壁垒:在超高选择性抑制剂、复杂天然产物衍生物、新型降解剂等领域,跨国企业在专利、工艺和技术诀窍上仍有明显优势。
质量与品牌认知:部分下游用户,尤其是高端工业客户,对国产试剂的质量稳定性和数据可靠性仍存疑虑,品牌信任度的建立需要时间。
市场竞争加剧:本土企业数量增多,同质化竞争在部分常规产品上开始出现,可能导致价格战,压缩利润空间。
人才短缺:既懂药物化学又懂生物学应用的复合型研发人才,以及具有国际视野的市场人才严重短缺。
6.3 未来发展趋势
产品向“新”与“深”发展:市场增长将从数量扩张转向质量升级。针对“不可成药”靶点(如KRAS G12C、SHP2)的变构抑制剂、PROTAC降解剂、分子胶、共价抑制剂等前沿机制产品将成为竞争焦点和价值高地。
服务化与解决方案化:单纯的化合物销售将向“产品+数据+服务”模式转变。供应商将提供化合物活性数据、使用protocol、甚至与CRO合作提供表型筛选服务,成为研究伙伴。
数字化与智能化赋能:人工智能将广泛应用于虚拟化合物库设计、ADMET性质预测和合成路线规划,极大提升新工具分子的发现效率。在线产品数据库、采购平台也将成为标准配置。
产业链纵向整合:头部试剂企业可能向上游延伸,布局关键中间体和砌块的生产,以保障供应链安全;向下游与CRO、模式动物公司合作,提供端到端的靶点验证解决方案。
全球化布局加速:领先的中国企业如MCE、阿拉丁等,将继续拓展海外市场,从“中国制造”走向“中国品牌”,参与全球竞争。
综上所述,中国科研与早期药物发现用小分子调节剂市场正处在一个规模快速扩张、结构持续升级、竞争格局重塑的黄金发展期。预计2024年市场规模在39-65亿元之间,并将在产业创新和国产替代双轮驱动下保持高速增长。
从产品维度看,市场已超越基础的信号通路抑制剂,向表观遗传、蛋白降解、变构调节等前沿领域快速演进。从竞争格局看,“外资主导、国产崛起”的态势愈发清晰,本土企业在特定赛道和客户服务上已形成差异化优势。
展望未来,这个市场的胜负手将取决于企业能否在前沿产品创新、质量体系构建、专业服务能力和全球化运营上取得突破。对于下游的科研人员和药物发现者而言,一个更加丰富、高效、可靠的国产小分子工具生态系统,必将为中国乃至全球的生命科学创新注入更强大的动力。
免责条款:本文内容仅代表作者观点,供行业交流参考,不构成任何投资或应用建议,如有侵权,请联系小编,多谢❀❀❀
蛋白降解靶向嵌合体
100 项与 上海赛徕克生物科技有限公司 相关的药物交易
登录后查看更多信息
100 项与 上海赛徕克生物科技有限公司 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2026年07月22日管线快照
管线布局中药物为当前组织机构及其子机构作为药物机构进行统计,早期临床1期并入临床1期,临床1/2期并入临床2期,临床2/3期并入临床3期
临床前
1
登录后查看更多信息
当前项目
| 药物(靶点) | 适应症 | 全球最高研发状态 |
|---|---|---|
Trigonelline hydrochloride ( Smad proteins x TGF-β ) | 矽肺 更多 | 临床前 |
登录后查看更多信息
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
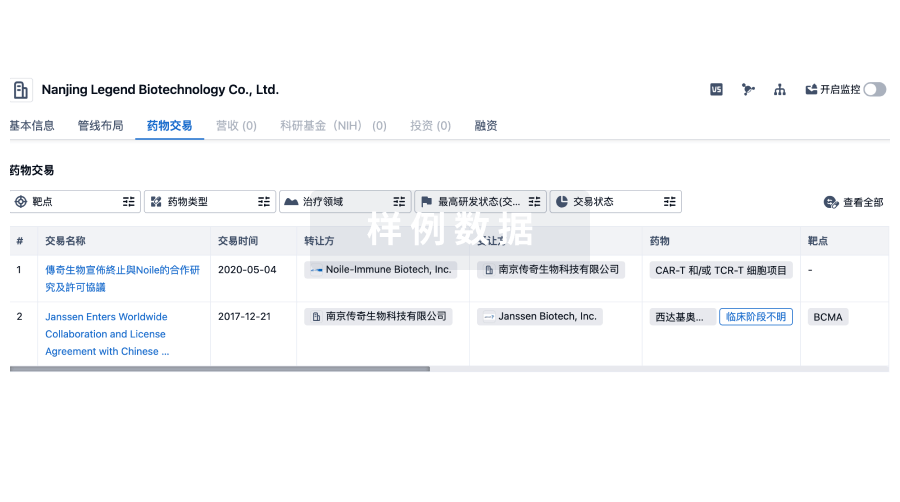
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
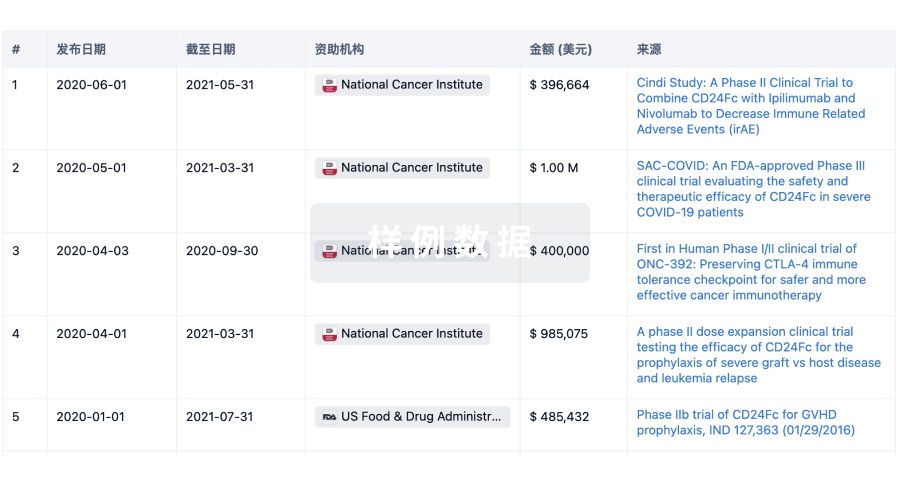
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
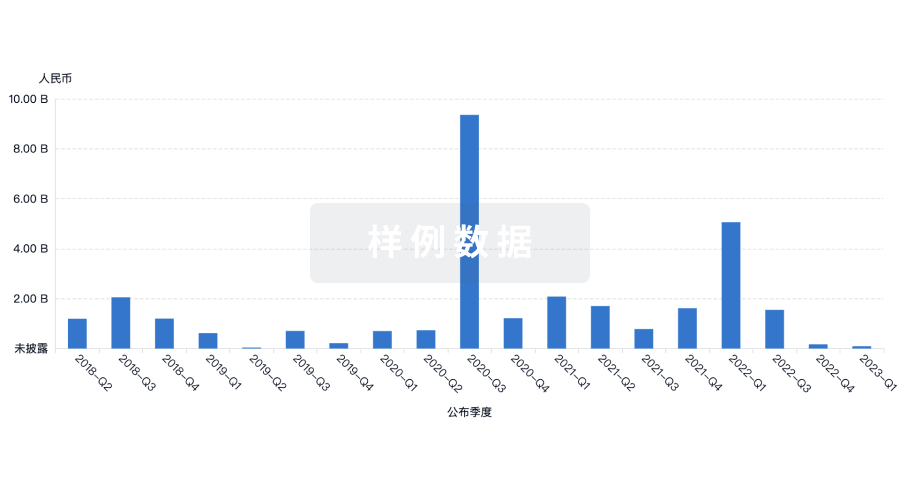
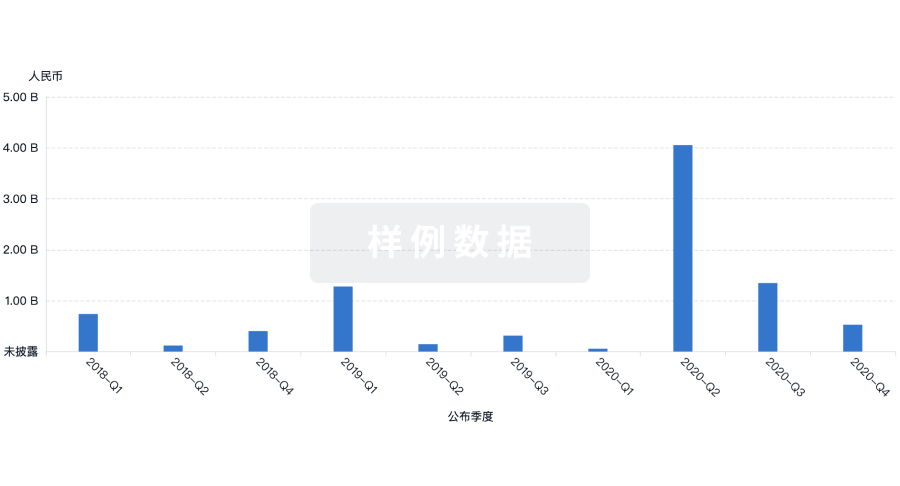
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
芽仔
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用