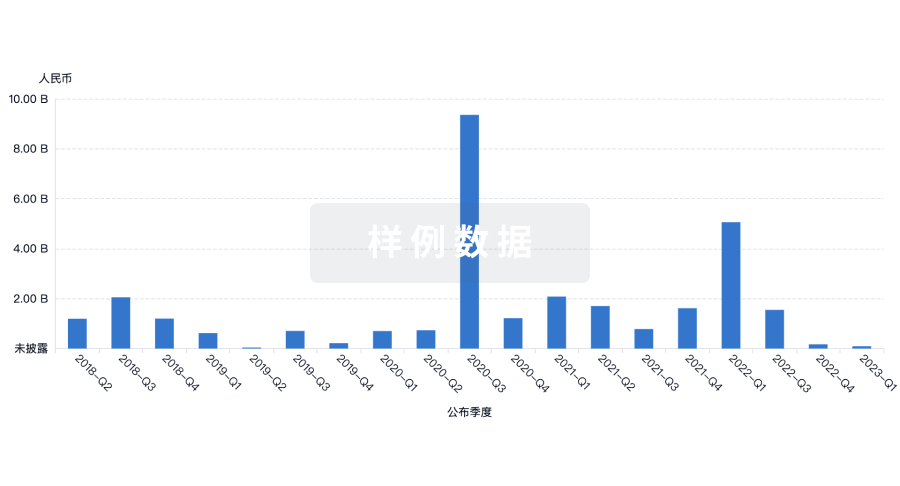
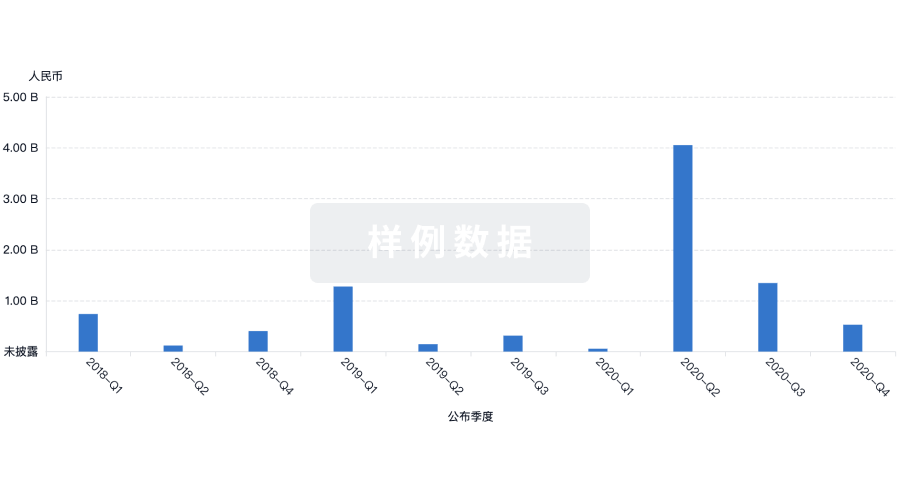
|
|
|
|
|
非在研适应症- |
最高研发阶段临床1期 |
首次获批国家/地区- |
首次获批日期- |
靶点- |
作用机制- |
|
|
|
非在研适应症- |
最高研发阶段临床申请批准 |
首次获批国家/地区- |
首次获批日期- |
靶点- |
作用机制- |
|
|
在研适应症- |
非在研适应症- |
最高研发阶段临床前 |
首次获批国家/地区- |
首次获批日期- |
评价环形RNA-HM2002注射液经心外膜心肌内注射治疗缺血性心脏病的安全性和耐受性的单中心、剂量递增、开放标签、随机、空白对照的I期临床研究
主要目的:
评价经心外膜心肌内注射HM2002注射液治疗缺血性心脏病安全性及耐受性,探索最大耐受剂量(MTD)。
次要目的:
评价HM2002注射液经心外膜心肌内注射联合冠状动脉旁路移植术治疗缺血性心脏病的初步有效性。
探索性目的:
探索HM2002注射液经心外膜心肌内注射治疗缺血性心脏病在外周血的药代动力学特征;
探索HM2002注射液经心外膜心肌内注射治疗缺血性心脏病外周血中表达蛋白VEGF-A变化的特征;
探索HM2002注射液经心外膜心肌内注射治疗缺血性心脏病的免疫原性;
探索HM2002注射液经心外膜心肌内注射治疗缺血性心脏病患者的血清生物标志物的变化。
/ Active, not recruiting早期临床1期IIT First-in-Human Pilot Study of Epicardial Circular RNA-HM2002 Injection in CABG for Ischemic Heart Failure
This exploratory, single-center, open-label clinical trial assesses the safety, tolerability, and potential efficacy of a single 5 mg dose of HM2002 injection administered via multiple-point epicardial injections for treating ischemic heart failure. Three patients aged 18-80 undergoing elective coronary artery bypass grafting (CABG) surgery will participate. Using a sentinel dosing approach, the first subject will receive HM2002, followed by a 14-day safety observation before dosing the remaining participants. If any serious adverse events or dose-limiting toxicities occur in the first subject, a Safety Review Committee will evaluate the safety data to determine whether to continue or terminate the trial.
100 项与 上海环码生物医药有限公司 相关的临床结果
0 项与 上海环码生物医药有限公司 相关的专利(医药)
2026年4月23日,由Informa Markets、上海博华国际展览有限公司主办,上海市生物医药科技产业促进中心指导的CPHI 思享会-2026 生物药创新与产业大会在上海张江海科雅乐轩酒店盛大开幕,展开为期两天的行业巅峰对话。
本次大会聚焦细胞与基因治疗(CGT)、抗体偶联药物(ADC)两大黄金赛道,精心设置1场主论坛与5大专题论坛,以In Vivo CAR-T新突破、细胞治疗、ADC药物开发与新突破、CGT工艺开发和CMC、ADC药物CMC挑战与商业化生产等热点议题为抓手,以“打通科研转化壁垒、加速创新疗法落地”为核心目标,汇聚全球顶尖院士专家、产业领袖、资本与创新力量,共话生物药前沿技术、工艺升级、CMC质控与商业化新路径,为中国生物药创新与全球化发展注入强劲动能。
明日精彩继续!赶快扫码报名☝
获得与行业大咖面对面交流的机会
(注:请实名注册参会,信息需经主办方审核)
【院士领衔 共绘生物药创新蓝图】
主论坛
洞见新机·擘画未来
主论坛由上海医药行业协会副秘书长肇晖主持,现场高朋满座、大咖云集。
俄罗斯工程院外籍院士刘中民率先带来题为《干细胞临床研究、转化与产业化》的主旨演讲,系统解析干细胞领域前沿进展与临床转化前景。过去干细胞行业存在灰色地带,缺乏相应规定。818 号令为生物医学临床研究打通了向技术转化的渠道,实现药品注册与医疗技术转化双轨并行,同时对细胞来源、应用场景和监管作出明确规定,利好产业发展。
海军军医大学附属长征医院风湿免疫科主任徐沪济教授从系统性硬化症与系统性红斑狼疮的临床研究案例切入,深入解读通用型CAR-T治疗自身免疫病的机遇与挑战,展示了通用型CAR-T细胞疗法在有效性和安全性方面的巨大潜力。
中国科学院院士、国际欧亚科学院院士陈润生围绕人工智能的过去、现在与未来发展路径,分享人工智能大模型助力药物设计、推动药物研发的应用现状,展现AI与生物医药融合的创新范式。
圆桌对话环节,围绕《从“License-out”到“全球商业化”:中国生物药企业出海的进阶挑战与资本赋能》的议题,复星医药联席总裁王兴利、诺和诺德亚太区外部创新和对外合作负责人Jenny Yang、辉瑞中国全球业务发展中国业务发展炎症与免疫领域搜索与评估及中国本土项目负责人沈蓉、百济神州全球高级副总裁单国洪、Lilly Ventures(China)Executive Director许兆颖,从Pharma并购Biotech、生物药企业出海商业化、资本赋能产业升级等角度深度碰撞,为行业发展提供创新思路。TVM Capital/ValueSeek Ventures管理合伙人谢暄晖主持了这场领军药企与投资机构齐聚的高管对话。
【首日焦点】
【In Vivo CAR-T 专场 破局转化难题】
In Vivo CAR-T新突破专场
前沿技术 破局转化难题
下午,大会首个重磅专题“In Vivo CAR-T 新突破”火热开启,成为全场焦点。上海女王之舟董事长&CEO陈立模担任专场主持人。
【报告嘉宾从左至右依次为】
自研LNP加速In vivo CAR-T药物的临床试验进展
张金晶,威斯津生物合伙人
In Vivo CAR-T和UCAR-T,谁先跑出来?
郭志刚,南京师范大学生命科学学院教授、博士生导师,南京奇迹生物科技有限公司创始人
【报告嘉宾从左至右依次为】
体内CAR-T技术专用mRNA免疫细胞自靶向递送系统tpLNP的研发
郭磊,北京百替生物创始人&CSO
基于环形RNA的体内CAR开发
杨赟,环码生物首席技术官
【报告嘉宾从左至右依次为】
当通用型CAR-T遇到In Vivo CAR-T
方宏亮,晨泰医药首席研发官
AI驱动体内CAR-T
周露,神拓生物CEO
在君联资本执行董事戚飞的主持下,星奕昂生物创始人、董事长兼首席执行官王立群、上海锋寻生物科技有限公司项目管理与业务拓展VP陈昂、北京百替生物创始人&CSO郭磊、神拓生物CEO周露、上海女王之舟董事长&CEO陈立模就“破局・协同・提速——In Vivo CAR-T转化的核心挑战与突围路径”展开圆桌讨论,直面In Vivo CAR-T转化瓶颈、协同创新、产业化提速等关键问题,为前沿技术从实验室走向临床提供务实路径。
从生物药的前沿研发到出海与商业化落地,从In Vivo CAR-T转化的核心挑战到突围路径,首日议程兼具学术高度、产业深度与落地价值,全面呈现中国生物药领域前沿硬核的创新成果。第二天议程将聚焦细胞治疗、ADC药物开发、CGT工艺与CMC、ADC药物CMC与商业化生产全链条,欢迎业内同仁明日再赴会场,共同为产业创新发展碰撞思想火花、寻找合作契机。
【4月24日会议议程】
上下滑动,查看更多
首日会场精彩瞬间
明日精彩继续!赶快扫码报名☝
获得与行业大咖面对面交流的机会
(注:请实名注册参会,信息需经主办方审核)
推荐阅读
参会指南👉就在明天丨2026生物药创新与产业大会完整参会指南
联系我们
展位申请
宓女士
电话:021-33392513
邮箱:millie.mi@imsinoexpo.com
李先生
电话:021-33392270
邮箱:BenLF.Li@imsinoexpo.com
会议合作
张女士
电话:021-33392180
邮箱:alisa.zhang@imsinoexpo.com
孙女士
电话:021-33392319
邮箱:xue.sun@imsinoexpo.com
(来源:商图药讯)
蓬勃生物主办的“2026 In Vivo X 创新峰会”将于 2026 年 5 月 19-20 日在上海盛大启幕。本次峰会以Accelerating ideas, Delivering impact为核心方向,汇聚国内外顶尖学者、产业领袖、临床 PI 及监管专家,打造覆盖全产业链的两天深度论坛,从科学前沿进展、产业难点专题研讨到商业化落地质量监管,沿脉络递进,解锁下一代CGT的创新密码。◆ 40+全球体内治疗先锋人物◆ 5大专题论坛+BD项目路演专场◆ 800+产业上下游精英汇聚深入拆解当前递送系统的突破与难点,推动in vivo新技术的开发,平衡技术创新与质量合规发展,共商产业合作与发展新范式。会议时间:5月19-20日主办单位:蓬勃生物会议地点:上海张江科学城希尔顿酒店
2026/5/19 上午大会开幕式及主旨报告大会欢迎致辞国 璋,蓬勃生物CEOThe future and clinical translation of in vivo immunotherapies Carl June, 宾尼法尼亚大学终身教授(嘉宾行程确认中)基于mRNA-LNP的体内CAR-M疗法的开发(暂定)谭蔚泓,中国科学院院士TBDCécile Bauche,Alaya.bio联合创始人兼CSO体内细胞治疗的实践和探索梁爱斌,同济大学血液肿瘤临床研究中心主任签约仪式• 圆桌对话:从in vivo CAR-T到in vivo X,体内唤醒的浪潮是否已来?戚 飞(主持),君联资本执行董事黄 浩,阿斯利康战略合作与投资发展部总监王立群,星奕昂生物董事长李雯佳,罗氏商务拓展总监徐应永,启函生物首席医学官嘉宾或议程如有变动,以现场实际为准……2026/5/19 下午分论坛1:LNP递送的进展与挑战mRNA递送技术用于在体细胞工程的研发与转化刘伟为,阿法纳生物副总经理mRNA-tLNP for in Vivo CAR-T:challenge and solutionKevin Chen,蓬勃生物 mRNA业务部负责人TBD英 博,艾博生物创始人、董事长、CEOmRNA肿瘤疫苗+CD7-tLNP in vivo CAR全球化开发夏 芳,思路迪医药副总裁LNP-mRNA 介导的体内CAR-T 研究熊长云,君健生物CEO基于mRNA靶向递送的in vivo CAR-T技术及临床进展张金晶,威斯津生物商务发展总监• 圆桌对话:新一代LNP递送技术:免疫原性控制、靶向性突破与产业化路径探索英 博,艾博生物创始人、董事长、CEO朱忠远,映恩生物创始人、董事长、CEO汤辰翔,环码生物CEO喻学亮,星锐医药副总经理孙 木,云顶新耀中国商务拓展总监嘉宾或议程如有变动,以现场实际为准……分论坛2:慢病毒载体开发的技术挑战In Vivo BCMA CAR-T细胞(ESO-T01)治疗复发/难治骨髓瘤的一期研究结果李春蕊,华中科技大学同济医学院附属同济医院血液内科党总支部书记&副主任,三级教授&三级主任医师用于体内CAR-T的慢病毒载体工艺开发的关键要点与核心挑战宣春玲,蓬勃生物LVV工艺开发部总监多维度工程化细胞疗法陈斯迪,耶鲁大学终身教授、Cellinfinity Bio创始人基于靶向慢病毒载体的体内 CAR-T江文正,恩替佰奥创始人In Vivo CAR-T generation in patients using an engineered lentiviral vector platform targeting BCMA, CD19 and CD20张永克,驯鹿生物CSO兼高级副总裁慢病毒体内CAR-T的利弊分析杨 林,博生吉医药创始人、董事长兼CSO提速与合规:病毒载体上游生产的强化实践与合规策略(暂定)默克嘉宾嘉宾或议程如有变动,以现场实际为准……2026/5/20 上午分论坛3:Beyond CAR-TIn Vivo CAR-Tumor, 免疫治疗的下一个突破性前沿:颠覆免疫细胞治疗周国庆,上海荣瑞医药创始人兼CEO碱基编辑体内递送药物的开发牟晓盾,正序生物CEO体内 CAR-M 技术及其在实体瘤治疗上的应用古宏晨,上海交通大学 Med-X 研究院副院长、教授In vivo TCR-T开发钟 时,香雪生命科学首席科学家自体TCR-T、TCR-TCE和体内TCR-T治疗实体肿瘤的机遇和挑战王江华,可瑞生物联合创始人兼首席生产官精简的环状DNA载体实现高效、安全的体内基因递送周 帆,金斯瑞生物 新型医药材料研究平台总监嘉宾或议程如有变动,以现场实际为准……分论坛4:in vivo产品质量与法规前沿tLNP技术的质量标准体系建立与浅析余 宙,优卡迪生物QC总监如何加速CGT药物开发的法规策略探讨Tina Yu,蓬勃生物法规注册部高级总监体内细胞治疗产品申报的生物安全性检测策略胡孟军,恒驭生物研发总监In vivo CAR-T药品的监管要求和CMC考量张长风,上药生物治疗质量与注册总监• 圆桌对话:全球监管视角下,in vivo产品的质控要点与监管趋势蔡昌祖(主持),夏尔巴投资投资总监陈 昂,锋寻生物VP甘蔚萍,原启生物分析方法开发部总监张长风,上药生物治疗质量与注册总监屠 冰,监管部门专家嘉宾或议程如有变动,以现场实际为准……2026/5/20 下午分论坛5:创新技术开发和递送策略通过开放式创新穿越CGT原始创新死亡谷——以体内CAR-T疗法为例谢暄晖, TVM capital/ValueSeek Ventures 管理合伙人AI赋能的体内CAR-T周 露,神拓生物创始人CREATE Medicines: Multi-immune In Vivo Cell ProgrammingDaniel Getts,CREATE Medicines CEO以LNP为载体的体内CAR-T研究(暂定)Cytiva嘉宾AAV innovation for in vivo CAR-T(暂定)马丽佳,西湖云谷智药创始人、西湖大学生命科学学院PI新一代基于CircRNA的in vivo CAR免疫治疗技术姚 璇,简接生物创始人兼CEO嘉宾或议程如有变动,以现场实际为准……BD与项目路演专场罗氏自免布局:B细胞疗法的探索与未来展望何剑奇,罗氏Business Development Project Leaderin vivo CAR-T:早期技术,前景广阔袁清慧,中信建投医药首席多家路演企业,聚焦in vivo生物医药最前沿
2026 年 4 月 13 日,上海环码生物医药有限公司宣布,旗下核心环形 RNA 创新药HM2003 注射液成功获得美国 FDA快速通道资格(Fast Track),全球临床开发进程迎来关键突破。
截至目前,HM2003 已集齐FDA 临床试验许可(IND)、儿科罕见病药物认定(RPDD)、血栓闭塞性脉管炎孤儿药资格(ODD)、快速通道资格四项重磅国际认证,形成完整的全球监管支持体系,为研发提速与商业化落地奠定坚实基础。
FDA 快速通道资格将为 HM2003 带来滚动审评、优先审评、高频监管沟通等多重审评便利,助力这款针对严重下肢缺血性病变的创新疗法更快完成临床验证,惠及全球患者。该产品依托环码生物自主研发的环形 RNA 技术平台,以高稳定性、低免疫原性为核心优势,通过促进侧支循环重建、改善患肢血供,为临床缺乏有效方案的缺血类血管疾病提供全新治疗路径。
环码生物作为全球领先的环形 RNA 疗法企业,凭借全链条自主可控的技术平台与完善专利布局,持续领跑赛道。其另一核心产品 HM2002 注射液此前亦获 FDA 快速通道资格,双核心管线同步迈入国际化加速开发新阶段,彰显公司技术成熟度与全球竞争力。
作为环码生物的战略合作伙伴,衍因科技谨向环码生物致以热烈祝贺!
2025 年 9 月,衍因科技与环码生物达成三年战略合作(相关报道),以 “数字化先行,AI 赋能” 为核心,携手推进环形 RNA 药物研发数智化升级。目前,环码生物已在研发体系中部署衍因智研云平台,应用电子实验记录本(ELN)与样品管理两大核心模块,实现实验数据标准化、结构化留存与高效追溯,夯实研发数据资产底座,提升实验室运营效率。
战略合作签约图
双方将持续深化合作,聚焦联合开发与核心业务流程紧密相关的AI 智能体,把衍因科技在 AI 大模型与智能应用的技术积累,与环码生物在 RNA 疗法领域的深厚行业经验深度融合,赋能从靶点发现到临床前研究全流程,以数智力量助力环形 RNA 创新药高效研发,早日造福全球病患。
关于环码生物
环码生物(CirCode),是全球领先的环形RNA疗法的引领者,先后荣获国家级高新技术企业、上海市专精特新中小企业等多项殊荣。公司基于自主研发的多项关键技术,打造了全链条自主可控的环形RNA技术平台,构筑了完善的全球专利保护体系,为环形RNA药物开发扫清障碍。
环码生物从未被满足的临床需求出发,储备管线覆盖多个治疗领域:传染病疫苗、心血管、自身免疫性疾病、肿瘤等。公司凭借自身优势,在产业领域备受瞩目,吸引众多知名投资机构关注,成功斩获数亿元融资,为公司发展注入强劲动力。
环码生物已与多家国际知名药企达成合作,共同探索及开发环形RNA疗法的潜力,推动更多以环形RNA技术为疗法的药物的创新。
关于衍因科技
衍因科技是中国领先的分子生物学科研平台,致力于成为全球AI for R&D Cloud领导者。衍因智研云基于AI大模型的企业级生物科研协作平台,推行“3 +1 + N”的科研AI First战略框架,集成了生物信学套件、科研知识库套件、实验室协作套件、基因尺度大模型平台和多个智能助手。目前,衍因智研云已为数百家生物医药企业、高等院校和科研机构提供服务,助力他们高效开展科研实验和数据分析,显著提升科研效率与成果质量,加速科研成果转化。
责声明:所载内容、图片来源互联网,微信公众号等公开渠道,我们对文中观点保持中立,仅供参考、交流之目的。转载的稿件版权归原作者和机构所有,如有侵权,请联系我们删除。本公众号原创文章,欢迎转载,转载时请注明出处。
更多行业资料或入交流群,欢迎添加“小助手”
衍因科技是中国领先的分子生物学科研平台,致力于成为全球AI for R&D Cloud领导者。
公司专注于开发基于AI大模型的企业级生物科研协作平台——衍因智研云,推行“3 +1 + N”的科研AI First战略框架,集成了生物信学套件、科研知识库套件、实验室协作套件、基因尺度大模型平台和多个智能助手。目前,衍因科技已为数百家生物医药企业、高等院校和科研机构提供服务,助力他们高效开展科研实验和数据分析,显著提升科研效率与成果质量。
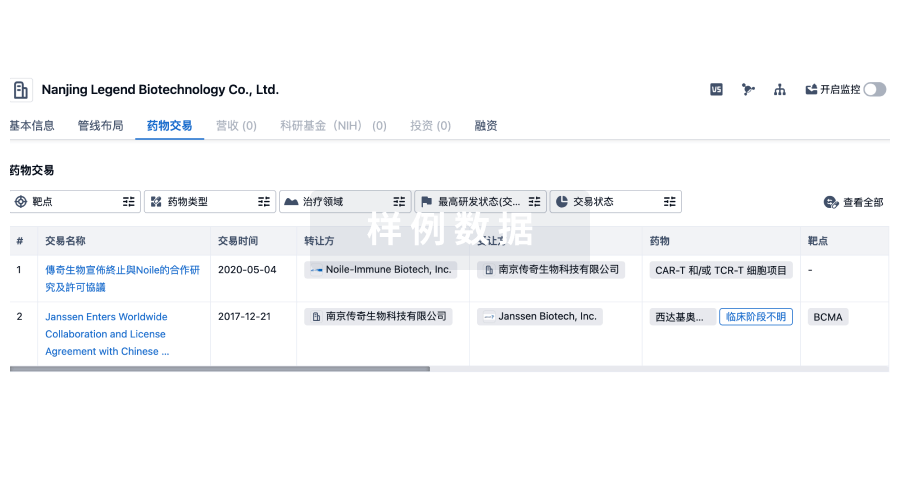
100 项与 上海环码生物医药有限公司 相关的药物交易
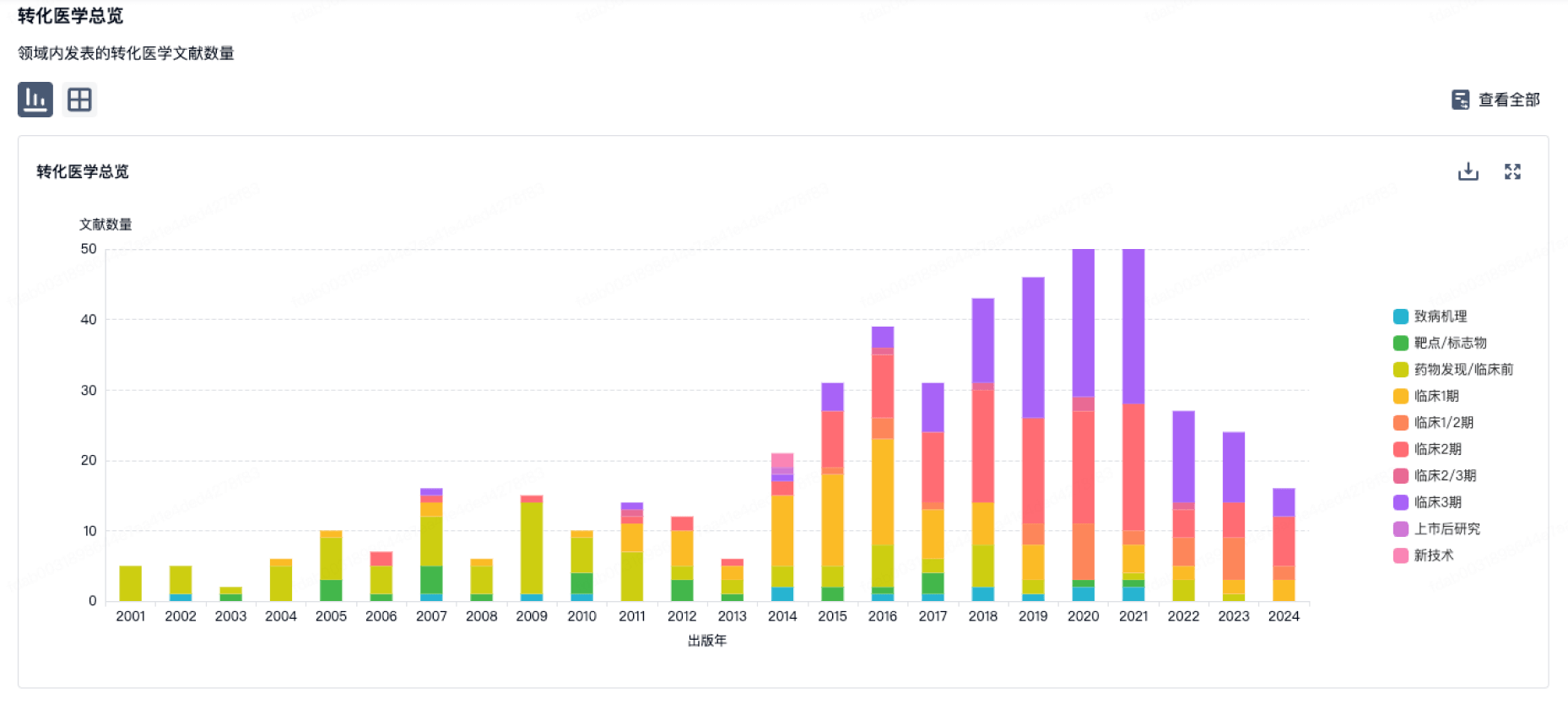
100 项与 上海环码生物医药有限公司 相关的转化医学