预约演示
更新于:2026-03-04
UNC Systems AB
更新于:2026-03-04
概览
标签
肿瘤
神经系统疾病
皮肤和肌肉骨骼疾病
蛋白水解靶向嵌合体(PROTAC)
多肽偶联核素
诊断用放射药物
疾病领域得分
一眼洞穿机构专注的疾病领域

暂无数据
技术平台
公司药物应用最多的技术
暂无数据
靶点
公司最常开发的靶点
暂无数据
| 排名前五的药物类型 | 数量 |
|---|---|
| 多肽偶联核素 | 1 |
| 诊断用放射药物 | 1 |
| 蛋白水解靶向嵌合体(PROTAC) | 1 |
| 腺相关病毒基因治疗 | 1 |
关联
3
项与 UNC Systems AB 相关的药物靶点 |
作用机制 PPT1调节剂 [+1] |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症- |
最高研发阶段临床1/2期 |
首次获批国家/地区- |
首次获批日期- |
靶点 |
作用机制 PSMA1抑制剂 |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症- |
最高研发阶段临床前 |
首次获批国家/地区- |
首次获批日期- |
靶点 |
作用机制 PRMT5 降解剂 |
非在研适应症- |
最高研发阶段临床前 |
首次获批国家/地区- |
首次获批日期- |

100 项与 UNC Systems AB 相关的临床结果
登录后查看更多信息
0 项与 UNC Systems AB 相关的专利(医药)
登录后查看更多信息
100
项与 UNC Systems AB 相关的新闻(医药)2026-03-04
2026年3月2日,艾伯维宣布其3期、随机、安慰剂对照、双盲AFFIRM研究取得了积极结果。该研究评估了risankizumab(SKYRIZI®)在中度至重度活动性克罗恩病成人患者中,皮下 (SC) 诱导治疗与安慰剂相比的疗效和安全性。
AFFIRM研究结果显示,与安慰剂组相比,risankizumab皮下诱导治疗在第12周达到联合主要终点——达成克罗恩病活动指数(CDAI) 临床缓解(55% vs.30%;p<0.0001)以及内镜应答(44% vs.14%;p<0.0001)的患者比例显著更高。在接受risankizumab皮下诱导治疗12周后获得临床缓解,继而接受12周维持治疗的患者中,67%的患者在第24周达到CDAI临床缓解,57%的患者在第24周达到内镜应答。
这项3期研究入组的患者主要为难治性人群**(65%),其中50%的患者既往使用两种或以上先进疗法治疗失败,23%的患者既往使用ustekinumab治疗失败,12%的患者既往使用Janus激酶抑制剂 (JAKi) 治疗失败。
Kori Wallace博士
艾伯维免疫学临床开发
全球副总裁
本研究评估正对的是难治性克罗恩病患者人群,其中大多数患者是既往使用先进疗法后治疗失败的人群。这些数据进一步证实了risankizumab是这类患者领先的有效治疗选择。对于克罗恩病患者而言,达到如此水平的内镜应答是一项特别有意义的成果;而对艾伯维来说,这些结果突显了我们为提升治疗标准而持续进行的创新与研究。
Millie D. Long博士
北卡罗来纳大学教堂山分校胃肠病学和肝病学系主任、公共卫生硕士、AFFIRM研究主要研究者
克罗恩病是一种复杂且常常令人身心俱疲的疾病。它不仅仅影响患者的消化系统健康,还会打乱患者的工作、人际关系和日常生活。这些在不同患者群体中取得的高内镜应答率,尤其是在那些既往无先进疗法治疗失败的患者中,展示了risankizumab皮下给药诱导治疗在克罗恩病中的有效潜力。
在12周、双盲、安慰剂对照期间,risankizumab皮下给药的安全性特征与在克罗恩病中观察到的安全性特征一致,未观察到新的安全性风险。在接受risankizumab治疗的患者中观察到的最常见不良事件为上呼吸道感染、腹痛和关节痛。risankizumab皮下给药组0.5%的患者发生了严重不良事件,而安慰剂组为3.1%。
完整结果将在即将出版的医学期刊中发表,并在未来的医学大会上分享。
关于克罗恩病
克罗恩病是一种慢性、全身性疾病,表现为胃肠道内的炎症,最常见于小肠(回肠)和结肠之间的区域,导致持续性腹泻和腹痛。克罗恩病是一种进行性疾病,会随着时间的推移而恶化导致出现威胁生命的并发症或者需要手术。由于克罗恩病的体征和症状不可预测,它给患者带来了身体、情感及经济上的巨大的负担。
关于AFFIRM研究
AFFIRM是一项全球、3期、随机、安慰剂对照、双盲研究,旨在评价risankizumab皮下 (SC) 给药作为中度至重度活动性克罗恩病成人患者的诱导治疗的疗效和安全性。联合主要终点为第12周时达到CDAI临床缓解 (CDAI<150) 的受试者百分比以及达到内镜应答的受试者百分比。共有289例患者以2:1的比例随机接受risankizumab皮下给药或安慰剂治疗。risankizumab皮下给药组和安慰剂组之间的关键人口统计学和基线特征大致平衡;65%的受试者既往使用CD先进疗法后治疗失败。本研究包括3个治疗阶段:安慰剂对照阶段A(基线至第12周),旨在评价risankizumab皮下诱导治疗的疗效和安全性;扩展阶段B(第12-24周),患者根据其第12周的临床应答接受盲态或开放性治疗;52周开放性扩展阶段C,所有患者均接受获批的risankizumab维持治疗。关于本试验的更多信息,请参见www.clinicaltrials.gov(NCT06063967)。
声明:本文所列信息涉及尚未在中国获批的产品和适应症。本文章仅作信息分享用途,不应被理解为这些产品或适应症将在中国注册或获批。
上下滑动查看更多信息
关于艾伯维
艾伯维的使命是发现和提供创新药物,解决复杂且棘手的疾病难题和未来的医学挑战。我们不懈努力,凭借在多个关键治疗领域的科研创新为人们的生活带来深远影响,包括免疫学、肿瘤学、神经科学、眼科学、病毒学、胃肠病学,以及艾尔建美学产品组合中的产品和服务。更多关于艾伯维的信息,请访问www.abbvie.com。
艾伯维在中国的总部位于上海,专注于在免疫学、肿瘤学、眼科学、麻醉学、神经科学和美学等领域为人们发展和提供创新的医疗方案。
如欲了解更多关于艾伯维在中国的信息,请浏览艾伯维中文官网www.abbvie.com.cn。
临床3期临床结果临床成功
2026-03-03
具有里程碑意义的VIVID-2研究数据显示,在治疗后一年实现无激素缓解的患者中,超过90%在三年内仍维持无激素缓解
新披露数据显示,接受安妥来利®和安妥来®治疗的克罗恩病与溃疡性结肠炎患者手术及住院率均较低,显示其有潜力从根本上改变疾病进程
目前,安妥来利®和安妥来®为唯一一款通过简便每月一次给药,即可为溃疡性结肠炎患者带来长达四年,及克罗恩病患者长达三年持续且可靠作用的IL-23p19抑制剂
最近,礼来公司(纽约证券交易所代码:LLY)公布的最新长期研究数据显示,安妥来利®和安妥来®(以上为两个剂型商品名,通用名均为:米吉珠单抗)在中重度活动性克罗恩病成人患者中,作用可持续维持三年。1这些数据来自3期VIVID-2开放标签扩展研究,并已在斯德哥尔摩举行的第21届欧洲克罗恩病与结肠炎组织(ECCO)大会上公布。此外,来自3期VIVID-1(克罗恩病)和LUCENT-3(溃疡性结肠炎)临床研究中的数据显示,在两种主要类型的炎症性肠病(IBD)患者中,接受米吉珠单抗治疗的患者住院和手术发生率均处于较低水平。2,3米吉珠单抗是目前唯一一款在溃疡性结肠炎中显示长达四年持续临床改善、并在克罗恩病中观察到最长三年持续获益,同时被证实可降低疾病相关并发症的 IL-23p19 抑制剂。
礼来执行副总裁兼免疫学总裁Adrienne Brown表示:
太多IBD患者从未获得持久缓解,炎症控制不佳引发的累积性损伤可能最终导致急诊住院或手术治疗。米吉珠单抗正在重新定义IBD的持久疾病控制。长期研究数据显示,接受米吉珠单抗治疗的患者在三年内持续维持缓解状态,且严重并发症发生率降低,这进一步凸显了其改变疾病进程的潜力。
克罗恩病的长期缓解与排便急迫感改善
在3期VIVID-1临床研究中接受米吉珠单抗治疗并在第1年达到内镜应答的患者,在VIVID-2研究中显示出长期作用。在为期三年的米吉珠单抗治疗过程中,绝大多数患者持续维持临床缓解及无激素缓解状态,并保持排便急迫感的改善。1
*指在VIVID-1研究第52周达到各指定研究终点,并在VIVID-2延长期继续接受治疗的患者;基于观察病例。上述数据亦采用改良无应答者插补法(mNRI)进行评估,相关结果见下文“VIVID临床研究项目说明”部分。
† 克罗恩病活动指数(CDAI)总分<150分。
‡ CDAI评分<150分,且在既往12周内未使用糖皮质激素。
§ 排便急迫感数值评分量表(UNRS)为以患者为评估主体的0-10分量表,用于评估排便急迫感的严重程度,其中0分表示无排便急迫感,10分表示最严重的排便急迫感。
¶ 指基线UNRS≥3分的患者。
北卡罗来纳大学教堂山分校医学副教授、医学博士、公共卫生硕士Edward Barnes表示:
对于克罗恩病患者而言,当病情未达到缓解或无法维持缓解时,不可预测的急性发作和腹痛可能持续存在;此外,当病情未得到充分控制时,排便急迫感、疲乏等持续性症状也会继续影响患者的日常生活。通过三年内持续每月给药,超过90%的患者维持无激素缓解,80%的患者排便急迫感获得缓解,使得医护人员对米吉珠单抗带来的持久作用充满信心。
最新长期研究还显示,在接受安妥来利®和安妥来®治疗并在第一年达到内镜应答的克罗恩病患者中,炎症状况持续改善,表现为炎症生物标志物(C反应蛋白和粪便钙卫蛋白)的持续降低,且这一改善状态可维持三年。1中重度活动性克罗恩病患者接受米吉珠单抗治疗的长期安全性,与其已知安全性特征一致。从治疗第一年末至第三年末报告的常见不良事件(米吉珠单抗治疗一年时实现内镜应答人群中发生率≥5%)包括新冠感染、鼻咽炎及上呼吸道感染。1
IBD患者严重疾病相关并发症的发生率长期保持低水平
作为克罗恩病三年研究结果的补充,VIVID-1(克罗恩病)和LUCENT-3(溃疡性结肠炎)临床研究的额外事后分析数据显示,接受米吉珠单抗治疗的IBD患者严重疾病相关并发症的发生率长期保持低水平。在VIVID-1研究中,前12周内,与安慰剂相比,米吉珠单抗使与克罗恩病相关的住院和/或手术发生率几乎降低了一半(发生率:16.9 vs. 30.9/100人年);在第12周至第52周期间,降幅接近70%(4.5 vs. 14.0*)。2在LUCENT-3研究中,在三年长期扩展研究期间,接受米吉珠单抗治疗的患者仅报告1例与溃疡性结肠炎相关的住院事件,未报告任何与溃疡性结肠炎相关的手术事件(发生率分别为0.1/100人年和0/100人年)。3
* 第12至第52周期间的安慰剂组发生率仅包括在第12周时达到临床应答的安慰剂组患者。
在此前已公布的克罗恩病两年及溃疡性结肠炎四年研究结果的基础上4,5,上述研究结果进一步丰富了米吉珠单抗在炎症性肠病领域的长期临床数据。
礼来正在推进米吉珠单抗的联合治疗研究,旨在实现突破性的诱导期效果,同时维持患者的长期缓解与用药安全性。相关研究包括针对溃疡性结肠炎的两项联合研究:一项为米吉珠单抗与eltrekibart(NCT06598943)联合,eltrekibart为靶向中性粒细胞介导炎症的单克隆抗体;另一项为米吉珠单抗与LY4268989(MORF-057)(NCT07186101)联合,LY4268989为口服α4β7整合素抑制剂。礼来还在推进创新科学研究,探索肠促胰素在免疫学中的潜力,并已启动COMMIT-UC(NCT06937086)和COMMIT-CD(NCT06937099 )研究,评估米吉珠单抗联合肠促胰素类疗法在成人溃疡性结肠炎或克罗恩病患者中的作用与安全性,这些患者同时患有肥胖,或超重并至少伴有一种体重相关并发症。此外,米吉珠单抗在儿童溃疡性结肠炎患者(NCT05784246)和克罗恩病患者(NCT05509777)中的临床研究正在进行中。
米吉珠单抗已获得中国国家药品监督管理局批准,用于治疗成人中重度活动性溃疡性结肠炎及中重度活动性克罗恩病,并已在全球47个国家获批上市。
关于VIVID临床研究项目
VIVID-1是一项针对中重度活动性克罗恩病成人患者的3期随机、双盲、安慰剂对照临床研究,研究周期为52周。随机分配至安妥来利®和安妥来®的患者,在第0、4和8周接受900 mg静脉(IV)输注治疗;随后自第12周起,每4周进行一次300 mg皮下(SC)注射,持续40周作为维持治疗。6
完成VIVID-1研究(包括第52周内镜检查)的受试者均可进入VIVID-2研究。在VIVID-2中,主要研究终点为评估受试者在研究第52周(累计连续治疗104周)时的长期作用,包括经克罗恩病活动指数(CDAI)评估的临床缓解情况以及内镜应答。安全性评估自受试者首次接受VIVID-2给药起进行。1
采用改良无应答者插补法分析显示,在安妥来利®和安妥来®治疗第一年达到内镜应答的患者中,82.8%在三年内维持克罗恩病活动指数(CDAI)临床缓解,81.1%维持无激素临床缓解,72.7%排便急迫感获得持续临床改善,64.0%维持排便急迫感缓解状态。1
关于LUCENT临床研究项目
两项3期临床研究评估了安妥来利®和安妥来®在中重度活动性溃疡性结肠炎成人患者中的作用与安全性,研究对象包括生物制剂初治患者以及既往接受过生物制剂或Janus激酶抑制剂(JAKi)但治疗失败的患者。LUCENT-1(诱导期)为随机、双盲、安慰剂对照研究,纳入了对糖皮质激素、免疫调节剂、生物制剂或Janus激酶抑制剂(JAKi)应答效果不佳、失应答或不耐受的患者,LUCENT-2(维持期)研究评估了在 LUCENT‑1 中对安妥来利®达到临床应答的患者中,维持治疗与安慰剂的比较。7 LUCENT‑3是LUCENT‑1和LUCENT‑2的单臂、长期、3期开放标签扩展研究,评估了溃疡性结肠炎患者在额外三年治疗期间(总治疗时间最长达四年)使用安妥来利®和安妥来®的作用和安全性。
关于安妥来利®和安妥来®
安妥来利®和安妥来®(米吉珠单抗)为 IL-23p19(白介素-23p19)拮抗剂,适用于治疗成人中重度活动性溃疡性结肠炎和克罗恩病。安妥来利®和安妥来®选择性靶向IL-23的p19亚基,抑制其信号通路。IL-23信号通路的过度激活在炎症性肠病的发病机制中起关键作用,因此抑制该通路对于控制炎症具有重要意义。8
安妥来利®和安妥来®及其给药装置底座为礼来公司(Eli Lilly and Company)拥有的商标。
参考文献
Please scroll down for more.
1.Laharie D, et al. P0563 Mirikizumab demonstrated sustained and durable long-term efficacy and favorable safety in week 52 endoscopic responders with Crohn’s disease: 3-year VIVID-2 open-label extension interim results. Journal of Crohn’s and Colitis. 2026;20(Suppl 1):jjaf231.744. https://doi.org/10.1093/ecco-jcc/jjaf231.744
2.Sands B, et al. Mirikizumab treatment reduces Crohn’s disease–related surgery and hospitalization rates: analyses from VIVID-1. Journal of Crohn’s and Colitis. 2026;20(Suppl 1):jjaf231.042. https://doi.org/10.1093/ecco-jcc/jjaf231.042
3.Magro F, et al. Mirikizumab treatment decreases ulcerative colitis–related surgery and hospitalisation rates: 4-year LUCENT studies results. Journal of Crohn’s and Colitis. 2026;20(Suppl 1):jjaf231.1300. https://doi.org/10.1093/ecco-jcc/jjaf231.1300
4.Edward L Bames et al. DDW 2025.Tu1814
5.Sands BE, et al. 2025 UEGW. MP721
6.Ferrante M, et al. Efficacy and safety of mirikizumab in patients with moderately-to-severely active Crohn’s disease: a phase 3, multicentre, randomised, double-blind, placebo-controlled and active-controlled, treat-through study. The Lancet. 2024;404(10470):2423-2436.
7.Sands, B, et al. Three-year efficacy and safety of mirikizumab following 152 weeks of continuous treatment for ulcerative colitis: results from the LUCENT-3 open-label extension study. Inflammatory Bowel Diseases, 2024;izae253, https://doi.org/10.1093/ibd/izae253
8.Omvoh. Prescribing Information. Lilly USA, LLC.
Lilly
关于商标和商品名
本文中提及的所有商标或商品名称均为礼来公司所有,若提及其他公司的商标或商品名称,其所有权则归各自所有者所有。为便于阅读,本文中提及的商标和商品名未标注®和™符号,但这并不意味着礼来公司或(在适用情况下)其他各自所有者不会在适用法律允许的最大范围内主张对这些商标和商品名的权利。我们使用或展示其他公司的商标和商品名,并非暗示与这些公司存在任何关系或意味获得这些公司的背书或赞助。
Lilly
前瞻性陈述警示声明
本新闻稿包含关于安妥来利®和安妥来®(Omvoh)治疗中重度溃疡性结肠炎及中重度克罗恩病的前瞻性陈述(该术语定义参见1995年《私人证券诉讼改革法案》),反映礼来当前的信念和预期。然而,与任何药品一样,药物在研究、开发及商业化过程中存在重大风险与不确定性。除此之外,无法保证计划中或正在进行的研究能够按计划完成、未来研究结果与迄今结果一致、安妥来利®和安妥来®能够获得额外监管批准、或安妥来利®和安妥来®能够取得商业成功。有关可能导致实际结果与礼来预期存在差异的各类风险与不确定性的详细讨论,请参阅礼来向美国证券交易委员会提交的10-K表及10-Q表文件。除法律要求外,礼来不承担更新前瞻性陈述以反映本公告发布后事件的义务。
Lilly
关于礼来公司
礼来公司是一家致力于通过科学创新改善人类健康水平,惠及全球患者的医药公司。作为医疗健康行业的领军者,礼来公司拥有近150年的历史。今天,我们的药物已帮助全球数千万人。运用生物技术、化学和基因医学的力量,我们的科学家正在积极推动新的医学进展,以应对严峻的全球健康挑战。重新定义糖尿病与肥胖疗法,减少肥胖对人体的长期影响;助力阿尔茨海默病的防治行动;为一系列威胁人类健康的免疫性疾病提供解决方案;以及将难以治愈的癌症转变为可控的疾病。礼来公司迈向健康世界的每一步,都源自于我们“致力于让数百万患者生活得更美好”的信念。这包括致力于解决全球多重挑战的创新临床试验,同时确保药物的可及性和可负担性。如果需要了解更多关于礼来公司的信息,请登录:www.lilly.com.cn。
CMAT-16089
2026-03-02
在生物科技与人工智能、大数据深度融合的今天,传统“纯生物”早已升级为“生物+计算+工程+医学”的交叉学科风口。
未来10年,生物信息学与计算生物学、合成生物学与生物工程、神经科学与认知生物学、生物医药与转化医学,将成为美国生物留学最吃香的四大方向。选对方向+选对学校,等于提前锁定高薪与高成长赛道。
01
生物信息学与计算生物学:
AI+生物大数据的“黄金赛道”
1
专业定位与核心价值
生物信息学与计算生物学是计算机科学、统计学与生命科学的深度交叉,核心是用算法和模型解析海量生物数据(基因组、蛋白质组、代谢组等),解决疾病预测、药物靶点发现、精准医疗等前沿问题。典型应用包括癌症基因组分析、肠道微生物宏基因组研究、AlphaFold2等AI蛋白质结构预测等。
2
课程体系与技能栈
核心课程:生物信息算法、基因组组装与注释、单细胞RNA-seq分析、CRISPR靶点设计、分子动力学模拟、系统生物学建模等。
硬核技能:Python数据处理(Pandas/Biopython)、Linux命令行、高通量计算(HPC/Slurm)、统计学(p值/FDR校正)、多组学整合分析。
实践资源:Rosalind生物信息编程题库、NCBI/Ensembl/UCSC Genome Browser等公共数据库、Coursera《生物信息学专项》等在线课程。
3
就业前景与薪资水平
行业分布:约55%进入生物技术公司(如23andMe、Moderna等),30%进入学术机构,15%进入科技大厂生物部门。
薪资水平:生物技术公司平均起薪约$82,000/年,学术机构博士后年薪约$60,000–$75,000,科技大厂生物部门起薪更高。
职业路径:生物信息科学家、计算生物学家、AI药物发现工程师、精准医疗算法工程师等,STEM认证优势明显,OPT实习期长达3年,国际生留美就业率超75%。
4
专业定位与核心价值
第一梯队:麻省理工学院(MIT)、斯坦福大学、哈佛大学、约翰霍普金斯大学、卡内基梅隆大学、加州大学圣地亚哥分校、华盛顿大学等。
高性价比:佐治亚理工学院、伊利诺伊大学香槟分校、加州大学洛杉矶分校等。
02
合成生物学与生物工程:
设计生命的“工程师”
1
专业定位与核心价值
合成生物学与生物工程是生物学、工程学、计算机科学和物理学的深度融合,核心是通过设计和构建新的生物部件、设备与系统,或重新设计自然生物系统,解决健康、能源、环境和材料等领域的重大挑战。典型方向包括代谢工程与生物制造、基因电路与细胞编程、基因组设计与合成、环境合成生物学等。
2
课程体系与技能栈
核心课程:分子生物学与生物化学、基因工程与DNA合成技术、生物信息学与基因组分析、生物系统建模与模拟、合成生物学伦理与政策等。
硬核技能:基因编辑(CRISPR)、DNA合成与组装、代谢通路设计、生物反应器工程、生物过程放大、数据建模与仿真。
实践资源:iGEM国际基因工程机器大赛、合成生物学实验室、生物制造中试平台等。
3
就业前景与薪资水平
行业分布:生物技术公司(药物、生物燃料、化工原料)、制药公司、农业科技公司、能源公司、政府监管机构、政策咨询与知识产权法律等。
职业路径:合成生物学家、生物工程师、代谢工程师、细胞治疗工程师、生物制造工程师、创业公司创始人等。
薪资水平:博士毕业生进入工业界起薪普遍在$80,000–$120,000/年,具备企业家精神的毕业生通过创业可获得更高回报。
4
代表院校推荐
顶尖项目:麻省理工学院(MIT)、加州大学伯克利分校、斯坦福大学、伊利诺伊大学香槟分校、杜克大学等。
特色项目:加州大学默塞德分校等院校提供合成生物学或合成分子与细胞生物学方向的硕士项目。
03
神经科学与认知生物学:
解码大脑的“终极密码”
1
专业定位与核心价值
神经科学与认知生物学是生物学、心理学、计算机科学和医学的交叉学科,核心是研究大脑的结构、功能与行为之间的关系,探索感知、记忆、语言、决策等高级认知功能的神经机制。典型方向包括细胞与分子神经科学、认知神经科学、计算神经科学、系统神经科学等。
2
课程体系与技能栈
核心课程:细胞与分子神经科学、认知神经科学、计算神经科学、系统神经科学、神经解剖学与生理学、脑成像技术(fMRI、EEG)等。
硬核技能:电生理记录、光遗传学、脑成像数据分析、计算建模、行为实验设计、统计学与编程(Python/R)。
实践资源:神经科学实验室、脑成像中心、认知与行为研究中心等。
3
就业前景与薪资水平
行业分布:学术研究机构、医院与临床研究中心、生物技术公司(脑机接口、神经疾病药物研发)、科技公司(AI与脑科学结合)、教育机构等。
职业路径:神经科学家、认知科学家、临床神经心理学家、神经工程工程师、脑机接口研发工程师、神经疾病药物研发科学家等。
薪资水平:博士毕业生进入工业界起薪约$80,000–$120,000/年,学术机构博士后年薪约$60,000–$75,000,具备AI与计算背景的毕业生薪资更高。
4
代表院校推荐
顶尖项目:哈佛大学、斯坦福大学、麻省理工学院、哥伦比亚大学、普林斯顿大学、宾夕法尼亚大学、约翰霍普金斯大学、加州大学伯克利分校、加州大学洛杉矶分校等。
04
生物医药与转化医学:
从实验室到病床的“桥梁”
1
专业定位与核心价值
生物医药与转化医学是生物学、医学、药学和工程学的交叉学科,核心是将实验室的基础研究成果转化为临床应用,开发新型药物、疫苗、诊断工具和治疗手段。典型方向包括抗体药物、mRNA疫苗、细胞与基因治疗、临床转化研究等。
2
课程体系与技能栈
核心课程:分子生物学与生物化学、药理学与毒理学、免疫学、生物制药工艺、临床试验设计与管理、转化医学研究方法等。
硬核技能:药物靶点发现、抗体工程、疫苗设计、细胞治疗技术、临床试验数据分析、监管科学与政策。
实践资源:制药公司实习、CRO(合同研究组织)项目、医院临床研究中心、生物技术孵化器等。
3
就业前景与薪资水平
行业分布:制药巨头(辉瑞、默克、诺华等)、生物科技公司(药物研发部门)、CRO公司、医院与临床研究中心、政府监管机构等。
职业路径:药物研发科学家、临床研究科学家、转化医学研究员、监管事务专员、医学事务专员、生物医药创业公司创始人等。
薪资水平:硕士毕业生起薪约$70,000–$100,000/年,博士毕业生进入工业界起薪约$90,000–$130,000/年,具备临床与监管背景的复合型人才薪资更高。
4
代表院校推荐
顶尖项目:哈佛大学、斯坦福大学、约翰霍普金斯大学、加州大学旧金山分校、麻省理工学院、北卡罗来纳大学教堂山分校、密歇根大学安娜堡分校等。
未来10年,生物学不再是传统“纯理科”,而是技术驱动的“交叉学科风口”。
生物信息学与计算生物学、合成生物学与生物工程、神经科学与认知生物学、生物医药与转化医学四大方向,将重塑生物领域的技术版图与就业市场。
选择适合的方向+顶尖院校,等于提前锁定高薪与高成长赛道。无论你是科研导向还是产业导向,都能在这四大方向中找到属于自己的“王炸组合”。
AmazingX
AmazingX辅导课程
Academy
USABO & BBO
课程独家讲义:
课程设计是基于BBO和USABO考试大纲,根据竞赛特点、重点、要点设计课程框架,高效地覆盖所有的考点。
AmazingX独家专属自研教材,将USABO和BBO的知识点对比整合,针对性攻克竞赛难题,包含全真模考题库,全方位提升与巩固知识点。
<< 滑动查看下一张图片 >>
Brain Bee
课程独家讲义:
课程设计是基于Brain Bee考试大纲,根据竞赛特点、重点、要点设计课程框架,高效地覆盖所有的考点。
AmazingX 独家专属自研教材,针对性攻克竞赛难题、赛事真题,全真模考题库,全方位提升与巩固知识点。
<< 滑动查看下一张图片 >>
课程导师介绍
牛老师:
指导国际高中8年,拥有丰富教学经验,熟悉国际教育体系各个阶段,了解学生在每个阶段遇到的瓶颈提供有针对性的解决方案,并注重课程与大学阶段的衔接。
尤其对于英国,美国教育体系十分了解,丰富的教学经验帮助学生达到中英,中美知识点转化并引导学生如何分析国际考试中解题思路,使学生快速适应国际课程的学习。
主要教授来自深国交、深大师院附属国际高中、贝塞斯、万科梅沙,香港汉基,北师大实验,人大附等学校的学生。
累计带领超过60名学生在IB生物,AP生物,化学获得满分成绩。
其中去年指导9名学生,带出了8名usabo/bbo双金奖,累计5年带出45+双金学生。
牛导师凭战绩说话,连续9年辅导学生参加Brainbee斩获一等奖(前1%参赛选手)。
更多USABO & BBO项目信息
文末扫码添加顾问老师咨询!
长按识别二维码添加AmazingX专业顾问
AmazingX
AmazingX历届战绩
Academy
USABO 18年5名学生全部获得金奖
USABO 19年8名学生7名获得金奖
USABO 20年9名学生7名获得金奖
USABO 21年10名学生8名获得金奖
USABO 22年10名学生8名学生获得金奖,2名学生获得银奖
USABO 23年指导12名学生9名学生获得金奖,3名学生银奖
USABO 24年指导16名学生9名学生获得金奖
BBO 18年4名学生全部获得金奖
BBO 19年8名学生6名获得金奖
BBO 20年9名学生7名获得金奖
BBO 21年10名学生8名获得金奖
BBO 22年指导12名学生10名学生获得金奖,2名学生获得银奖
BBO 23年指导15名学生12名学生获得金奖,3名学生银奖
BBO 24年12名学生获得金奖,4名学生银奖
Brainbee 19年80%学生获得全国一等奖,4名学生获得省状元,1名学生获得全国冠军&世界冠军
Brainbee20年全部学生获得全国一等奖,1名学生获得全国总分冠军
Brainbee21年全部学生获得全国一等奖,包括全国总分冠军
Brainbee22年全部学生获得全国一等奖,包括全国总分冠军
Brainbee23年指导出12个全国一等奖,包括全国总分冠军,季军以及3个省冠军
丘成桐中学科学奖(生物)19年1个队伍获得全球银奖(全球第2名),1个队伍获得全国二等奖
丘成桐中学科学奖(生物)20年1个队伍获得全球铜奖(全球第4名),1个队伍获得全国一等奖
奖状展示
恭
喜
大
家
战
绩
USABO竞赛历年真题包
扫码添加顾问老师!
转发推文到朋友圈集5赞
即可获得真题礼包
长按识别二维码添加AmazingX专业顾问
往期推荐 ·
三大国际生物竞赛时间轴,附小班辅导课程
【免费领取】四大生物赛事备考书单大合集
零基础友好!超全Brain Bee脑科学大赛备考攻略
生物竞赛er的上分神器,2025 iHOSA竞赛备考指南
USABO学霸冲奖的秘诀!竞赛规则/考试难点全解析
一篇吃透BBO英国生物奥赛,国际生都要看的避雷指南!
保姆级AP生物备考指南,AP生物5分拿定了!
一文读懂A-Level生物,拿A*的秘诀都在这里了!
国际生物竞赛生人手必备的自学工具,看完这篇生物竞赛不发愁!
【福利领取】国际生物竞赛学霸拿高分的秘密武器,就是这10本书!
丘成桐中学科学奖决赛名单来了!AmazingX全球前10占4席!
看完这篇备考网站&书单合集,25年Brain Bee势在必得
【资料派送】6本实用书籍助你拿下IB生物7分,书单免费领~
IB生物超全备考指南,改革后的内容变化/备考建议
BBO竞赛从入门到拿奖,只需要做好这几点!
寒假内卷计划!6部让你生物水平暴涨的生物纪录片!
IB生物学霸高分的秘籍分享!附自学网站
学霸珍藏!这5个网站带你实现Alevel生物飞跃!
入门生物竞赛前需要搞懂的4个问题!附快速记忆知识点秘诀!
AmazingX Biology
更多国际课程|留学或背景提升
扫码咨询|AmazingX专业顾问
核酸药物基因疗法
100 项与 UNC Systems AB 相关的药物交易
登录后查看更多信息
100 项与 UNC Systems AB 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2026年07月15日管线快照
管线布局中药物为当前组织机构及其子机构作为药物机构进行统计,早期临床1期并入临床1期,临床1/2期并入临床2期,临床2/3期并入临床3期
临床前
3
1
其他
登录后查看更多信息
当前项目
| 药物(靶点) | 适应症 | 全球最高研发状态 |
|---|---|---|
MS-115 ( PRMT5 ) | 前列腺癌 更多 | 临床前 |
[64Cu]NOTA-UNC-PSMA-2 ( PSMA1 ) | 肿瘤 更多 | 临床前 |
ABO-202 ( PPT1 ) | 神经元蜡样质脂褐质沉积症 更多 | 临床前 |
Dihydrexidine ( D1 receptor ) | 认知障碍 更多 | 终止 |
登录后查看更多信息
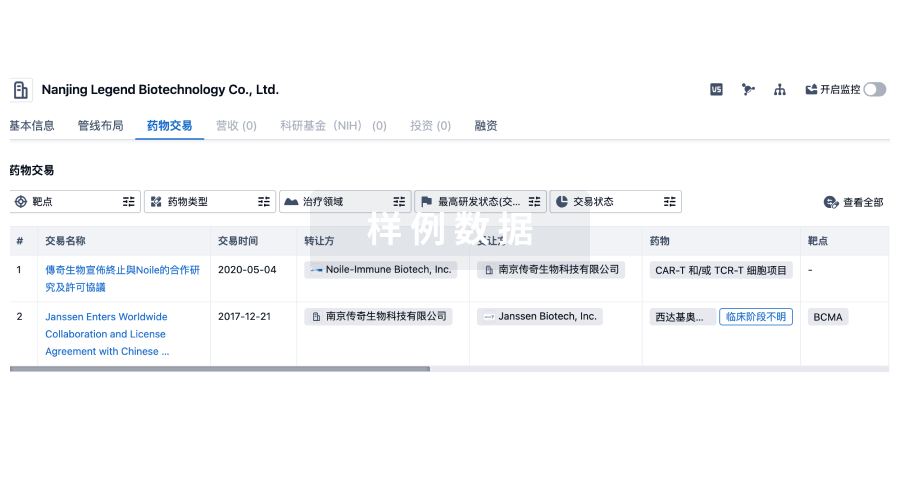
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
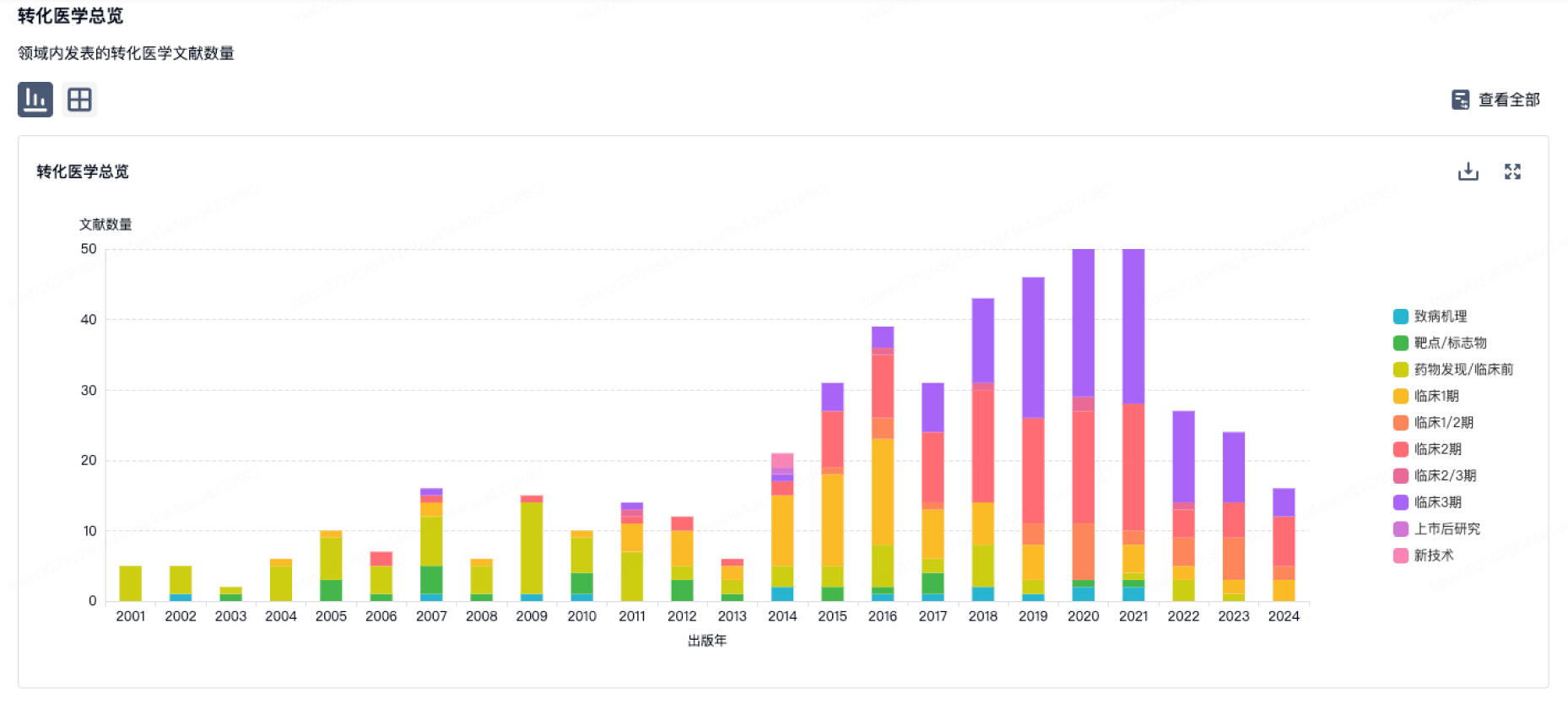
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
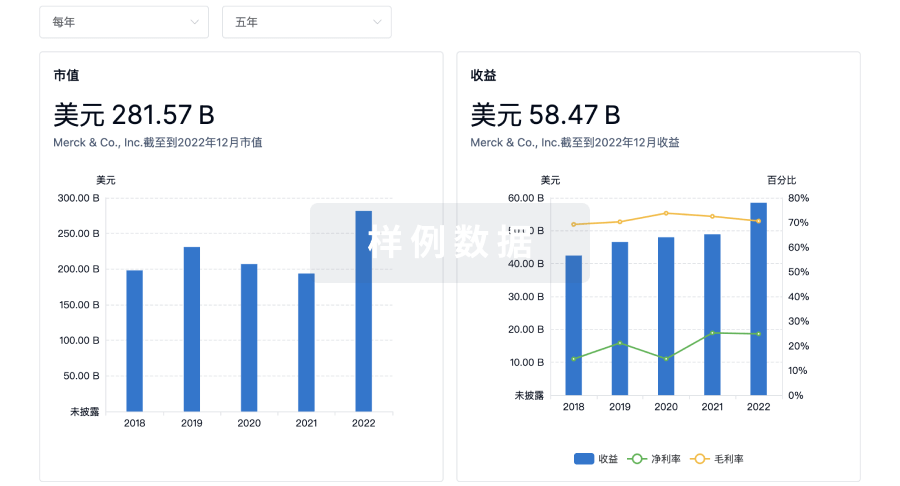
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
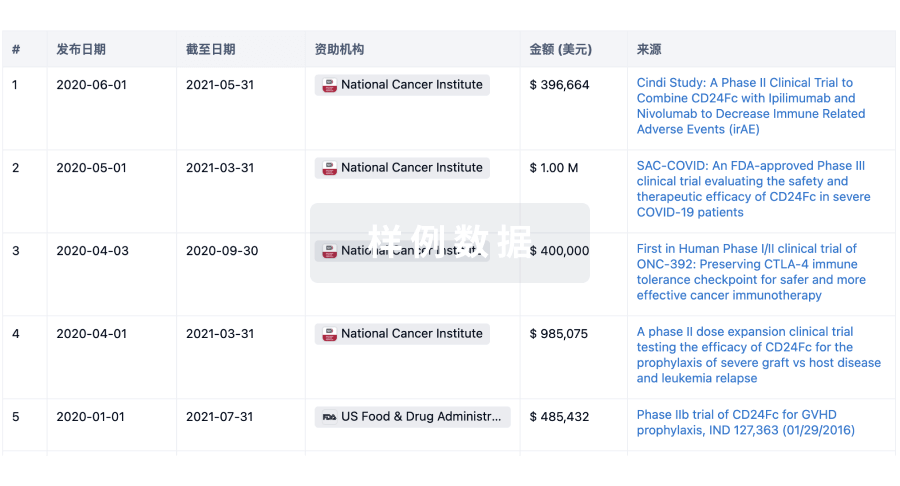
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
生物医药百科问答
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用