预约演示
更新于:2025-05-07
bFGF x VEGFR
更新于:2025-05-07
关联
靶点 |
作用机制 |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症 |
最高研发阶段 |
首次获批国家/地区 |
首次获批日期 |
靶点 |
作用机制 |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症 |
最高研发阶段 |
首次获批国家/地区 |
首次获批日期 |

靶点 |
作用机制 |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症 |
最高研发阶段 |
首次获批国家/地区 |
首次获批日期 |
100 项与 bFGF x VEGFR 相关的临床结果
登录后查看更多信息
100 项与 bFGF x VEGFR 相关的转化医学
登录后查看更多信息
登录后查看更多信息
2025-03-21Int. Journal of Clinical Pharmacology and Therapeutics
Network pharmacology of ginsenoside Rg3 in the treatment of ovarian cancer: Mechanism of action and experimental verification
Article
作者: Lu, Xizi ; Ding, Xiaofeng ; Wei, Liliang ; Zhu, Lujia ; Yun, Yi ; Zhu, Yibo ; Li, Xiang ; Ren, Boyu ; Zeng, Guishu
2025-03-01Advances in Medical Sciences
Knockout of histone deacetylase 8 gene in breast cancer cells may alter the expression pattern of the signaling molecules
Article
作者: Abdi, Mohammad ; Bahrami, Nahid
2025-01-01Journal of Animal Science and Technology
Genetic insights into avian influenza resistance in Jeju Island
chickens: the roles of Mx1 and oligoadenylate synthetase-like single nucleotide
polymorphisms
Article
作者: Jeong, Seohyun ; Yang, Hyoung-Seok ; Ko, Eun-A ; Ko, Jae-Hong ; Yang, Ju-Hee ; Kim, Young-Won ; Kang, Sung Hyun ; Tark, Dongseob ; Mun, Seong-Hwan ; Kim, Woo Hyun
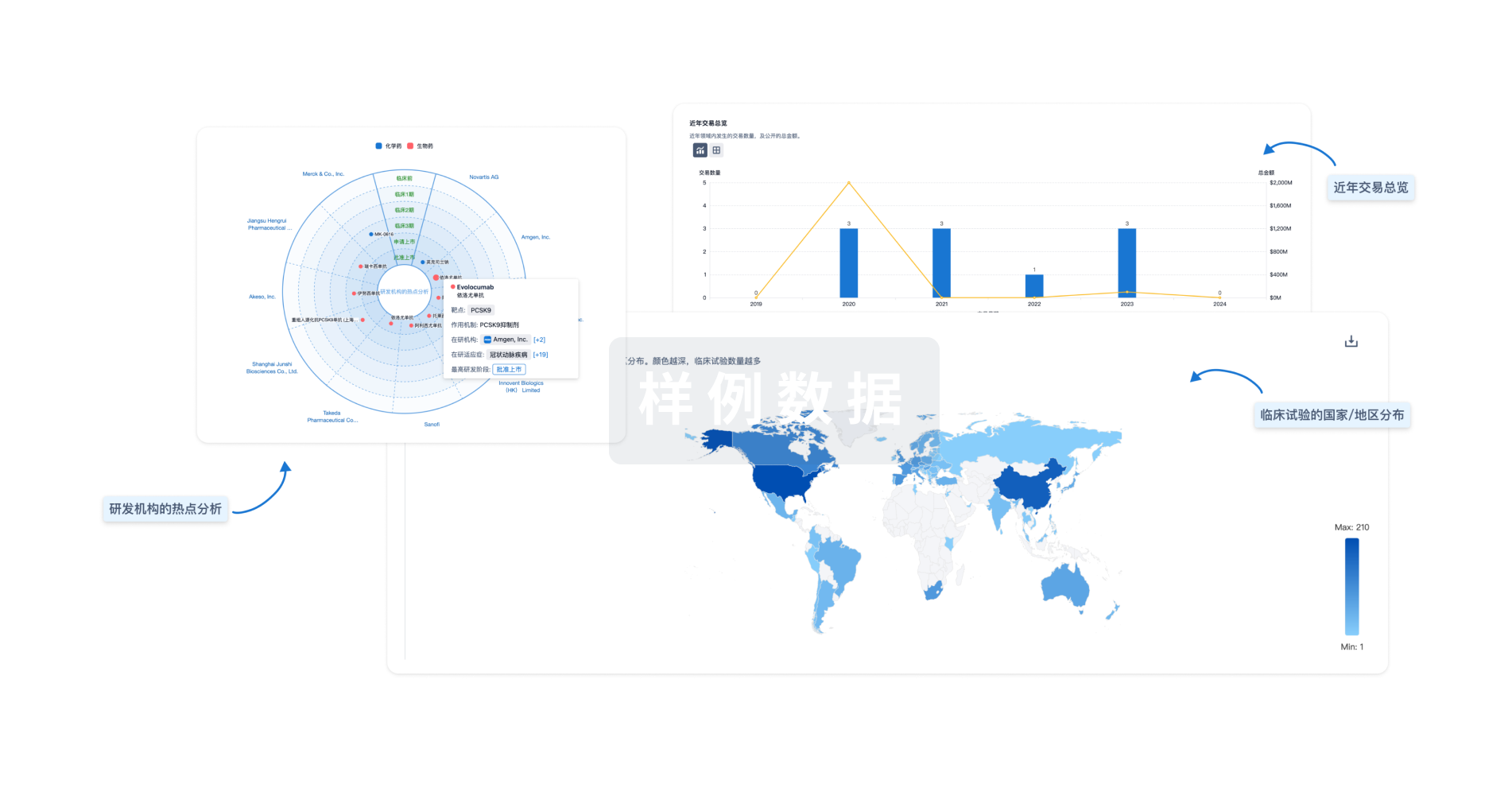
分析
对领域进行一次全面的分析。
登录
或
生物医药百科问答
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用