预约演示
更新于:2026-07-05

Imago BioSciences, Inc.
更新于:2026-07-05
概览
标签
血液及淋巴系统疾病
肿瘤
其他疾病
小分子化药
疾病领域得分
一眼洞穿机构专注的疾病领域

技术平台
公司药物应用最多的技术
靶点
公司最常开发的靶点
关联
靶点 |
作用机制 |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症 |
最高研发阶段 |
首次获批国家/地区 |
首次获批日期 |
ACTRN12623001006639
An Open-Label, Phase 1 Study to Characterize the Effects of a Moderate CYP3A4 and P-glycoprotein Inhibitor on the Pharmacokinetics of Bomedemstat (IMG-7289) in Healthy Participants
NCT05558696
A Phase 2 Multi-Center, Open Label Study to Assess the Safety, Efficacy, Pharmacokinetics and Pharmacodynamics of Bomedemstat in Patients With Polycythemia Vera (PV)
NCT05569538
Phase 2 Study to Assess the Safety and Efficacy of Bomedemstat (IMG-7289) in Combination With Ruxolitinib in Patients With Myelofibrosis

100 项与 Imago BioSciences, Inc. 相关的临床结果
登录后查看更多信息
登录后查看更多信息
2024-07-01The Prostate
Characterization of structural, biochemical, pharmacokinetic, and pharmacodynamic properties of the LSD1 inhibitor bomedemstat in preclinical models
Article
作者: Luo, Jun ; Denmeade, Samuel R. ; Antony, Lizamma ; Rienhoff, Hugh Y. ; Brennen, W. Nathaniel ; Speranzini, Valentina ; Kleppe, Maria ; Dalrymple, Susan L. ; Wang, Yuzhuo Z. ; Dias, Jennifer ; Jasmine, Sumer ; Isaacs, John T. ; Celatka, Cassandra A. ; Tapper, Amy E. ; Carducci, Michael A. ; Trock, Bruce J. ; Mandl, Adel ; Kanayama, Mayuko ; Mattevi, Andrea ; Krueger, Timothy E. G. ; Jing, Yuezhou
Abstract:
Introduction:
Lysine‐specific demethylase 1 (LSD1) is emerging as a critical mediator of tumor progression in metastatic castration‐resistant prostate cancer (mCRPC). Neuroendocrine prostate cancer (NEPC) is increasingly recognized as an adaptive mechanism of resistance in mCRPC patients failing androgen receptor axis‐targeted therapies. Safe and effective LSD1 inhibitors are necessary to determine antitumor response in prostate cancer models. For this reason, we characterize the LSD1 inhibitor bomedemstat to assess its clinical potential in NEPC as well as other mCRPC pathological subtypes.
Methods:
Bomedemstat was characterized via crystallization, flavine adenine dinucleotide spectrophotometry, and enzyme kinetics. On‐target effects were assessed in relevant prostate cancer cell models by measuring proliferation and H3K4 methylation using western blot analysis. In vivo, pharmacokinetic (PK) and pharmacodynamic (PD) profiles of bomedemstat are also described.
Results:
Structural, biochemical, and PK/PD properties of bomedemstat, an irreversible, orally‐bioavailable inhibitor of LSD1 are reported. Our data demonstrate bomedemstat has >2500‐fold greater specificity for LSD1 over monoamine oxidase (MAO)‐A and ‐B. Bomedemstat also demonstrates activity against several models of advanced CRPC, including NEPC patient‐derived xenografts. Significant intra‐tumoral accumulation of orally‐administered bomedemstat is measured with micromolar levels achieved in vivo (1.2 ± 0.45 µM at the 7.5 mg/kg dose and 3.76 ± 0.43 µM at the 15 mg/kg dose). Daily oral dosing of bomedemstat at 40 mg/kg/day is well‐tolerated, with on‐target thrombocytopenia observed that is rapidly reversible following treatment cessation.
Conclusions:
Bomedemstat provides enhanced specificity against LSD1, as revealed by structural and biochemical data. PK/PD data display an overall safety profile with manageable side effects resulting from LSD1 inhibition using bomedemstat in preclinical models. Altogether, our results support clinical testing of bomedemstat in the setting of mCRPC.
2022-10-14Clinical cancer research : an official journal of the American Association for Cancer Research
Inhibition of LSD1 with Bomedemstat Sensitizes Small Cell Lung Cancer to Immune Checkpoint Blockade and T-Cell Killing
Article
作者: Arnold, Henry U. ; Srivastava, Shivani ; Sandborg, Holly ; Martins, Renato ; Norton, Justin P. ; Hiatt, Joseph B. ; Friesen, Travis J. ; Wu, Feinan ; Sutherland, Kate D. ; MacPherson, David ; Houghton, A. McGarry ; Liao, Sheng-You ; Rienhoff, Hugh Y. ; Garrison, Sarah M.
Abstract:
Purpose::
The addition of immune checkpoint blockade (ICB) to platinum/etoposide chemotherapy changed the standard of care for small cell lung cancer (SCLC) treatment. However, ICB addition only modestly improved clinical outcomes, likely reflecting the high prevalence of an immunologically “cold” tumor microenvironment in SCLC, despite high mutational burden. Nevertheless, some patients clearly benefit from ICB and recent reports have associated clinical responses to ICB in SCLC with (i) decreased neuroendocrine characteristics and (ii) activation of NOTCH signaling. We previously showed that inhibition of the lysine-specific demethylase 1a (LSD1) demethylase activates NOTCH and suppresses neuroendocrine features of SCLC, leading us to investigate whether LSD1 inhibition would enhance the response to PD-1 inhibition in SCLC.
Experimental Design::
We employed a syngeneic immunocompetent model of SCLC, derived from a genetically engineered mouse model harboring Rb1/Trp53 inactivation, to investigate combining the LSD1 inhibitor bomedemstat with anti-PD-1 therapy. In vivo experiments were complemented by cell-based studies in murine and human models.
Results::
Bomedemstat potentiated responses to PD-1 inhibition in a syngeneic model of SCLC, resulting in increased CD8+ T-cell infiltration and strong tumor growth inhibition. Bomedemstat increased MHC class I expression in mouse SCLC tumor cells in vivo and augmented MHC-I induction by IFNγ and increased killing by tumor-specific T cells in cell culture.
Conclusions::
LSD1 inhibition increased MHC-I expression and enhanced responses to PD-1 inhibition in vivo, supporting a new clinical trial to combine bomedemstat with standard-of-care PD-1 axis inhibition in SCLC.
2020-03-24Blood advances2区 · 医学
RUNX1-mutated families show phenotype heterogeneity and a somatic mutation profile unique to germline predisposed AML
2区 · 医学
Article
作者: Mar Fan, Helen ; Brautigan, Peter ; Babic, Milena ; Natsoulis, Georges ; Krämer, Alwin ; Fine, Miriam ; Nicola, Mario ; Vakulin, Cassandra ; King-Smith, Sarah L. ; Popat, Uday ; Papaemmanuil, Elli ; Mercorella, Belinda ; Wilkins, Ella ; Feng, Jinghua ; Moore, Sarah ; Cooney, Julian ; Poplawski, Nicola K. ; Carmichael, Catherine L. ; Hiwase, Devendra K. ; D’Andrea, Richard J. ; McKavanagh, Grace ; Wei, Andrew H. ; Patton, Nigel ; Morgan, Susan ; Chong, Chan-Eng ; Godley, Lucy A. ; Forsyth, Cecily ; Schreiber, Andreas W. ; Rienhoff, Hugh Y. ; Ha, Thuong ; Wang, Paul P. S. ; Burdett, Jessica ; Jaensch, Louise ; Izraeli, Shai ; Mapp, Sally ; Horwitz, Marshall S. ; Wells, Amanda ; Bradstock, Kenneth ; Michaud, Joelle ; Lewis, Ian D. ; Fröhling, Stefan ; Butcher, Carolyn ; Homan, Claire C. ; Brown, Anna L. ; Phillips, Kerry ; Rawlings, Lesley ; Altree, Meryl ; Arts, Peer ; Li, Xiao-Chun ; Bochtler, Tilmann ; Susman, Rachel ; Hahn, Christopher N. ; Dubowsky, Andrew ; Dobbins, Julia ; Cannon, Ping ; Scott, Hamish S. ; Henry, Denae ; Currie, Mark ; Suttle, Jeffrey
Abstract:
First reported in 1999, germline runt-related transcription factor 1 (RUNX1) mutations are a well-established cause of familial platelet disorder with predisposition to myeloid malignancy (FPD-MM). We present the clinical phenotypes and genetic mutations detected in 10 novel RUNX1-mutated FPD-MM families. Genomic analyses on these families detected 2 partial gene deletions, 3 novel mutations, and 5 recurrent mutations as the germline RUNX1 alterations leading to FPD-MM. Combining genomic data from the families reported herein with aggregated published data sets resulted in 130 germline RUNX1 families, which allowed us to investigate whether specific germline mutation characteristics (type, location) could explain the large phenotypic heterogeneity between patients with familial platelet disorder and different HMs. Comparing the somatic mutational signatures between the available familial (n = 35) and published sporadic (n = 137) RUNX1-mutated AML patients showed enrichment for somatic mutations affecting the second RUNX1 allele and GATA2. Conversely, we observed a decreased number of somatic mutations affecting NRAS, SRSF2, and DNMT3A and the collective genes associated with CHIP and epigenetic regulation. This is the largest aggregation and analysis of germline RUNX1 mutations performed to date, providing a unique opportunity to examine the factors underlying phenotypic differences and disease progression from FPD to MM.
2026-01-19
·研发客
并购临床3期抗体药物偶联物免疫疗法
100 项与 Imago BioSciences, Inc. 相关的药物交易
登录后查看更多信息
100 项与 Imago BioSciences, Inc. 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2026年07月21日管线快照
管线布局中药物为当前组织机构及其子机构作为药物机构进行统计,早期临床1期并入临床1期,临床1/2期并入临床2期,临床2/3期并入临床3期
临床2期
1
1
其他
登录后查看更多信息
当前项目
登录后查看更多信息
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
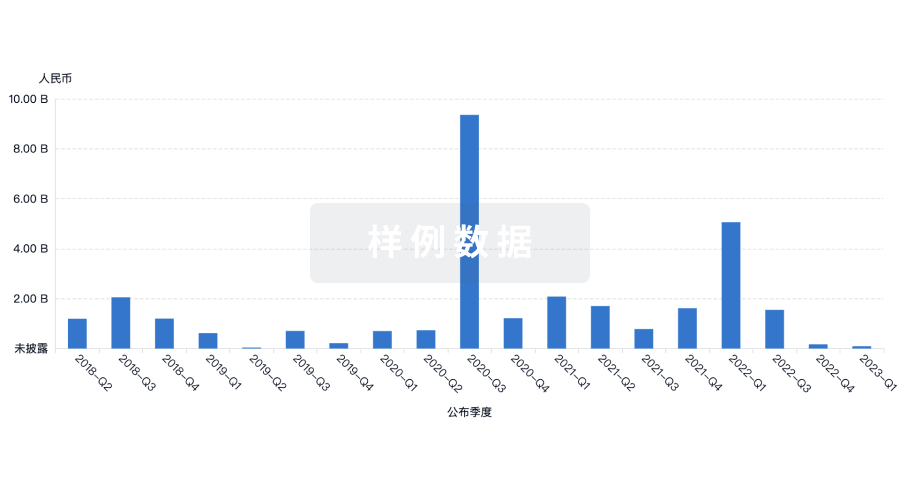
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
芽仔
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用