预约演示
更新于:2026-03-18
JAB-3068
更新于:2026-03-18
概要
基本信息
原研机构 |
在研机构- |
最高研发阶段终止临床2期 |
首次获批日期- |
最高研发阶段(中国)终止 |
特殊审评- |
登录后查看时间轴
结构/序列
分子式C22H26F2N6O2S |
InChIKeyHGYTYZKWKUXRKA-MRXNPFEDSA-N |
CAS号2169223-48-5 |
关联
3
项与 JAB-3068 相关的临床试验NCT04721223
A Phase 1b/2a, Multi-Center, Open-Label,Dose Escalation and Expansion Study to Evaluate the Safety, Tolerability, Pharmacokinetics, and Anti-tumor Activity of JAB-3068 in Combination With JS001 in Patients With Advanced Solid Tumors
To evaluate the safety and tolerability of JAB-3068 administered in investigational regimens in adult participants with advanced solid tumors.
开始日期2021-04-26 |
申办/合作机构 [+1] |
NCT03565003
A Phase 1/2a, Multi-Center, Dose Escalation/Dose Expansion, Open-Label Study to Evaluate the Safety, Tolerability, Pharmacokinetics, and Preliminary Evidence of Antitumor Activity of JAB-3068 in Adult Patients With Advanced Solid Tumors
This is a Phase 1/2a, open-label, multi-center study of JAB-3068 in Patients with advanced solid tumors.This study has two phases: dose escalation phase and dose expansion phase.
开始日期2018-11-20 |
申办/合作机构 |
NCT03518554
A Phase 1, Multi-Center, Dose Escalation, Open-Label Study to Evaluate the Safety, Tolerability, Pharmacokinetics, and Preliminary Evidence of Antitumor Activity of JAB-3068 in Adult Patients With Advanced Solid Tumors
This is a phase 1, multi-center, dose escalation, open-label study to evaluate the safety, tolerability, pharmacokinetics, and preliminary evidence of antitumor activity of JAB-3068 in adult patients with advanced solid.
开始日期2018-04-23 |
申办/合作机构 |
100 项与 JAB-3068 相关的临床结果
登录后查看更多信息
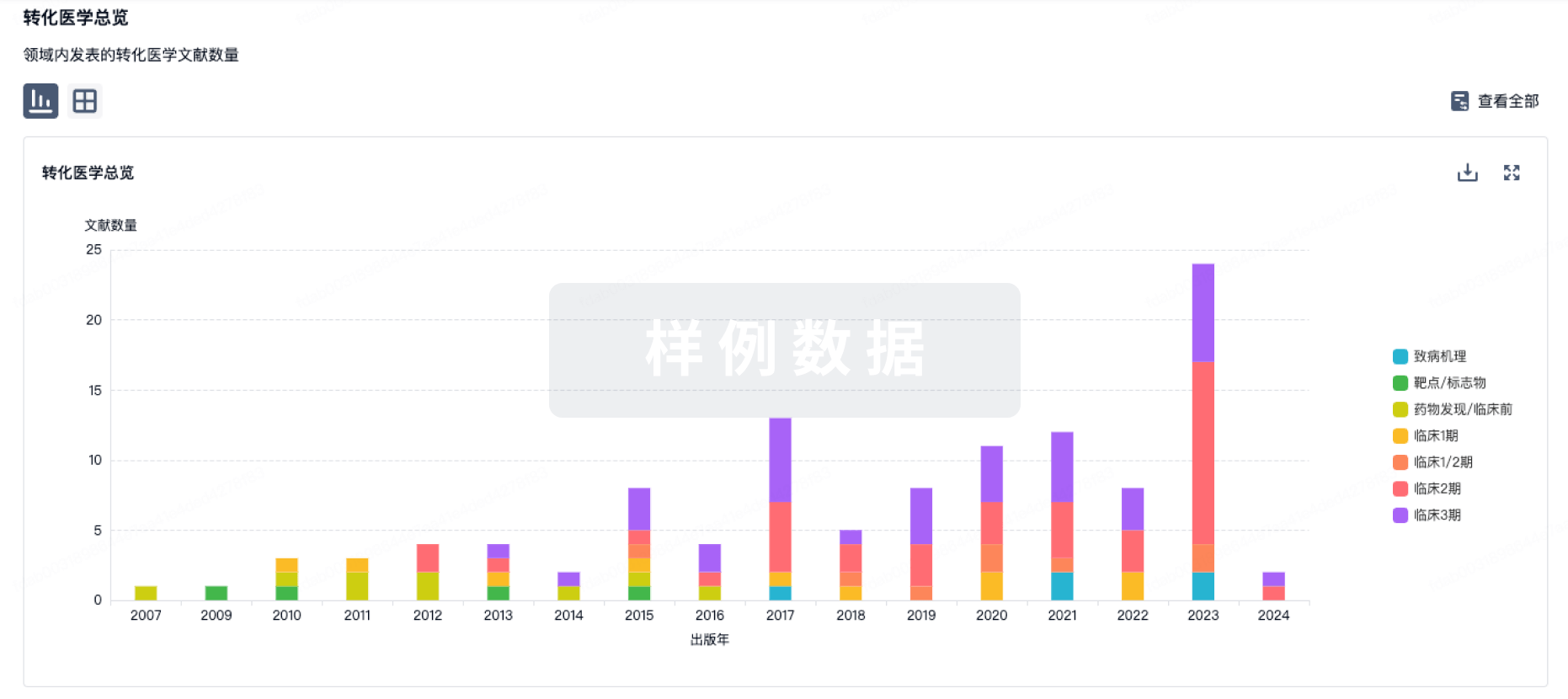
100 项与 JAB-3068 相关的转化医学
登录后查看更多信息
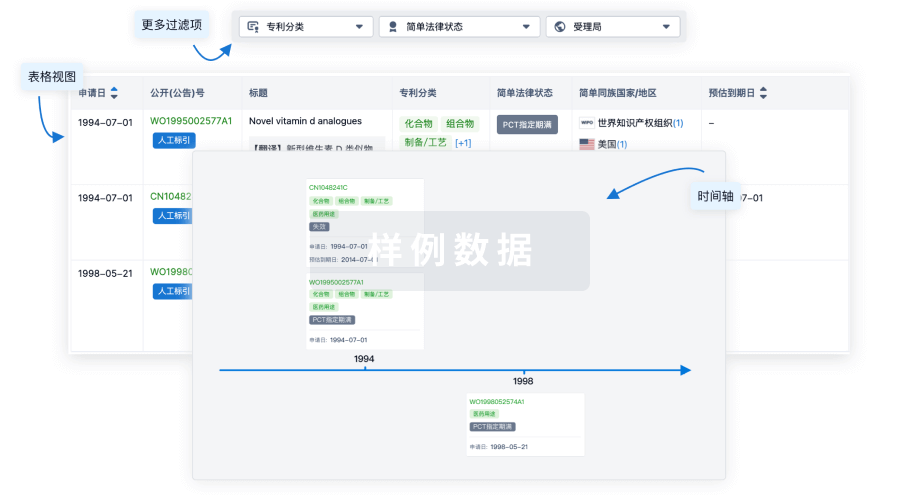
100 项与 JAB-3068 相关的专利(医药)
登录后查看更多信息
51
项与 JAB-3068 相关的新闻(医药)2026-03-17
·毕凯今选
结直肠癌的发病机制、致癌信号传导网络与靶向治疗进展
摘要
摘要
结直肠癌构成了一个突出的全球健康负担,就发病率而言,它是全球第三大常见恶性肿瘤,也是癌症相关死亡的第二大原因。结肠上皮细胞的恶性转化源于细胞内信号转导网络的复杂失调。尽管与传统治疗相比,靶向治疗已显著改善了患者生存,但驱动癌变的分子网络的复杂性继续限制着总体预后。本综述描述了控制CRC发生和发展的核心信号级联反应,重点关注该疾病的分子特征。基于大量高质量的临床前和临床证据,我们总结了目前可用的靶向药物,并批判性地评估了它们潜在的作用机制、临床疗效以及在当代治疗格局中的固有局限性。此外,我们讨论了免疫检查点抑制剂的最新进展以及对肿瘤微环境的更深入理解如何影响全球临床指南,并揭示了有前景的新靶点和组合策略。总之,对致癌信号传导通路认识的不断扩展,正在指导新疗法的开发,并使得识别适合药物干预的关键元素成为可能。最终,本综述旨在支持合理设计精确和个性化的治疗策略,以改善CRC预后。
引言
结直肠癌对全球健康构成了巨大且日益沉重的负担,是全球第三大常见恶性肿瘤和第二大癌症死亡原因。从1990年到2019年,全球年度CRC发病率急剧上升,从84万例激增至217万例,增幅超过150%,而相关死亡人数从52万增加到109万。根据发布的2022年统计数据,全球CRC发病率达到估计193万新病例,该疾病导致全球超过90万人死亡。根据2022年世界卫生组织和中国国家癌症中心的数据,中国记录了517,100例新发CRC病例(占全球总数的26.8%)和240,000例死亡(占26.5%)。作为该国第二大常见和第四大致命癌症,它构成了重大的预防挑战。根据“癌症统计学,2026”报告,预计2026年美国CRC将新增158,850例病例和55,230例死亡。这种风险因已确定的因素而加剧,包括生活习惯、饮酒和肠道微生物群失调。预计到2030年,全球新的CRC病例将超过220万,总体疾病负担预计将增加60%。
CRC通过遗传和环境因素的复杂相互作用而发展,汇聚于关键的肿瘤生长和进展通路。关键的分子驱动因素包括腺瘤性息肉病、KRAS、BRAF和TP53基因的突变,这些突变异常激活Wnt、表皮生长因子受体和PI3K/AKT信号级联反应。与染色体不稳定性、微卫星不稳定性和CpG岛甲基化表型相关的改变进一步促进了肿瘤发生。这些机制的复杂性和相互依赖性深刻影响着CRC的发生和进展以及药物治疗的疗效。
考虑到CRC的巨大负担,开发和实施有效的治疗方法是一项紧迫的优先事项。过去二十年见证了针对CRC分子发病机制的重大研究,并且这些研究已急剧加速。这一进展直接迎来了靶向治疗的时代,始于2004年美国食品药品监督管理局批准西妥昔单抗和贝伐珠单抗。此后,一系列靶向药物相继获批,目前还有更多药物正在研究中。与医学科学的持续进步一起,这些发展推动了更精确和个性化治疗方法的出现,导致患者生存率显著提高。当代管理策略范围从手术、化疗和放疗等常规治疗,到更先进的模式,如靶向治疗、免疫治疗和肠道菌群调节。本研究探讨了CRC治疗的分子演变,重点关注靶向治疗的机制、影响和未来方向。
分子发病机制:靶向治疗的基础
结直肠癌的起源与演变
开发有效的CRC靶向治疗需要更深入地了解其起源和进展。CRC是一种进行性和异质性疾病,通过一系列细胞和分子变化经历多步骤发展,从正常的肠道上皮细胞经过癌前病变进展为浸润性癌,最终发展为转移性疾病。这种进展由遗传突变、表观遗传修饰和失调的信号传导通路的顺序积累驱动,这些共同破坏了细胞稳态、增殖和凋亡。临床上,CRC的发病机制遵循三种不同的进化途径:传统的腺瘤-癌序列、锯齿状途径和炎症驱动的途径。每条途径都以独特的起始事件、特定的分子改变和病理进展模式为特征(图1)。致病菌等外部因素也可能促进CRC的发生。
腺瘤-癌序列
腺瘤-癌序列约占散发性CRC的65-70%,遵循一个明确的进展过程:正常上皮-早期腺瘤-小腺瘤(低级别异型增生)-大腺瘤(高级别异型增生)-浸润性腺癌。这一序列基本上由CIN驱动,导致结构和数目染色体异常,从而促进驱动基因改变的积累。APC的缺失使β-连环蛋白破坏复合物失稳,导致β-连环蛋白在细胞质和细胞核内稳定和积累。这随后激活靶基因(例如c-Myc、细胞周期蛋白D1),促进不受控制的细胞增殖和早期腺瘤的形成。同时,存在于约40%的CRC中的癌基因KRAS的激活突变,通过组成性激活MAPK/ERK通路促进腺瘤生长,而肿瘤抑制基因TP53的失活则促进腺瘤向CRC的进展。CIN阳性的CRC亚型主要通过腺瘤-癌途径发展。这一序列是CIN阳性CRC的主要驱动力,该亚型与侵袭性临床行为和高发远处转移相关。
图1. 关于结直肠癌发病机制的三种主要途径的图示(基于上下文推断)。图中应描绘了传统腺瘤-癌序列、锯齿状途径和炎症驱动途径,并标注了各途径的关键分子事件,如APC突变、KRAS突变、BRAF突变、CIMP和MSI状态等。
对这些分子改变的认识已直接转化为成功的临床疗法。靶向EGFR的单克隆抗体,如西妥昔单抗和帕尼单抗,通过抑制MAPK通路,成为转移性KRAS野生型CRC的标准治疗方案。值得注意的是,赋予这些抗EGFR药物耐药性的KRAS突变现在可以被直接靶向:KRAS G12C特异性抑制剂(例如索托拉西布和阿达格拉西布)最近已被批准用于携带这种特定突变的患者。此外,起始性的Wnt/β-连环蛋白通路正在被积极研究。像β-连环蛋白/TCF相互作用抑制剂(例如CWP232291)和Porcupine抑制剂(例如WNT974)这样的临床前药物显示出有前景的疗效,并且目前正在BRAF突变的转移性CRC的临床研究中推进。
锯齿状途径
除了腺瘤-癌序列,锯齿状途径(占CRC的15%-20%)是另一个主要的替代分子机制,主要由表观遗传异常驱动。这种表观遗传驱动的进展从锯齿状息肉演变而来:正常上皮-增生性息肉-无蒂锯齿状腺瘤(SSA,关键癌前病变)-锯齿状腺癌。与传统序列不同,锯齿状途径的独特特征在于两个标志性的早期分子事件:致癌性BRAF突变和CIMP。研究还发现S100A14可作为结直肠锯齿状通路的潜在生物标志物。
存在于约80%的锯齿状病变中的BRAF V600E突变的激活是锯齿状途径中的关键起始事件。该突变组成性刺激MAPK/ERK信号轴,介导生存和生长信号,确保增殖并抑制程序性细胞死亡。同时,以肿瘤抑制基因启动子(例如MLH1、CDKN2A和RUNX3)中CpG岛的广泛高甲基化为特征的CIMP,诱导转录沉默,破坏细胞周期控制和DNA错配修复。这种双重分子改变(BRAF突变 + CIMP)驱动了从增生性息肉到SSA和浸润性癌的进展。CIMP与此途径密切相关:大约75%的SSA和90%的锯齿状腺癌表现出CIMP-High状态。这一区别,连同APC突变的罕见发生以及高频出现的微卫星不稳定性-High状态(通常由甲基化介导的MLH1沉默导致),定义了锯齿状CRC独特的生物学特性。
炎症驱动的途径
第三种关键机制,炎症驱动的途径,占CRC的5%-10%,与慢性肠道炎症密切相关,特别是在被诊断患有炎症性肠病(包括溃疡性结肠炎和克罗恩病)的个体中。长期炎症通过反复的上皮损伤和修复循环破坏粘膜稳态。这导致了一个促癌的微环境,其由三个关键特征定义:升高的活性氧物种、增加的活性氮物种以及促炎性细胞因子的环境。这些介质诱导严重的氧化应激和DNA损伤,加速遗传和表观遗传修饰的逐渐积累。整个过程促进了从慢性炎症到异型增生,并最终发展为colitis相关CRC的进展。
炎症小体是关键性的胞质免疫复合物,响应细胞应激,介导炎症和细胞死亡。在各种colitis相关的CRC中,可以观察到炎症小体的激活以及白介素-1β和IL-18等细胞因子的释放。诸如NLRP3、NLRP1、NLRP6和Pyrin等感应器被激活,以发挥针对这种特定CRC亚型的保护作用。其他炎症小体感应器(例如AIM2、NLRC4、NAIP)也独立发挥作用。一个关键的致癌机制涉及转录因子SIX4,它在肠炎-腺瘤-癌序列中被上调。SIX4被IL-6/STAT3信号传导激活,随后与c-Jun相互作用,建立一个放大IL-6转录并维持慢性炎症的正反馈回路。此外,SIX4通过诱导ΔNp63表达和激活癌症干性通路促进CRC进展。在治疗上,靶向SIX4可减轻colitis和腺瘤形成。与分子数据一致,一项人群研究(NHANES 2001-2020)发现,炎症指标,特别是升高的中性粒细胞与淋巴细胞比值结合低的淋巴细胞与单核细胞比值,与增加的CRC风险相关。此外,低的LMR、高中性粒细胞与血小板比值和高的系统性免疫炎症指数与更差的临床结局相关,强调了它们作为非侵入性预后生物标志物的潜力。
结直肠癌的肿瘤微环境
TME构成一个复杂的生态系统,其中异质性的细胞组成与非细胞成分积极互动,关键性地调节癌症进展和抗肿瘤免疫。TME内的细胞和非细胞成分共同塑造疾病进展。这些包括恶性细胞、浸润的免疫细胞、可溶性因子(例如细胞因子)和细胞外基质。CRC微环境中的免疫细胞——包括肿瘤相关巨噬细胞、髓源性抑制细胞、肿瘤浸润T细胞、肿瘤相关中性粒细胞、调节性T细胞和成纤维细胞——关键地参与疾病发病机制。由髓样细胞、T细胞和成纤维细胞产生的细胞因子,如白介素-1、IL-11、IL-22和肿瘤坏死因子,通过激活核因子κB等信号传导通路,促进CRC进展。研究发现,TAMs中二肽基肽酶VII(DPP7,也称为DPP2)的表达在CRC中升高。在多个临床队列中,该基因的高表达与转移和较低的生存率相关。进一步的研究揭示,DPP7通过增强脂肪酸氧化和增加三磷酸腺苷(ATP)产生来促进TAMs的M2极化,从而加剧CD8+ T细胞耗竭并产生免疫抑制。对于MDSCs,它们通过Gasdermin C(GSDMC)介导的肿瘤细胞焦亡促进趋化因子的表达,进一步招募MDSCs到肿瘤微环境,从而促进CRC进展。进一步的研究表明,在CRC中,微生物代谢物4-羟基苯乙酸通过激活JAK2/STAT3信号传导通路促进肿瘤进展。这种激活导致CXCL3的转录上调,进而促进多形核髓源性抑制细胞(PMN-MDSC)招募和浸润到肿瘤微环境中。积累的PMN-MDSC随后抑制CD8+ T细胞的抗肿瘤活性,从而在体内促进CRC进展。Tregs被细胞因子(CCL17和CCL20)招募到肿瘤微环境中发挥免疫抑制作用。然而,最近的研究表明,SIRT1促进CRC细胞中CX3CL1的上调,从而增强Treg功能并抑制抗肿瘤免疫。研究还发现,Treg细胞的表型和功能稳态是通过乳酸通过Foxp3依赖的方式调节RNA剪接来维持的。升高的乳酸浓度抑制T细胞和自然杀伤细胞中活化T细胞核因子的活性,削弱干扰素-γ的产生,从而促进免疫逃逸。
在一项评估瑞戈非尼联合ICIs(特别是信迪利单抗)作为MSS mCRC挽救性治疗的II期、开放标签、单臂队列研究中,该方案显示出中位总生存期(OS)为14.1个月,客观缓解率(ORR)为21.4%,疾病控制率(DCR)为63.1%。RAS/RAF野生型肿瘤患者获得更大的临床获益,中位OS达到23.3个月。联合治疗耐受性良好,安全性可控。该研究表明,这种方案为MSS mCRC患者提供了一种有前景的后线治疗选择。另一项独立研究检查了实体瘤患者中序贯与同步施用ICIs和放疗的治疗结果和安全性概况。结果表明,在放疗后施用ICIs显著提高了OS和无进展生存期(PFS)。相比之下,同步治疗方法并未提高生存率,并且与增加的毒性相关,这突出表明在放疗完成后开始ICI治疗是首选策略。
微生物群与结直肠癌
微生物群作为CRC的重要外部驱动因素,通过多种方式促进肿瘤发生,包括免疫抑制、DNA损伤和致癌信号传导通路的激活。条件致病菌粪产碱杆菌在colitis期间易位至结肠组织,导致Peyer's patch IgA+ B细胞减少和免疫抑制。它还通过增加上皮紧密蛋白乙酰化,破坏其与β-连环蛋白的结合来促进癌变。由pks+ 大肠杆菌产生的基因毒素大肠杆菌素可直接导致宿主DNA双链断裂,其突变特征常见于邻近的正常结肠组织中。研究表明,富含可溶性纤维的低碳水化合物饮食可通过减少肠道炎症、增强PPAR-γ信号传导和降低硝酸盐水平来抑制pks+ E. coli的定植和大肠杆菌素介导的DNA损伤,从而预防结肠息肉形成。具核梭杆菌是另一种促进CRC癌变的机会性病原体。它通过黏附素(如FadA)形成生物膜,破坏肠道微生物群稳态并创造促炎性微环境。机制上,它抑制cGAS-IFNβ通路以损害CD8+ T细胞免疫反应,导致免疫治疗耐药。同时,感染重编程隐窝细胞,激活LY6A+ 复活干细胞并促进其向癌症干细胞转化,加速癌变。这种细菌通过多种协同途径促进癌症,包括毒力因子、免疫逃逸、干细胞重编程和代谢干扰。死亡梭杆菌产生5-氨基戊酸,这是一种通过抑制KDM6B、下调DKK2,从而激活Wnt/β-连环蛋白通路来促进肿瘤发展的代谢物。摩瑞氏梭杆菌通过其含有Cna B型结构域的细胞壁蛋白与宿主细胞上的整合素α2/β1结合,从而驱动CRC进展。这种结合激活了致癌的FAK-PI3K-AKT-mTOR-C-myc轴。通过阻断整合素α2/β1来靶向这种相互作用,为减轻其促癌作用提供了一种潜在策略。
目前针对类似微生物失调、致病菌或机会性病原体的治疗方法包括多种方法,如益生菌、益生元和抗生素来调节肠道微生物群。然而,针对特定致病菌的治疗包括靶向噬菌体疗法和特定疫苗。例如,针对瘤内具核梭杆菌,研究人员使用阳离子聚合物选择性包裹噬菌体头部,这可以在抑制细胞内致病菌的同时保持其完整性。此外,研究人员通过将具核梭杆菌膜与载药脂质体融合,制备了一种仿生纳米药物,可以选择性地杀死肿瘤内的细菌,而不伤害肠道菌群。
在详细描述了驱动结直肠癌发生的不同进化途径和微环境影响之后,显然需要系统的分子分类法将这种生物学复杂性转化为临床实践。CRC起源固有的异质性需要精确的分子分析来对患者进行分层、预测行为并选择靶向干预措施,从而弥合机制理解与治疗应用之间的差距。
结直肠癌的分子谱分析和分类
基于关键的分子变化,已经建立了多种CRC分子分类系统。最广泛采用的临床框架根据CIN状态、CIMP和MSI对CRC进行分类。这些分子类别涵盖了驱动CRC的主要遗传和表观遗传异常,对于指导精准治疗策略至关重要。
染色体不稳定性
CIN是CRC和腺瘤中最常见的基因组不稳定性形式,见于80%-85%的病例。其特征是广泛的染色体拷贝数变异和结构异常,导致肿瘤抑制基因(APC和TP53)的失活和癌基因(KRAS)的激活。KRAS编码能够促进细胞存活的GTP结合蛋白,而APC功能丧失破坏β-连环蛋白信号传导,最终导致腺瘤形成。CIN阳性的CRC通常为左侧和微卫星稳定(MSS),这些特征与不良预后相关。值得注意的是,F-box蛋白EMI1的表达降低已被证明可显著诱导结肠上皮细胞产生CIN。这伴随着DNA损伤增加和转化能力增强,突显了EMI1在CRC早期发展中的关键作用,并将其确立为一种有前景的生物标志物,可用于早期检测和靶向干预。
CpG岛甲基化表型
CIMP是一种表观遗传现象,其特征是多个CpG位点广泛高甲基化。CIMP被分层为高(CIMP-H;占CRC的15%-20%)和低(CIMP-L)亚型。CIMP-H与BRAF V600E突变、近端肿瘤位置和患者年龄较小密切相关,尽管分类标准仍不一致。CIMP-H通过沉默肿瘤抑制基因驱动CRC发病机制;例如,MLH1启动子甲基化导致错配修复缺陷(dMMR)和随后的MSI。临床上,CIMP-H CRC在早期阶段显示出改善的结局,但在转移期生存期差,并对5-氟尿嘧啶化疗耐药。最近一项关键性研究利用细胞和小鼠模型证明,IDH突变可直接诱导CRC中的CIMP表型。这些IDH突变与BRAF突变共分离,但与KRAS突变或MSI互斥。这一突破确定了常见恶性肿瘤中这种表观遗传机制的第一个遗传驱动因素,驳斥了与衰老相关的表观现象的学说,并立即指出IDH突变肿瘤是一个新的治疗靶点。
微卫星不稳定性
MSI存在于12-15%的CRC中,由dMMR导致,dMMR损害了短重复DNA序列的修复能力。大约90%的MSI是散发性的(由于MLH1甲基化),10%是遗传性的(林奇综合征)。MSI-H肿瘤通常位于近端,分化差,并且常携带BRAF突变。从预后角度看,MSI-H肿瘤在早期疾病中与改善的生存率相关,但在转移背景下化疗后结局较差。dMMR肿瘤表现出高肿瘤突变负荷(TMB),使其对ICIs高度敏感。虽然这些肿瘤在早期阶段显示出较好的预后,但它们在转移性疾病中表现出较差的结局和对5-FU的潜在耐药性。最近一项关键性研究揭示,丁酸梭菌通过调节GRP78-PI3K-AKT-NF-κB信号传导通路,进而减弱IL-6介导的免疫抑制微环境,从而提高抗PD-1疗法在MSI-H和MSS肿瘤中的疗效。值得注意的是,这些表型相互作用,因为CIMP可以通过甲基化沉默MMR基因来诱导MSI。此外,MSI状态不仅为免疫治疗提供信息,而且研究还发现Werner综合征RecQ解旋酶(WRN)可作为MSI亚型癌症的治疗靶点。抑制特定WRN残基的解旋酶结构域在MSI-H癌症中表现出合成致死性。此外,靶向WRN的药物正在积极开发中,抑制剂如HRO761和VVD-133,214目前正在进行临床研究。
此外,共识分子亚型(CMS)代表了最具影响力的CRC分子分类系统。基于基因转录组特征,它将CRC分为四个具有显著生物学和临床异质性的不同亚型。CMS1亚型(MSI-免疫)以MSI-H和高免疫浸润为特征,对免疫治疗敏感。CMS2亚型(经典型)以WNT/MYC通路激活为标志,并受益于基于奥沙利铂的化疗和抗EGFR治疗。CMS3亚型(代谢型)经常表现出KRAS突变和代谢重编程。CMS4亚型(间质型)表现出上皮-间充质转化和促纤维化的肿瘤微环境,与最差的预后相关。该分类也为指导治疗决策提供了重要框架。
这些分子谱为分类CRC提供了一个关键框架,然而,它们最终通过使核心细胞信号传导通路失调来发挥其生物学影响。因此,理解这些通路对于破译分子改变如何驱动肿瘤发生以及识别可操作的治療靶点至关重要。以下部分详细介绍了CRC中经常被破坏的关键信号级联反应。
结直肠癌肿瘤发生中的信号传导通路
分子亚型之间的差异源于各种信号传导通路的异常激活。CRC的发生和发展是由一个复杂的、相互交织的网络共同介导的,该网络涵盖血管内皮生长因子、Hippo、AMP激活的蛋白激酶、Wnt/β-连环蛋白、PI3K/AKT/mTOR、TGF-β、JAK/STAT、ErbB和Notch通路。这些级联反应之间复杂的串扰在不同程度上驱动恶性肿瘤(图2)。
图2. 参与细胞调控和增殖的信号通路示意图。该图阐释了多条调节基因达、蛋白质合成、细胞生长和增殖的信号通路。图示通路包括 Wnt/β-连环蛋白、受体酪氨酸激酶介导的(例如 ErbB 家族、PI3K-Akt-mTORC1)、IL-6-JAK-STAT、NOTCH、Hippo、能量应激相关的以及 VEGF-RAS-RAF-MEK-ERK 通路。这些通路汇聚于转录调控和细胞功能,如增殖、生长和代谢,并在信号转导的每个步骤中突出了关键分子。
Wnt/β-连环蛋白信号传导通路
Wnt/β-连环蛋白通路通过调节关键过程,包括细胞增殖、干性、凋亡、自噬、代谢、免疫、微环境调节、耐药性和转移,从根本上影响CRC的发生和进展。在CRC中,Wnt配体与LRP5/6和Frizzled受体复合物的结合招募Dishevelled,后者使通常靶向β-连环蛋白进行降解的破坏复合物失活。Wnt信号的激活导致β-连环蛋白积累、核输入,随后与TCF/LEF1因子结合,从而诱导致癌Wnt靶基因的转录。相反,当Wnt信号关闭时,破坏复合物保持功能,导致β-连环蛋白处于低稳态水平。
先前的研究已广泛总结了Wnt/β-连环蛋白通路在CRC中的作用。近年来,研发进展揭示了新的调控机制。例如,在CRC中过表达且对翻译起始至关重要的真核起始因子3亚基H,是由Wnt/β-连环蛋白信号转录诱导的。EIF3H作为HAX1的去泛素化酶,使其稳定免受βTrCP介导的降解。这种稳定增强了RAF1-MEK1-ERK1复合物,促进ERK1/2磷酸化和随后的致癌信号传导。除了EIF3H介导的调控分支,Wnt/β-连环蛋白靶基因SOX9在CRC中表现出背景特异性作用。APC和SOX9的同时失活驱使肿瘤向高度侵袭性表型发展,并伴有EMT和SOX2表达增加。SOX9的缺失加速转移并预测CRC预后不良。然而,在胃癌中,SOX9通过Wnt信号传导通路促进增殖和干性。
此外,Wnt/β-连环蛋白通路还可以通过明确的分子机制与VEGF和EGFR通路形成交叉调控网络。Wnt通路的激活可通过上调下游靶基因的转录表达间接促进VEGF的合成和分泌,从而调节肿瘤血管生成过程。Wnt通路可与EGFR通路下游的ERK信号形成协同效应,调节细胞周期蛋白D1等增殖相关基因的表达,协同促进肿瘤的恶性进展。
VEGF信号传导通路
VEGF通路对CRC的血管生成和转移至关重要。VEGF配体通过三个酪氨酸激酶受体以及共受体神经纤毛蛋白-1和神经纤毛蛋白-2进行信号传导。VEGFR2对血管生成至关重要,其通过二聚化和自身磷酸化激活,触发PLCγ-PKC-Ca²⁺(血管通透性)、PI3K-Akt(细胞存活)和Ras-Raf-MEK-ERK(增殖和迁移)通路。相反,VEGFC和VEGFD主要结合VEGFR3以诱导淋巴内皮细胞增殖。这些配体形成VEGFR2异源二聚体的能力允许血管和淋巴系统之间发生关键的串扰,从而促进肿瘤播散。
VEGF信号在各种癌症中的核心作用已被充分证实。在CRC中,肿瘤来源的VEGFA驱动异常血管生成,支持生长和转移。因此,抗VEGF/VEGFR药物是转移性CRC的标准治疗方法。除了其已确立的激酶活性外,VEGFR2还具有重要的非经典功能。它通过招募β-arrestin以激酶非依赖性方式发挥作用,调节细胞迁移和血管完整性,并与成纤维细胞生长因子受体和EGFR等受体形成跨家族复合物,增加通路多样性并导致耐药性。VEGFR2还充当白细胞跨内皮迁移的中介,与PECAM-1和VE-钙黏蛋白形成机械传导复合物,通过Y1175位点的磷酸化启动TEM,此过程不依赖于VEGF结合或其内在激酶活性。内皮特异性缺失VEGFR2可阻断中性粒细胞渗出,将炎症性外渗减少≥75%,并确定了一个新的抗炎靶点。这项研究确立了VEGFR2作为IBD背景下血管生成的核心驱动因素。此外,炎症触发的STAT1上调TGM2,后者直接结合并磷酸化VEGFR2,在IBD中激活促血管生成信号。靶向TGM2可有效缓解colitis。此外,β-羟基丁酸通过抑制缺氧诱导因子1-α(HIF-1α)/VEGFA通路,减少CAC模型中的肿瘤负荷和血管生成,提供新的预防和治疗策略。
值得注意的是,VEGF通路通过激活下游的Ras-Raf-MEK-ERK级联反应,与CRC中核心的促存活EGFR/RAS/RAF/MEK/ERK通路形成了一个关键的交叉点。信号网络的这种复杂性解释了抗血管生成治疗的多效性效应,并强调了肿瘤细胞内源性促增殖通路的重要性。
EGFR/RAS/RAF/MEK/ERK通路
EGFR/RAS/RAF/MEK/ERK信号级联反应是一条核心的致癌通路。它在控制细胞增殖、存活和分化中起着基础性作用。该通路的异常激活是CRC发病机制中的一个关键驱动因素。该通路起始于细胞外配体(例如EGF)与EGFR酪氨酸激酶的结合。由这一事件诱导的受体二聚化和自身磷酸化导致衔接蛋白如Grb2和鸟嘌呤核苷酸交换因子SOS的招募。SOS促进膜结合G蛋白RAS上的GDP与GTP交换,有效地将RAS从非活性状态转变为活性状态,这是一个关键的分子开关。活性RAS-GTP招募并激活RAF,启动核心磷酸化级联反应:RAF-MEK-ERK。激活后,ERK发生核转位。在细胞核内,它磷酸化转录因子如c-Fos和c-Myc,这些转录因子调节包括细胞周期蛋白D1在内的靶基因表达,以驱动细胞周期进程。KRAS的高突变频率导致组成性RAS激活,持续驱动下游RAF-MEK-ERK级联反应并促进细胞存活。同样,BRAF V600E突变导致持续的MEK/ERK磷酸化,绕过上游调控。这些频繁且特征明确的功能获得性突变突显了该通路的核心作用,并定义了CRC治疗中的关键治疗靶点。
EGFR构成ErbB家族中的四个受体酪氨酸激酶之一。该家族在CRC中经常过表达,并通过共享的结构架构影响众多促肿瘤过程。ErbB2在CRC中经常过表达,与不良预后和增加的肿瘤侵袭性相关,特别是在KRAS G13D突变背景下。新的调控机制表明,转录因子ELF3在HER2过表达下上调KRAS表达,促进西妥昔单抗耐药。此外,细菌口腔消化链球菌通过其FBA蛋白与CRC细胞上的整合素αβ/β4结合,启动MEK-ERK-p90信号级联反应,以增强增殖、减少凋亡并破坏肠道屏障完整性,从而促进HER2激活。ErbB3尽管具有最小的内在激酶活性,主要通过异源二聚化(例如与HER2)发挥作用,放大致癌信号。ErbB4被神经调节蛋白和β细胞素等配体激活,通过PI3K/Akt和Shc通路促进增殖和转移,同时抑制分化。该网络的关键性质通过以下观察得到证实:HER4的缺失或HER3的抑制会增加凋亡,可能通过损害HER3-HER4异源二聚体依赖的Akt信号传导。一致地,ErbB4敲除减少小鼠肠道肿瘤的形成,并且其抑制使KRAS突变的CRC细胞对凋亡敏感。
RAS-RAF-MEK-ERK通路是ErbB受体下游的核心致癌信号。EGFR可以激活RAS-RAF-MEK-ERK和PI3K-AKT-mTOR通路,但像HER2-HER3这样的复合物优先通过PI3K-AKT进行信号传导。这种差异性的通路激活为将ErbB抑制剂与下游通路抑制剂联合使用以规避耐药机制提供了理论基础。
PI3K/AKT/mTOR通路
PI3K/AKT/mTOR通路是CRC中常见的激活事件,与EGFR/RAS通路密切相关,是肿瘤代谢重编程、血管生成和转移的关键调节因子。在CRC中,当I类PI3K酶将PIP2转化为脂质第二信使PIP3时,该通路显著激活。这一事件招募并诱导Akt磷酸化,启动下游级联反应。新的调控机制揭示了多种调节因子,包括编码和非编码RNA以及外部因素。最近的研究表明,富含脯氨酸/精氨酸的末端亮氨酸丰富重复蛋白(PRELP)在CRC中发挥肿瘤抑制作用。其表达在CRC组织中显著下调。机制上,PRELP结合成纤维细胞生长因子1(FGF1),促进其降解。这随后使PI3K/AKT/mTOR通路失活,最终减轻肿瘤血管生成和转移。此外,外泌体长链非编码RNA MALAT1通过充当miR-26a/26b的microRNA海绵,驱动CRC进展。这种机制减轻了对岩藻糖基转移酶4的抑制,并导致PI3K-AKT-mTOR-C-myc信号轴的激活。此外,在CRC患者粪便中富集的厌氧菌摩瑞氏梭杆菌促进CRC肿瘤生长,其细胞壁与CRC细胞上的整合素α2/β1结合,从而激活PI3K/AKT/mTOR通路以刺激肿瘤生长。重要的是,阻断这种特定的整合素成功地消除了观察到的促肿瘤效应。此外,SLIT和NTRK样家族成员4的过表达显著激活PI3K/AKT通路,导致细胞外基质和细胞因子谱发生变化。SLITRK4促进TAMs的浸润和极化,促进CRC发生和随后的肝转移。同样,在约30%的CRC病例中过表达的LIN28B,也利用PI3K/AKT通路加剧向肝脏的转移播散。
PI3K/AKT/mTOR通路是来自不同上游通路信号的中心整合枢纽,包括EGFR的强效激活和Notch信号的转录增强。该级联反应是肿瘤存活和代谢重编程的关键介质。其广泛的通路内串扰决定了在临床研究中需要进行联合靶向抑制,以阻断补偿性信号传导并克服治疗耐药性。
JAK/STAT信号传导通路
JAK/STAT信号传导通路通过整合细胞增殖、代谢、炎症驱动的肿瘤发生和免疫逃避信号,在CRC中发挥关键作用。细胞因子和生长因子的结合诱导受体多聚化并激活相关的JAK激酶。然后JAKs经历自身磷酸化并磷酸化被招募的STAT转录因子。这种机制将信号直接从膜传导至细胞核,影响肿瘤进展和肿瘤微环境。此外,组织金属蛋白酶抑制剂-2在5-FU耐药的CRC细胞中显著上调。研究表明,TIMP-2通过激活JAK-STAT通路特异性促进对5-FU化疗的耐药性,建立了一种新的化疗耐药机制,对联合治疗具有潜在意义。
JAK/STAT通路与NF-κB和PI3K/AKT信号传导相交。促炎性细胞因子同时激活JAK/STAT和NF-κB。此外,PI3K/AKT通过磷酸化STAT3增强其转录活性,从而强化信号。这种协同作用驱动炎症驱动的肿瘤发生,并支持在CRC治疗中将JAK/STAT抑制剂与抗炎或PI3K/AKT靶向药物联合使用。
Notch信号传导通路
高度保守的Notch信号传导通路对于协调发育和稳态过程中的生理过程至关重要,其失调对于CRC中的细胞命运决定、干细胞维持和转移至关重要。该通路围绕Notch跨膜受体构建,该受体具有Notch胞外结构域、跨膜结构域和Notch胞内结构域(NICD)。激活通过一个独特的、多步骤的蛋白水解切割机制发生:配体(例如DLL1)与NECD结合诱导构象变化并释放胞外结构域。这一关键事件允许NICD从膜上切割下来。一旦被切割,NICD转位到细胞核,在那里它作为转录共激活因子发挥作用。在细胞核中,其RAM结构域介导与CSL转录因子的结合。这种相互作用促进了像MAML这样的共激活因子的招募,导致转录活性NICD-CSL-MAML三元复合物的组装。然后该复合物启动效应基因的转录,从而调节对CRC进展至关重要的核心细胞过程。
在对CRC中Notch信号进行全面解读之后,新的调控机制已经出现。内皮细胞中胶质瘤肿瘤抑制候选区域基因1的缺失导致Notch配体JAG1上调。JAG1随后与CRC细胞上的Notch受体结合,激活该通路并促进持续的肿瘤增殖和转移。此外,炎症微环境中的免疫细胞激活PI3K/AKT通路,该通路特异性作用于Notch4。这种级联反应通过新的Notch4-GATA4-IRG1轴促进肿瘤存活和增殖。锌指蛋白217的过表达也通过激活癌症干细胞标志物和刺激Notch通路,增强CRC干细胞的自我更新能力并促进肿瘤发生。与Wnt/β-连环蛋白通路的功能协同作用,其中Wnt增强Notch配体表达,Notch上调β-连环蛋白靶标,证明了联合靶向这些通路以消除癌症干细胞和预防转移的合理性。
Hippo信号传导通路
Hippo信号传导通路是感知细胞接触、机械力和极性的核心,是CRC进展和化疗耐药的关键调节因子。核心激酶级联涉及MST1/2激酶激活LATs1/2和MOB1,导致转录共激活因子YAP的抑制。此外,Hippo激活的MST1/2激酶磷酸化并抑制PI5P4Ks,这些酶通常抑制MOB1磷酸化和LATS激活。这种调控的缺失增强了YAP去磷酸化和核转位,从而促进CRC中的增殖。此外,具核梭杆菌通过调节Hippo通路介导CRC化疗耐药。这种微生物作用上调B细胞淋巴瘤2表达,并抑制化疗诱导的半胱天冬酶-3/GSDME介导的细胞焦亡,揭示了一种决定治疗结果的特定微生物-宿主信号相互作用。
CRC中对pan-TEAD抑制剂的耐药涉及AP-1的上调和YAP-TEAD致癌复合物的重建。虽然抑制剂有效阻断初始YAP-TEAD相互作用并降低fos样抗原1(FOSL1)活性,但耐药细胞通过恢复YAP/TEAD染色质结合和增强MAPK信号传导来克服这一点,其中FOSL1对于稳定的YAP-TEAD结合至关重要。这种揭示的耐药机制突显了CRC中Hippo和MAPK通路之间的关键串扰,为将TEAD靶向治疗与MAPK抑制剂联合使用提供了明确的理由。这种方法因Hippo通路与MAPK和PI3K/AKT级联的调控趋同性而得到进一步加强,其中MAPK通过磷酸化增强YAP核定位,而PI3K-AKT通过抑制LATs1/2促进YAP激活。因此,将Hippo靶向治疗与MAPK或PI3K抑制剂联合使用对于阻断补偿性存活机制至关重要。
AMP激活的蛋白激酶通路
AMPK通路代表了细胞能量稳态的核心调控系统,在代谢重编程和应激适应中发挥关键作用。AMPK通过感知AMP/ATP比率来协调生长和代谢过程。在CRC中,其激活受变构调节和磷酸化的协同调控。在能量应激下,AMP/ADP与γ亚基结合诱导变构变化,暴露活性位点,促进上游激酶对α亚基Thr172的磷酸化。相反,高ATP水平稳定了非活性构象,并且磷酸酶使Thr172去磷酸化,从而抑制活性。在代谢重编程的背景下,AMPK激活对脂质代谢发挥抑制作用。它磷酸化关键的脂肪生成酶,如ACC-1,从而抑制对脂质生物合成和癌细胞增殖至关重要的FASN、SREBP-1c和SCD-1的表达。此外,在CRC模型中,像白藜芦醇这样的化合物可以通过AMPK激活来减轻Warburg效应。参与代谢调节的关键葡萄糖转运蛋白GLUT3在CRC中高表达,其过表达与不良预后相关,进一步突出了其在代谢控制中的双重作用。在葡萄糖缺乏条件下,GLUT3通过增加葡萄糖摄取和促进核苷酸合成来促进CRC进展。AMPK/CREB1通路在应对低葡萄糖应激时显著上调GLUT3表达,关键性地调节这一过程。
信号传导通路作为一个复杂的交叉调控网络运行,共同驱动结直肠癌变。AMPK通路作为一个核心代谢调节因子,与致癌级联反应相互作用。该通路负向调节PI3K/AKT下游的mTOR信号传导,通过磷酸化调节Wnt/β-连环蛋白活性,并通过能量感应影响Hippo效应器。同样,EGFR/RAS/RAF/MEK/ERK轴在血管生成中与VEGF信号传导相交,而JAK/STAT和Notch通路促进炎症驱动的进展。这种广泛的通路串扰网络有助于解释单通路靶向治疗疗效有限的原因,并突显了需要能够同时抑制补偿性信号机制和克服耐药性的多靶点策略。
CRC靶向治疗的发展直接受到其复杂的分子发病机制的推动,该机制涉及信号传导通路之间的复杂相互作用。同时,破译这些通路内的关键分子节点为这种治疗策略提供了理论基础和可行的靶点。
结直肠癌的靶向治疗
基于CRC的分子发病机制,靶向治疗已成为精准治疗的关键方法,特异性阻断关键的致癌通路。这种方法通过选择性地破坏致癌信号和凋亡通路,从而最大限度地减少对正常组织的毒性,实现精确的肿瘤根除。基因测序的快速发展导致了许多关键靶点的识别,促使形成了一个包括小分子抑制剂和单克隆抗体的治疗武器库。这些关键靶点及其相应药物的机制、临床应用和未来轨迹将在以下部分进行系统探讨。
靶向Wnt/β-连环蛋白信号传导通路
Wnt/β-连环蛋白通路是大多数CRC中的关键致癌驱动因素,但由于β-连环蛋白缺乏可成药的结合口袋、其在肠道干细胞中的重要作用以及反馈耐药机制,它仍然是一个“不可成药”的靶点,尚无批准的直接抑制剂。
因此,治疗焦点已转向间接策略,这些策略可以按发展阶段分类。正在临床研究的药物:PORCN抑制剂通过阻断Wnt配体分泌,并已显示出早期的机制验证,但其发展受到骨疾病等剂量限制性毒性的阻碍。它们现在主要应用于生物标志物选择的患者。另一种方法是靶向β-连环蛋白转录复合物,使用如PRI-724或E7386等药物,旨在破坏致癌转录,目前正处于早期临床评估阶段。
临床前和药物再利用候选药物包括恢复β-连环蛋白降解的策略。Tankyrase抑制剂稳定AXIN,但会导致靶向毒性,而再利用药物pyrvinium作为CK1α激动剂以增强β-连环蛋白破坏。诸如PROTACs和靶向共激活因子BCL9的合成致死策略等新型模式也正在探索中。关键的转化障碍是靶向毒性、通路复杂性和缺乏生物标志物。克服这些障碍将需要创新的递送系统和合理的组合策略,以最终将Wnt抑制整合到CRC精准医学中。
靶向VEGF通路抑制剂
作为病理性血管生成的主要调节因子,VEGF信号传导是CRC增殖和转移的核心,验证了抗血管生成治疗作为核心治疗策略的合理性。
临床批准的药物:当前的VEGF靶向药物根据其作用机制进行分类。单克隆抗体,如贝伐珠单抗,在细胞外起作用,通过螯合VEGF配体,阻止其与VEGF受体结合并抑制下游信号传导。尽管这种方法带来了生存获益,但长期治疗与药物耐药性的产生相关,通常由替代性促血管生成因子的上调介导,并且临床上存在具有挑战性的系统性不良反应,包括高血压和蛋白尿。第二类是小分子抑制剂,如瑞戈非尼,是多激酶抑制剂,在细胞内起作用。从药理学角度看,这些药物受到选择性差的限制,这导致剂量限制性脱靶毒性,并限制了最大耐受剂量下的药物暴露,导致VEGF抑制不充分且短暂。总的来说,已建立的和较新的药物如呋喹替尼、安罗替尼和faricimab的局限性,包括获得性耐药和系统性毒性,突显了持续开发新型VEGF靶向疗法的迫切需求,以为CRC患者提供更安全、更持久的治疗结果。
正在临床研究的药物:Brivanib是一种口服酪氨酸激酶抑制剂,选择性靶向FGFR和VEGFR,从而阻断对肿瘤血管生成和增殖至关重要的信号传导通路。在临床前研究中,brivanib在肝细胞癌异种移植模型中显著抑制肿瘤生长,降低微血管密度并增加凋亡。然而,来自brivanib-西妥昔单抗治疗mCRC的临床观察表明,治疗诱导的高血压与患者反应或生存无关(P>0.05)。代谢组学分析确定了29种与血管舒缩失调相关的TI-HTN相关代谢物,与子痫前期的特征重叠;然而,缺乏生存获益表明TI-HTN不应作为治疗疗效的替代标志物,将其相关性局限于毒性机制的研究。Nintedanib单独或联合卡培他滨治疗难治性mCRC。
细胞培养和肿瘤异种移植模型的实验表明,呋喹替尼与阿霉素和奥沙利铂等化疗药物有效协同,肿瘤生长抑制率增加约30%。值得注意的是,在较低剂量下,呋喹替尼与抗PD-1抗体联合使用时,可增强抗肿瘤免疫反应。这种免疫调节作用的特点是血管生成减少、CD8+ T细胞迁移增加以及巨噬细胞和MDSCs等免疫抑制细胞比例降低,从而重塑肿瘤免疫微环境。针对这一特定分子通路的临床研究包括四项已完成的研究,尽管详细结果尚未公布。其他研究目前正在开发中。
靶向EGFR/RAS/RAF/MEK/ERK通路
除了抗血管生成,EGFR/RAS/RAF/MEK/ERK通路是一个主要靶点。其异常激活是CRC癌变和进展的核心。关键信号节点明确的突变状态为治疗选择提供了明确的分子基础,为这种疾病的靶向干预策略提供了清晰的路线图。
靶向EGFR家族
考虑到异常EGFR激活与肿瘤细胞增殖和生存的强关联,靶向EGFR通路是抑制CRC增殖和生存的关键方法。抗EGFR药物的疗效与下游组分的突变状态内在相关,证实了分子谱分析对于治疗选择至关重要。
临床批准的药物:主要获批的抗EGFR药物是单克隆抗体,西妥昔单抗和帕尼单抗,它们通过结合EGFR胞外域起作用。这种结合抑制配体诱导的二聚化和自身磷酸化,抑制下游RAS-RAF-MEK-ERK和PI3K-AKT-mTOR信号传导通路。它们的治疗功效严格依赖于下游效应物的突变状态。这些药物仅对RAS/BRAF野生型的mCRC患者有效。发生在40%-50%患者中的KRAS突变,组成性激活MAPK/PI3K轴,绕过上游EGFR阻断,代表了内在耐药的主要机制。原发性和获得性耐药的高发生率极大地限制了抗EGFR单药治疗的临床获益。因此,持续研究的重点是开发具有额外、次要靶点的药物,以规避这种耐药性并提高治疗效果。
正在临床研究的药物:为了克服单靶点抑制的局限性,正在研究新一代药物,分类如下:1)新型TKI:BAY 2927088是一种选择性化合物,是一种具有双重靶点机制的低分子量TKI,并抑制二氢乳清酸脱氢酶(DHODH)。它在非小细胞肺癌中显示出有前景的初步疗效,并正在CRC的临床研究中进行评估。2)双特异性抗体:这些药物旨在克服通路旁路引起的耐药性。SI-B001,一种EGFR×HER3双特异性抗体,同时抑制EGFR同源二聚体和EGFR/HER3异源二聚体,旨在抢先关闭下游RAS-RAF-MEK-ERK和PI3K-AKT-mTOR信号传导,以克服旁路介导的耐药性。另一个关键药物MCLA129是一种双特异性抗体,同时靶向EGFR和间充质-上皮转化因子受体。通过阻断这两条致癌通路,MCLA-129抑制肿瘤生长,并通过增强抗体依赖性细胞介导的细胞毒性作用来增强宿主免疫力,从而促进癌细胞破坏。3)生物类似药和结构优化的抗体:QL1203是一种重组全人源抗EGFR单克隆抗体注射液,作为帕尼单抗的生物类似药。在用于RAS野生型mCRC一线治疗的III期临床研究中,QL1203显示出与参比产品相当的疗效和安全性概况。考虑到这种临床等效性,预计QL1203将成为已建立的EGFR靶向单克隆抗体疗法的一种可及且成本效益高的替代方案,特别是在中国等地区。此外,结构优化的药物如人源化抗EGFR单克隆抗体正在开发中,旨在提高亲和力并降低与西妥昔单抗相比的免疫原性。目前,该药正在进行mCRC的II期临床研究。
HER2是ErbB受体家族的致癌成员。使用特异性抗体或抗体药物偶联物靶向HER2,对于大约3%的具有HER2过表达或扩增的CRC患者(一个通常对常规化疗耐药的亚组)来说,代表了一种高效的治疗策略。基于HERACLES、MyPathway、MOUNTAINEER和DESTINY-CRC01研究的有力数据,HER2已被正式验证为CRC中治疗上可操作的驱动生物标志物。
临床批准的药物:单克隆抗体曲妥珠单抗和帕妥珠单抗的联合是一种已确定的方案。抗HER2药物如曲妥珠单抗选择性地与HER2结合,破坏下游信号传导以抑制肿瘤生长和增殖。除了单克隆抗体,新型模式也在研究中。ADC是由通过合成连接子与强效细胞毒性药物偶联的单克隆抗体组成的先进复杂生物药物。它们有助于将化疗精确递送至HER2表达的肿瘤,最大限度地提高治疗效果且不影响健康组织。抗HER2单克隆抗体曲妥珠单抗和帕妥珠单抗与ADC德曲妥珠单抗的联合提供了一种有前景的新治疗选择。T-DXd在DESTINY-CRC01研究中显示出卓越的疗效,实现了45%的ORR和8.2个月的中位PFS。值得注意的是,尽管存在通常对上游靶点产生耐药的RAS/RAF突变,T-DXd仍能维持疗效。这一发现提示了HER2阳性CRC治疗模式的潜在转变。
正在临床研究的药物:几种下一代HER2靶向ADC目前正在开发中。这包括新型ADC,如A166,I期研究已证明其具有可控的安全性概况、良好的药代动力学和有前景的初步抗肿瘤活性。新型抗体药物偶联物ARX788在预处理过的HER2阳性晚期乳腺癌患者中显示出优于先前治疗的效果,显著提高了中位无进展生存期和客观缓解率。目前正在多项针对HER2表达的晚期实体瘤(包括CRC)的临床研究中对其进行评估。I期研究已完成。新兴的抗HER2治疗领域包括ACE1702,一种现成的抗体偶联NK细胞疗法。这种细胞药物目前正在一项针对包括CRC在内的HER2表达晚期或转移性实体瘤的I期临床研究剂量递增阶段进行评估。此外,小分子抑制剂sevabertinib,一种口服、可逆、突变选择性的双靶点EGFR/HER2 TKI,提供了一种有效靶向特定突变的策略。该药进展迅速,目前正在进行针对CRC和其他HER2突变实体瘤的全球II期临床研究。AZD8931是一种靶向EGFR、HER2和HER3的小分子TKI,在与FOLFIRI联合用于mCRC的I/II期研究中显示出良好的耐受性和初步疗效,此后已推进至II期。
一项评估双特异性抗体zanidatamab联合化疗在HER2表达的晚期胃食管腺癌、胆道癌和CRC中的II期研究是最近完成的三项研究之一。该研究揭示了ZW25在HER2阳性晚期胃食管腺癌中的强大一线疗效,确认的ORR为76.2%,中位OS为36.5个月,PFS为12.5个月。该方案保持了可控的安全性概况,腹泻是主要不良事件,通过预防性用药可显著减轻。这些有力的结果表明,ZW25联合化疗可能成为HER2阳性晚期胃食管腺癌新的标准一线治疗方案。其余两项研究的数据有待公布。
靶向KRAS和BRAF
KRAS和BRAF突变是关键致癌驱动因素,导致通路组成性激活和细胞自主生长。靶向关键通路节点对于携带这些特定突变的肿瘤来说是一种高度合理的方法。为了更清晰地展示药物研发格局,下文根据发展阶段讨论靶向药物。
临床批准的疗法:由于固有的耐药机制,靶向BRAF突变的CRC具有挑战性。BRAF抑制剂单药治疗显示出较差的反应率,因为快速且补偿性的EGFR反馈回路迅速重新激活MAPK通路。为克服反馈和化疗耐药的需求,导致采用恩考芬尼加西妥昔单抗联合mFOLFOX6作为BRAF V600E突变mCRC的一线标准治疗方案,而二线治疗则使用双联疗法。
正在临床研究的药物:几种靶向突变型KRAS和BRAF的药物正在进行积极的临床研究。KRAS靶向药物:索托拉西布,首个被美国FDA批准用于转移性NSCLC的KRAS G12C抑制剂,在CRC中仅显示出9.7%的适度ORR。目前,有多种靶向KRAS癌基因的药物正在进行临床研究,包括靶向KRAS G12C和G12D等突变的抑制剂。KRAS G12C选择性共价抑制剂adagrasib是一个有前景的替代品,于2018年通过针对晚期实体瘤的KRYSTAL-1研究开始临床评估。Adagrasib已推进至针对KRAS G12C突变CRC患者的III期临床研究。多种靶向KRAS癌基因的新型药物目前正在进行临床研究。BRAF靶向药物:BRAF突变的CRC对伊立替康等标准化疗药物表现出耐药性。需要新型BRAF靶向药物和优化的联合方案来改善患者结局。下一代研发管线包括几种候选药物,如napafenib,一种双靶点BRAF/CRAF激酶抑制剂,在研究中显示出有前景的结果后,目前正在CRC中进行评估。
KRAS或BRAF突变的NSCLC和NRAS突变的黑色素瘤。一种极具前景的新药是BGB-3245,一种选择性口服小分子BRAF抑制剂。BGB-3245表现出广谱活性,靶向BRAF V600和非V600突变以及RAF融合蛋白。临床前研究表明其能够抑制单体和二聚体BRAF形式,从而减少早期抑制剂常见的二聚体驱动的耐药性。BGB-3245的临床开发始于2020年2月启动的I期研究。截至2022年的入组数据证实,42名患者接受了治疗,结果显示该药物具有可控的安全性概况。此外,一项I/IIa期联合研究目前正在进行中,研究BGB-3245与mirdametinib联合的治疗潜力。这些举措是更广泛的、活跃的临床领域的一部分,该领域还有几种其他的BRAF抑制剂。
两项已完成或正在进行的早期临床研究展示了实体瘤治疗中的新方法。主要研究ELI-002 2P是一项首次人体I期研究,评估一种新型免疫疗法在胰腺癌或CRC微小残留病背景下的安全性、耐受性和免疫原性。这项研究的重点是剂量递增,以确定合适的II期剂量。相反,研究LGX818报告了在mCRC患者亚组中的初步疗效信号,在300mg和400mg剂量组中分别报告了4.5个月和4.0个月的OS。这些研究共同描绘了当前临床发展的轨迹,涵盖剂量探索、初步疗效信号读取以及针对实体瘤的靶向和免疫疗法待公布的数据。
靶向MEK和ERK
作为RAS/RAF/MEK/ERK信号传导通路中的关键激酶,MEK是一个主要的治疗靶点。MEK抑制剂如曲美替尼和考比替尼通常用于联合方案以实现增强的抗肿瘤效果。临床批准的疗法:曲美替尼,一种FDA批准的高选择性MEK1/2抑制剂,是达拉非尼治疗BRAF V600E突变转移性黑色素瘤的既定合作伙伴。在CRC中,抑制MEK有效阻断RAS-RAF-MEK-ERK级联反应,在抑制肿瘤细胞增殖和存活中起关键作用。考比替尼和比尼替尼也是FDA批准的口服、非ATP竞争性MEK1/2抑制剂,同样可破坏CRC中的RAS-RAF-MEK-ERK通路。这些抑制剂已在多种癌症中显示出显著的临床疗效,特别是当与BRAF抑制剂策略性联合以克服反馈激活时。正在临床研究的药物:在临床研发管线中推进,下一代药物avutometinib正在一项III期研究中与defactinib联合用于复发性低级别浆液性卵巢癌的研究。此外,一项独立的II期临床研究正在评估avutometinib在CRC中的疗效。
ERK是RAS-RAF-MEK-ERK信号传导通路中的关键下游因子,其过度激活与CRC增殖、存活和耐药相关。ERK激活驱动肿瘤发生,其抑制有效抑制肿瘤细胞活力。正在临床研究的药物:Ulicetinib是一种口服、可逆、ATP竞争性ERK1/2抑制剂,通过抑制催化活性来阻断下游MAPK信号传导。I期研究结果表明,ERK1/2抑制剂ulixertinib有明确的推荐剂量为600mg每日两次。该药物表现出良好的安全性概况,并在携带NRAS或BRAF突变的晚期实体瘤患者中显示出初步的抗肿瘤活性。此外,IPN01194是一种同类首创的口服小分子ERK1/ERK2抑制剂,目前正在进行I/IIa期临床开发,该药物靶向MAPK通路的下游节点以阻断异常增殖信号。MK-8353是另一种口服生物可利用的ATP竞争性小分子ERK1/2抑制剂。其Ib期研究评估了MK-8353与MEK抑制剂selumetinib在晚期实体瘤中的联合应用,重点关注安全性、耐受性和确定推荐的II期剂量。
临床前/实验性:LY3214996是一种口服可用的ERK1/2抑制剂,作为ATP竞争性拮抗剂,从而阻断ERK1和ERK2的激酶活性,抑制下游MAPK级联反应。通过抑制底物的磷酸化,LY3214996促进G1期阻滞和凋亡。值得注意的是,该化合物能够克服通常由上游BRAF或MEK抑制剂引起的ERK再激活介导的耐药性。CheckMate 142研究证实了纳武利尤单抗联合伊匹木单抗在复发/转移性MSI-H CRC中的强大疗效,与单药治疗相比实现了高ORR。这种疗效在非MSI-H/MS患者或某些联合方案中未得到复制,表明MSI状态与治疗反应之间存在强烈关联。
靶向PI3K/AKT/mTOR通路和SHP2
PI3K/AKT/mTOR信号传导通路对CRC的发生、进展和转移至关重要,其失调与肿瘤侵袭性和耐药性密切相关。由于这一核心致癌作用,PI3K靶向疗法备受期待。以下按发展阶段介绍PI3K/AKT/mTOR靶向药物。
临床批准的药物:抑制剂如依维莫司成功抑制该通路并抑制肿瘤细胞增殖。此类药物用于治疗其他癌症已获FDA批准,包括idelalisib、copanlisib、duvelisib和alpelisib。正在临床研究的药物:在CRC中实现疗效的途径很复杂。早期的广谱抑制剂,如pan-PI3K抑制剂PX-866,在联合治疗中未能改善患者结局,反而增加了治疗相关毒性。这一结果反映了特异性有限的问题,因为正常细胞中的PI3K抑制会引起剂量限制性不良反应。新型小分子有机杂环药物BBO-10203特异性阻断致癌RAS-PI3Kα相互作用,从而限制肿瘤增殖,而不会引起高血糖症等系统性影响。临床前异种移植模型已证明其疗效和良好的低毒性特征,促使其目前进入临床研究。PI3K通路仍然是肿瘤学的焦点,其他抑制剂,如用于B细胞恶性肿瘤的parsaclisib,正处于积极的临床评估中。
此外,肿瘤细胞内的补偿性信号机制会进一步削弱治疗效果。因此,开发高选择性的下一代PI3K靶向药物以及战略性探索有效的联合方案是推进CRC治疗的关键方向。
非受体蛋白酪氨酸磷酸酶Src同源2结构域含磷酸酶2是调节包括RAS-MAPK和PI3K-AKT通路在内的多个下游级联反应的关键信号节点。其作为多种致癌信号汇聚点的功能使SHP-2成为同时抑制多个失调增殖通路的高潜力治疗靶点。SHP-2功能获得性改变驱动肿瘤发生,该磷酸酶的药理抑制减弱肿瘤细胞增殖和存活。尽管目前尚无获批的SHP-2抑制剂,但有几个候选药物正在临床开发中。正在临床研究的药物:TNO-155是最先进的,目前在II期临床研究中处于领先地位。
基于现有公开数据,针对晚期实体瘤的新型药物的临床格局显示出少数明确的阳性疗效信号,大多数结果仍悬而未决。一个显著的例外是DKN-01在二线MSS mCRC中的II期研究。尽管总体人群未达到统计学显著性,但DKK1高表达的患者获得了显著的临床获益,表现为改善的mPFS、mOS和ORR。这些发现突显了生物标志物驱动的患者选择的重要性。相比之下,其他正在进行的研究的临床相关性,包括针对BKM120、MEN1611、INCB050465、Nab-Rapamycin、TNO155和JAB-3068的研究,尚无法评估,因为关键的疗效和安全性数据尚未公布。
PARP抑制剂
靶向DNA修复机制的缺陷是CRC治疗中传统细胞增殖抑制剂的一种战略性补充。聚ADP-核糖聚合酶作为DNA修复的关键协调者,是CRC的一个有前景的靶点,特别是在具有潜在DNA修复缺陷的患者中。以下按发展阶段对PARP抑制剂进行系统性概述。
正在临床研究的药物:PARP抑制剂利用合成致死的原理,证明对携带BRCA1/BRCA2突变的肿瘤最有效。已获批药物如奥拉帕利和鲁卡帕利目前正在扩展其治疗应用。尽管如此,现有抑制剂的效用受到两个关键问题的限制:(1)获得性耐药的常见发展,这损害了长期疗效;(2)显著的剂量限制性不良反应,如骨髓毒性、疲劳和胃肠道毒性。下一代PARP靶向药物的设计应旨在克服耐药性并减轻这些毒性。几种PARP抑制剂正在CRC的临床研究中进行评估。例如,机制研究表明,在CRC干细胞中,PARP抑制剂veliparib通过使MSH6失调并随后抑制错配修复通路,增强5-FU的细胞毒性作用。
两项已完成的临床研究提供了关于奥拉帕利在晚期CRC中差异性疗效的见解。NCT04456699研究纳入了未经特定生物标志物选择的广泛患者队列,显示奥拉帕利单药治疗以及奥拉帕利联合贝伐珠单抗相比标准化疗有生存获益。相比之下,靶向MGMT高甲基化的NCT04166435研究报告,奥拉帕利联合替莫唑胺的肿瘤ORR为0%,但实现了55.6%的DCR,显示出显著的肿瘤稳定作用。这些发现表明,奥拉帕利的临床获益主要取决于治疗方案和患者选择:它在未经选择的人群中带来适度的生存获益,但在明确的分子亚组中主要实现疾病稳定。
多靶点联合治疗
单药治疗常诱发耐药性,通常通过激活替代或平行信号传导通路,使癌细胞能够逃避单药效应。对于晚期或mCRC患者,单药治疗疗效不足以实现长期疾病控制。我们还对现有CRC临床药物进行了比较分析,以阐明它们在临床实践中的各自优势、局限性和适用场景。这种系统性比较涵盖了主要的治疗途径,包括靶向EGFR和VEGF通路的药物、针对BRAF V600E和KRAS G12C特定突变的药物、HER2扩增、NTRK融合、用于dMMR/MSI-H肿瘤的ICIs,以及更广泛的多激酶抑制剂。因此,联合使用靶向不同通路或同一通路多个节点的药物,对于获得协同抗肿瘤效应、抑制耐药性并最终改善临床结局至关重要。
联合治疗策略的迅速采用为晚期癌症患者带来了新的希望。一项评估garsorasib联合西妥昔单抗治疗KRAS G12C突变晚期CRC的研究表明,ORR和PFS显著改善,且安全性良好,为III期研究铺平了道路。在免疫治疗方面,与单药治疗相比,纳武利尤单抗联合伊匹木单抗显著延长了MSI-H或MMR缺陷患者的PFS,无论既往治疗如何,考虑到其可控的安全性,表明它可能成为一种新的标准治疗。一项研究调查了在不可切除的RAS/BRAF野生型mCRC中,诱导治疗后采用FOLFIRI联合帕尼单抗间歇给药策略,结果显示间歇组治疗中12个月PFS率显著高于连续治疗组。间歇方法将中位PFS延长至17.5个月,并降低了严重皮肤不良事件的发生率,显示出改善生活质量的希望。另外,对于BRAF V600E突变mCRC,恩考芬尼联合西妥昔单抗加mFOLFOX6在既往未经治疗的患者中显著改善了ORR和中位缓解持续时间,其安全性特征与单药一致,确立其为一个极具前景的治疗选择。多模式策略的整合,如靶向治疗、免疫治疗和微环境/肠道菌群调节,显著提高了CRC的治疗效果和预后。实现最佳结果需要精准联合治疗,这依赖于预测患者反应和改进方案的先进技术。成功的治疗取决于识别合适的患者亚群,正如双靶向HER2阻断在HER2扩增的mCRC中的疗效所示。患者来源的类器官正逐渐成为必不可少的患者特异性离体模型,能够预测个体患者反应并为个性化联合策略提供信息。
随着研究的不断深入,越来越多的药物和治疗方法被开发出来。然而,由于人体和肿瘤本身固有的复杂性以及它们之间显著的多样性和异质性,患者对同一种药物的反应差异很大,这继续给CRC的治疗带来一系列严峻的挑战。
当前挑战与未来方向
尽管CRC的分子研究推动了靶向治疗的显著进展,但主要的临床挑战依然存在。这些挑战包括获得性治疗耐药性的产生和显著的患者反应差异,这使得识别合适的候选者变得复杂。核心问题是需要更特异的预测性生物标志物来确保疗效。此外,这些特殊药物的高昂经济负担以及联合方案的复杂性限制了患者的可及性和依从性,这强调了持续努力优化靶向策略和改善患者结局的必要性。
肿瘤异质性与克隆进化
肿瘤异质性是靶向和联合治疗疗效的主要障碍,导致CRC中的耐药性和治疗失败。这种高度异质性和复杂的疾病在肿瘤类型、分期、位置以及核心分子水平上表现出变异性。异质性可以从四个维度进行系统性定义。1)遗传异质性:在克隆扩增过程中,CRC细胞积累了大量的体细胞突变,导致肿瘤内部和肿瘤之间出现不同的遗传景观。Colantunoi的研究团队揭示了CRC体细胞突变负荷比正常组织高出数倍。这种异质性不仅在不同患者之间观察到,而且在单个肿瘤的不同区域内也存在,并且在不同阶段进化,对统一的靶向治疗构成了根本性挑战。2)表观遗传异质性:Bao等人的一个关键发现是,与邻近的正常上皮相比,CRC肿瘤中观察到的全基因组DNA甲基化普遍较低。值得注意的是,同一肿瘤组织内的不同亚克隆也表现出差异甲基化水平。此外,蛋白质乙酰化等翻译后修饰在CRC转移中起主要作用,正如原发灶和肝转移灶之间乙酰化蛋白表达水平的显著差异所证明的那样。3)转录组异质性:使用单细胞RNA测序的高分辨率分析系统地描绘了CRC进展过程中多达48种不同细胞亚型的动态变化和分子谱。这些亚型表现出不同的基因表达模式,突显了显著的组间差异,这些差异与基本的肿瘤生物学行为密切相关,包括细胞增殖、侵袭和内在的治疗敏感性。4)功能/表型异质性:基本生物学特征的差异,包括增殖能力、特定代谢途径的利用和整体转移潜力,共同决定了肿瘤进展的轨迹和化疗的最终结果。
CRC异质性的复杂性质可分类为瘤内异质性和瘤间异质性。瘤内异质性指单个肿瘤内癌细胞间的差异,包括空间异质性和克隆异质性。相比之下,瘤间异质性突显了不同患者之间相同肿瘤类型的显著差异,例如不同的驱动基因突变、生长动力学和药物敏感性,强调了精准医学的挑战和重要性。这种患者内部和患者之间的多层次异质性,要求个性化和动态的治疗方案。考虑到CRC复杂、进化的性质,单靶点疗法频繁失败。因此,未来的策略必须纳入多靶点治疗以及持续的遗传监测,以实现持久的临床反应。
耐药机制
耐药性是CRC治疗失败的核心驱动因素,分为原发性和获得性两类。1)原发性耐药——缺乏初始反应性,机制复杂,通常由基因突变驱动。例如,RAS突变通过异常且持续地促进增殖和存活信号,从而克服药物对肿瘤生长的抑制作用,赋予对西妥昔单抗的耐药性。定量地看,RAS突变存在于大约50%的转移性CRC病例中,是对EGFR抑制剂产生原发性耐药的主要机制,超过90%携带这些突变的患者无法从此类治疗中获得临床益处。像存在于约3%的mCRC中的KRAS G12C突变这样的特定亚型,赋予不同的耐药谱。表型变化,如获得干细胞样特性,进一步促成了这种内在的药物耐受性。2)获得性耐药发生在肿瘤经过一段有效治疗后逐渐变得耐药时。肿瘤的遗传和表型异质性共同促进获得性耐药的发展。肿瘤细胞在遗传上进化,其中一些携带预先存在的耐药突变,赋予即时的适应性优势,并允许在肿瘤群体内迅速占据主导地位。获得性耐药表现为在经过一段有效治疗后肿瘤复发,由遗传和表型进化驱动。纵向基因组研究揭示,在最初对抗EGFR治疗有反应的患者中,获得性耐药是由大约30%至70%的病例中可检测到的RAS突变或EGFR胞外域突变的出现驱动的。治疗压力通过两条路径选择耐药克隆细胞:具有预先存在耐药突变的克隆的迅速主导,或者在其不存在时,依赖动态的表型适应。这些适应,包括转录启动和表观遗传可塑性,在药物暴露时启动,导致前药物耐受持久性细胞的形成。这些细胞最终从非增殖状态转变为增殖的DTP细胞,确保持续治疗下肿瘤细胞的存活和进展。定量模型表明,在治疗开始时,DTP群体可能占肿瘤细胞群体的一小部分但显著的比例,作为最终复发的储存库。
耐药性是癌症治疗中的一个关键的、多机制的挑战,涉及内源性和外源性因素。对抗EGFR单克隆抗体的获得性耐药性的发展通常涉及下游信号组分中继发性突变的获得,包括KRAS、NRAS和BRAF,这些突变阻止药物有效结合其靶点并维持组成性信号传导。类似地,在BRAF V600E突变肿瘤中长期使用BRAF抑制剂可导致通过替代信号传导通路的补偿性激活产生耐药性。对抗HER2疗法的耐药性也与平行信号分支的异常激活有关,例如由HER2扩增导致的PI3K-AKT激活,绕过预期的抑制信号。自噬在肿瘤发生中起着背景依赖性作用,能够作为肿瘤抑制因子或肿瘤启动子。这种功能转换受关键信号传导通路(如PI3K-AKT-mTOR轴和AMPK)通过遗传和环境因素调节。机制上,TIPE3已被证明通过上调USP19表达,影响Beclin1蛋白水平,从而增强自噬,并因此诱导对靶向药物的耐药性。
来自TME的外源性因素在耐药中也扮演着重要角色。乳酸的积累创造了一个酸性、营养和氧气缺乏的代谢环境,促进肿瘤进展、转移,并触发特定的耐药机制。例如,组蛋白H3K18la是一种表观遗传修饰,激活rubicon样自噬增强子以增强自噬,导致对贝伐珠单抗的耐药性。此外,胆固醇代谢物27-羟基胆固醇通过多种途径介导耐药性,包括促进药物外排、细胞增殖、抑制凋亡、诱导EMT和代谢重编程。因此,CRC耐药性是综合因素和途径的复杂结果。有效的逆转耐药策略必须全面考虑这些TME驱动和代谢机制,以实现精准和疗效。
肿瘤微环境与免疫逃逸
TME由微生物因素和宿主免疫相互作用塑造,通过免疫逃逸关键地驱动癌变。这个过程产生基因组不稳定性,促进异常信号传导,并通过招募抑制性细胞培育一个免疫抑制生态系统。肿瘤细胞通过几个关键机制逃避免疫监视:(1)下调抗原呈递以防止T细胞识别;(2)通过分泌因子和招募免疫抑制细胞(如Tregs)积极塑造TME;(3)上调免疫检查点分子,导致T细胞耗竭。总的来说,这些策略削弱了抗肿瘤免疫反应并促进了肿瘤生长。这些机制见解成功地支持了免疫疗法(如ICIs)的发展,这些疗法通过阻断抑制性信号来逆转免疫逃逸,开辟了新的治疗途径。
ICIs正在通过其阻断免疫逃逸途径的机制彻底改变CRC治疗。这类药物包括靶向CTLA-4、PD-1和PD-L1的药物。关键的KEYNOTE-177和CheckMate-142研究已确立ICIs作为MSI-H/dMMR状态转移性CRC的基石疗法。具体而言,III期KEYNOTE-177研究显示,一线帕博利珠单抗显著改善了PFS,同时具有比标准化疗更有利的安全性特征。II期CheckMate-142研究证实,纳武利尤单抗联合伊匹木单抗在既往治疗过的患者中实现了持久且高的ORR,为后续治疗线提供了一种有效、无化疗的方法。
尽管在MSI-H/dMMR肿瘤的免疫治疗方面取得了突破,但对于绝大多数(约95%)微卫星稳定/错配修复功能完好的CRC患者,仍然存在显著的治疗挑战。这些肿瘤通常具有非炎症性或“免疫排除”的TME,其特征在于低TMB、T细胞浸润不足以及免疫抑制细胞的积极作用。临床数据,包括CheckMate-142研究,已证实单药治疗或标准的双免疫检查点抑制剂联合治疗均不能在这一MSS人群中诱导有意义的反应。这些肿瘤中的TME通常由免疫抑制性相互作用主导,例如涉及T-bet+ Treg细胞的那些,它们通过高CD39表达抑制CD8+ T细胞功能。靶向这些关键的调节细胞代表了一种有前景的策略,以增强抗肿瘤免疫并改善免疫疗法的疗效。
为了克服MSS CRC对免疫治疗的耐药性,正在开发合理的联合策略,旨在将“冷”的非炎性TME转化为免疫浸润的“热”微环境。关键方法涉及将ICIs与积极调节免疫抑制性景观的药物联合使用。1)多激酶抑制剂:像瑞戈非尼这样的药物通过破坏促肿瘤因子(如TAMs)和血管生成途径显示出临床前疗效。这种调节为与ICIs联合以增强抗肿瘤免疫提供了坚实的理论基础。2)新型免疫调节靶点:在MSS CRC中高表达的肽基脯氨酰异构酶Pin1,通过NF-κB-CCL3-CCR5轴驱动免疫抑制。临床前研究表明,抑制Pin1可增强抗PD-1疗效,代表了一种有前景的新治疗靶点。3)研究性策略还包括将ICI与常规治疗或新型靶向药物联合使用,以实现协同免疫启动和检查点阻断增敏。一项研究开发了一种双特异性抗体ATAPL1,可同时靶向PD-L1和TNFR2。它在CRC小鼠模型中表现出优异的肿瘤靶向积累和抗肿瘤功效,且无明显毒性。其作用机制涉及重塑肿瘤免疫微环境,减少免疫抑制细胞,激活CD8+ T细胞和巨噬细胞,并启动长期免疫监视,为CRC治疗提供了一种有前景的新策略。肿瘤疫苗开发的研究也在取得进展,例如靶向肿瘤相关抗原或共享的新抗原;根据患者肿瘤突变谱设计mRNA或肽疫苗;或通过瘤内注射免疫刺激剂诱导系统性抗肿瘤免疫。CRC疫苗仍面临肿瘤异质性、抗原逃逸和免疫抑制微环境等挑战,但将其与ICIs联合使用的策略显示出潜力。另一项研究开发了一种携带趋化因子CCL19的重组溶瘤流感病毒rPR8-CCL19用于治疗CRC。该病毒选择性感染并杀死CRC细胞,在肿瘤微环境中表达CCL19,招募并激活树突状细胞和T细胞,将“冷”肿瘤转化为“热”肿瘤,有效抑制肿瘤生长和转移,并诱导系统性肿瘤定向免疫和持久免疫记忆,具有良好的安全性。
识别和验证新型生物标志物
当前的mCRC管理依赖于已建立的生物标志物来指导抗EGFR药物、BRAF/EGFR联合疗法和ICIs的使用。然而,这种静态方法无法捕捉关键的时空异质性和克隆进化,这些是驱动获得性耐药的因素。此外,它忽视了疾病的关键维度,包括转录组学、蛋白质组学和TME,从而限制了疗效。这些局限性突显了对动态、多维生物标志物的迫切需求,以优化治疗选择并推进CRC精准肿瘤学。
先进技术的整合正在推动一个动态CRC生物标志物发现的新时代。1)液体活检:循环肿瘤DNA允许进行非侵入性的实时监测,其最具影响力的应用是检测术后微小残留病。DYNAMIC研究表明,ctDNA指导的管理可以安全地减少II期结肠癌的辅助化疗负担。ctDNA对于早期检测获得性耐药突变也至关重要。2)多组学和人工智能:多种多组学方法,如转录组学CMS分类,可以识别生物学相关的亚组。此外,蛋白质基因组学也可以揭示基因组学遗漏的失调机制。同时,应用于数字病理学的AI提供了一种成本效益高的工具来预测分子特征。3)肿瘤微生物组:瘤内微生物已作为与不良生存和化疗耐药相关的预后因素出现。将这些新型生物标志物转化为常规实践面临着多重挑战。肿瘤异质性限制了单个样本的代表性,并且需要技术标准化以确保检测的可重复性。这些新型生物标志物的临床效用必须在临床采用前通过前瞻性、随机研究来确定。
CRC生物标志物开发和治疗的未来极具前景。关键方向涉及积极靶向历史上具有挑战性的驱动因素。继KRAS G12C抑制剂成功之后,研究重点集中在其他突变体上,例如进入临床研究的MRTX1133。此外,像PROTAC这样的技术提供了一种通用的策略来降解而非抑制有问题的癌蛋白,可能靶向以前“不可成药”的驱动因素,如转录因子。此外,对于耐药性MSS/pMMR CRC的大多数,基于新抗原的疫苗和工程细胞疗法正在开发中,以诱导从头免疫反应,初步研究显示出强大的免疫原性。
联合疗法的合理设计
克服CRC中靶向单药治疗的局限性需要对肿瘤生物学和适应性耐药有详细的机制理解。固有的肿瘤异质性和高治疗失败率需要转向合理的联合设计,旨在创造协同和持久的结局。这种联合采用垂直和水平抑制的逻辑来阻断肿瘤逃逸机制。垂直抑制涉及对单个中心通路的平行阻断。BRAF V600E突变mCRC的治疗是一个范例:单药耐药性由快速EGFR反馈激活驱动。合理设计的三联疗法实现了强大的垂直MAPK通路阻断,并具有显著的临床益处。此外,水平抑制将靶向治疗与免疫疗法等不同机制相结合。多激酶抑制剂瑞戈非尼通过调节免疫抑制性TME来体现这一点。其与纳武利尤单抗联合在通常对免疫治疗耐药的MSS mCRC队列中显示出初步活性。AI驱动的大规模药物筛选平台正在显著加速新型协同药物对和预测性生物标志物的发现。
显著的挑战限制了当前联合策略的影响,特别是管理累积毒性和防止适应性耐药。未来的成功取决于严格的、动态的耐药性监测以及高效、创新方法的临床采用。适应性临床研究设计对于在标准化框架内同时评估多种联合方案至关重要。最终,将深入的分子见解与先进的功能和计算技术相结合,是为CRC开发真正个性化、合理设计的联合方案的必经之路。
结论与展望
本综述系统总结了CRC分子发病机制、靶向治疗和免疫治疗的进展,分析了当前管理中的核心局限性。尽管其发病率和死亡率很高,但靶向疗法的发展已经彻底改变了CRC的治疗。我们确定了关键信号传导通路,包括Wnt/β-连环蛋白、PI3K/AKT/mTOR、EGFR、JAK/STAT、VEGF和AMPK,作为关键的驱动因素和治疗靶点。靶向药物的类别涵盖了关键药物,如西妥昔单抗、恩考芬尼和贝伐珠单抗,以及用于MSI-H疾病的ICIs,这些药物从根本上改变了mCRC的结局。尽管如此,重大的治疗障碍依然存在,包括肿瘤异质性以及原发性和获得性耐药的出现等。这些障碍,加上经济限制,需要持续的研究重点来阐明耐药机制、验证下一代预测性生物标志物,并开发创新的、逆转耐药的联合疗法。
尽管在理解CRC的分子基础方面取得了显著进展,但相互关联的障碍依然存在,限制了精准肿瘤学的临床影响。这些挑战要求持续、集中地研究耐药机制、预测性生物标志物和创新治疗方法。CRC的内在异质性,涵盖肿瘤间差异和瘤内克隆多样性,从根本上使通用治疗策略的开发复杂化。像空间多组学这样的技术是解决这种异质性的关键,但需要进一步成熟以克服当前在分辨率和数据整合方面的局限性。因此,生物信息学和计算生物学的进步对于开发必要的复杂分析工具以从海量多组学数据中提取临床意义至关重要。此外,实现全球精准肿瘤学公平性受到显著经济负担的限制。高通量多组学分析和随后的个性化药物的高成本构成了主要障碍。同时,缺乏能够忠实代表CRC TME的人源相关临床前模型,这突显了需要先进的模型系统来弥合从实验室到临床的转化差距。
应对CRC构成的全球健康挑战需要跨学科合作,联合实验科学家、临床医生和生物信息学家。对研发的持续投入对于将先进技术成功融入临床实践不可或缺。CRC的未来是复杂的,需要一种动态的、个性化的、以患者为中心的策略。将人工智能技术整合到肿瘤学中,将为诊断和治疗计划中的精准医疗带来新的可能性。通过将靶向治疗与AI相结合,并基于对CRC发病机制的深入了解,我们可以为显著改进的治疗方案奠定基础。通过这样专注、协作的努力,才有可能实现精准肿瘤学的潜力,推动该领域发展,并通过使CRC成为一种可控的疾病,为全球患者带来希望。
靶向治疗与免疫疗法、微环境调节和微生物干预的整合构成了一种强大的多模式治疗策略。通过分子分型和个性化医疗,有望提高患者护理的精准度,从而更好地识别那些最有可能获益的患者。然而,根本性挑战依然存在,包括耐药性的产生和单药治疗效果有限。虽然联合方法提供了理论上的益处,但诸如不可预测的药物相互作用和高度的患者间变异性等复杂性,常常限制临床疗效并可能加剧毒性。需要更深入的机制研究来充分阐明这些联合用药背后的协同机制,从而在减轻不良反应的同时优化治疗效果。
参考文献:Chen Y, Zhang J, Ding Y, Zhu F, Chen Y. Colorectal cancer pathogenesis, oncogenic signaling networks and targeted therapeutic advances. Mol Biomed. 2026 Mar 14;7(1):32. doi: 10.1186/s43556-026-00433-4. PMID: 41826571.
-
本微信公众号内容仅供医疗专业人士参考
不代表对任何药物或者治疗的推荐
由于时间和水平有限,如有错漏敬请谅解,
不吝赐教
(以上内容借助KIMI+/DeepSeek翻译整理所成)
加入毕凯今选粉丝群获取更多更新文献资讯~
2026-03-12
·百度百家
全球第二个获FDA批准进入临床的SHP2抑制剂,终于迈出联合用药关键一步。加科思宣布,JAB-3312与PD-1派姆单抗、MEK抑制剂比美替尼的联合疗法已分别完成首例患者给药,合作伙伴艾伯维同步支付2000万美元里程碑款项。至此,双方合作累计已交付首付款+里程碑超过2.2亿美元。
2020年5月,加科思把自研SHP2抑制剂(JAB-3068/JAB-3312)授权给艾伯维,交易总额超8.55亿美元,创下中国自主研发小分子抗肿瘤药海外专利授权最高纪录。目前,JAB-3312正在全球30余家中心展开单药及多组合试验,适应症覆盖实体瘤与血液瘤。除SHP2外,加科思还有KRAS、BET、五款IND-Enabling及五款临床前候选分子,管线密度业内领先。
康方生物(9926.HK)的CTLA-4单抗Quavonlimab(MK-1308)已正式进入III期临床,与Keytruda联合疗法对比标准方案一线治疗晚期肾透明细胞癌(ccRCC)。默沙东一次性支付2亿美元总对价,并承担后续里程碑与销售分成。
这不是康方第一次“卖药”。2015年,公司将Quavonlimab全球独家权利授权给默沙东,交易总额同样高达2亿美元。如今,该分子已在肾癌、肝癌、微卫星不稳定高结直肠癌、黑色素瘤等5大瘤种6项研究中验证疗效,联合Keytruda的数据尤为亮眼。
凭借密集管线与临床推进速度,康方在首届BioChina高峰论坛上获评“中国双抗药物十大领军企业”,创始人夏瑜与联合创始人王忠民分别摘得“年度十大药物创新领军人物”与“年度十大药物创新科学奖”。
和铂医药(2142.HK)宣布与哈佛医学院附属丹娜-法伯癌症研究院达成战略合作,共同推进双特异性抗体与CAR-T细胞疗法的发现与临床转化。双方将整合和铂Harbour Mice®全人源抗体平台与丹娜-法伯在CAR-T研发及肿瘤基础研究上的优势。
Harbour Mice®平台可快速筛选出亲和力高、成药性好的新型抗体,包括双特异、多特异、ADC及CAR-T等衍生物。合作初期,双方将聚焦实体瘤适应症,优先推进HBM4003——一款全人源单克隆抗CTLA-4重链抗体——与PD-1联合用药的I期研究。今年3月,HBM4003黑色素瘤适应症已完成首例给药;此次肺癌适应症启动,再次验证和铂在创新生物疗法上的推进速度。
由云杉医疗联合深圳市精准医疗学会共同发起的“云杉健康会”在深圳正式揭牌。首批创始会员涵盖企业主、金融高管及医生群体,未来将享受精准体检、私人医生、全生命周期健康管理、全球诊疗协调等一站式服务。
中心选址深圳湾核心片区——天利名城,引入代表国际尖端水平的PET/MR分子影像设备,并设置封闭式会员专享区。云杉整合心内、消化、肿瘤、骨科、中医康复等重点专科资源,可提供肿瘤手术、关节置换、腰椎椎管狭窄、O型腿矫正、胆囊微创等全链条手术与康复服务。随着南山健康管理中心的落成,云杉医疗正式向“国际预防医学与健康管理领航者”目标迈出关键一步。
作为国内最早一批医疗健康产业投资机构之一,高特佳投资已布局深圳、上海、北京、南京、香港等地,累计管理医疗健康基金超25支、资产规模逾200亿元,先后投资150余家企业,其中80余家聚焦医疗健康主赛道;累计推动26家企业成功上市。通过战略性股权投资、并购、PE/VC、天使等全阶段工具箱,高特佳持续为成员企业输送资本、渠道与专业能力,助力四大巨头在研发、临床与商业端同步提速。
免疫疗法细胞疗法临床3期引进/卖出抗体药物偶联物
2026-03-08
申康党委系统医务专家廖美琳:勇当中国肺癌研究领域排头兵写意人物丨陆舜教授:从PI的角度看中国创新药的发展陆舜教授:有的放矢,做肺癌研究领域的“领潮儿”陆舜教授:沃利替尼有望成为首个代表中国走向全球的肺癌靶向创新药物陆舜:引领中国抗肿瘤药物研发,将肺癌变成慢性病他的愿望:让肺癌变成慢性病——访上海市胸科医院肿瘤科主任陆舜教授陆舜教授:临床研究如何快速获批?创新思路从何而来?肿瘤临床研究开展“有方”医语舜间丨记录陆舜教授肺癌诊疗领域30余年的精彩瞬间上海交通大学附属胸科医院陆舜教授团队肺癌国内外适应症相关研究阶段汇总致敬经典,传承未来丨中国胸部肿瘤原创临床研究二十年心路历程(2007-2029)迎难而上丨中国胸部肿瘤临床试验转化研究长三角模式探索长三角肺癌团魂丨中国胸部肿瘤临床试验转化研究长三角模式探索迎难而上的亮剑精神胸部肿瘤 “工业革命” 成果阶段汇总丨中国胸部肿瘤临床试验转化研究长三角模式探索国际多中心临床研究阶段汇总丨世界胸部肿瘤历史上的长三角肺癌贡献江浙沪地区历届重要肺癌论坛汇总(长三角地区肺癌论坛系列)传承与发展丨长三角肺癌代表性著作汇总传承与发展丨长三角肺癌协作组指南/专家共识汇总读书笔记丨长三角肺癌之光-中国肺癌原创临床研究阶段汇总读书笔记丨长三角肺癌科技成果奖励阶段汇总直播课丨长三角肺癌协作组-罕见靶点临床试验招募全国巡讲系列汇总携手共赢丨长三角肺癌与知名药企临床研究合作成果阶段汇总长三角肺癌协作组青年博士团队组织框架介绍长三角肺癌之光丨星星之火可以燎原-基层肺癌协作组的前世今生共谋江苏省基层医院肺癌未来发展之路齐聚浙江嘉兴,长三角肺癌再续新篇章——浙江省基层肺癌协作组成立齐聚浙江丽水,为患者搭建长三角罕见肿瘤新药转化研究平台——浙江省基层罕见肿瘤协作组成立长三角肺癌之光丨金陵肺癌传承与发展长三角肺癌之光丨浙江肺癌传承与发展长三角肺癌之光丨南昌肺癌传承与发展杭州肺癌之光丨嘉兴肺癌(杭州肺癌-嘉兴支)传承与发展杭州肺癌之光丨湖州肺癌(杭州肺癌-湖州支)传承与发展杭州肺癌之光丨台州肺癌(杭州肺癌-台州支)传承与发展光荣榜丨长三角肺癌-国科大支(杭州肺癌)国际会议大会发言汇总历史回顾丨浙江省肿瘤医院胸部肿瘤青年医师精彩亮相2016WCLC长三角实体瘤病友交流群系列全面上线长三角罕见肿瘤社区建设
人文关怀浙江女子一天打2份工:只要老公活着,我吃多少苦都可以!背后故事看哭女儿出生第五天,29岁的她确诊肺癌晚期
门诊安排中国科学院大学附属肿瘤医院I期临床试验门诊开诊中国科学院大学附属肿瘤医院肿瘤内科资深专家张沂平教授I期临床试验门诊开诊
临床试验汇总
临床试验招募丨HA121-28片在晚期RET融合实体瘤患者中的最大耐受剂量探索及单、多次给药药代动力学研究I期临床试验临床试验招募丨阿美替尼一线治疗EGFR突变的肺腺鳞癌或腺癌和鳞癌混合成分癌的疗效和安全性研究:一项前瞻性、多中心、单臂临床研究临床试验招募丨评价BPI-572270胶囊在RAS突变的晚期实体瘤患者中安全性、耐受性、药代动力学和有效性的I/II期临床研究临床试验招募丨评价RGL-232在携带KRAS突变的晚期恶性实体瘤患者中的安全性、耐受性、药代动力学特征及免疫原性的开放标签、多中心I期临床研究临床试验招募丨评价SYN608在局部晚期或转移性实体瘤患者中的安全性、耐受性、药代动力学和初步抗肿瘤活性的首次人体I期研究临床试验招募丨评估注射用DXC014在多种恶性实体肿瘤患者中的安全性、耐受性、药代动力学特征以及初步疗效的开放、多中心、首次人体、剂量递增和扩大入组的I期临床研究临床试验招募丨TCC1727联合贝莫苏拜单抗/奥拉帕利/托泊替康治疗晚期实体瘤的开放性、多中心Ib/II期临床试验临床试验招募丨HRS-4508联合其他抗肿瘤治疗在HER2异常非小细胞肺癌受试者中的安全性、耐受性及有效性的开放、多中心的II期临床研究临床试验招募丨一项评估YL242单药治疗和联合治疗在晚期实体瘤受试者中的安全性、耐受性、药代动力学和有效性的I/II期、多中心、开放性、首次人体研究临床试验招募丨一项在晚期实体瘤患者中评价CS2009(靶向PD-1/VEGFA/CTLA-4的三特异性抗体)的安全性、耐受性、药代动力学和抗肿瘤活性的I期、剂量递增和剂量扩展研究临床试验招募丨一项评价注射用BAT7111在晚期实体瘤患者中的安全性、耐受性、药代动力学特征和初步临床有效性的多中心、开放的I/II期临床研究临床试验招募丨评价JAB-23E73用于KRAS基因改变的晚期实体瘤的安全性、耐受性、药代动力学以及初步抗肿瘤活性的多中心、开放性I/IIa期研究临床试验招募丨一项评估注射用ALK-N001在晚期实体瘤患者中的安全性、耐受性、药代动力学和有效性的开放性、多中心的I/II期研究临床试验招募丨注射用SKB445治疗晚期实体瘤受试者的安全性、耐受性、药代动力学和初步抗肿瘤活性的I期临床研究临床试验招募丨评估FP008在晚期实体瘤受试者中的安全性、疗效、药代动力学和药效学特征的首次人体1期研究临床试验招募丨一项评价RNK08954在中国KRAS G12D突变晚期实体瘤受试者中的安全性、耐受性、药代动力学特征以及初步抗肿瘤疗效的多中心、开放的I期临床研究临床试验招募丨一项在晚期/转移性实体瘤受试者中评估DB-1310安全性、耐受性、药代动力学和初步抗肿瘤活性的I/IIa期多中心、开放性首次人体研究临床试验招募丨评估注射用DXC006在多种实体瘤、血液瘤患者中的安全性、耐受性、药代动力学特征以及初步疗效的开放、多中心、首次人体、剂量递增和扩大入组的I期临床研究临床试验招募丨HRS-6719治疗甲硫腺苷磷酸化酶(MTAP)缺失的晚期实体瘤患者的安全性、耐受性、药代/药效动力学和有效性的开放、多中心I/II期临床研究临床试验招募丨评价LIT0922胶囊在晚期实体瘤患者中的安全性、耐受性、药代动力学及初步抗肿瘤活性的多中心、开放性I期临床试验临床试验招募丨评估AK137疗晚期恶性肿瘤患者的安全性、耐受性、药代动力学特征和初步抗肿瘤活性的I期临床研究临床试验招募丨一项评估NTQ3617片在晚期恶性实体肿瘤患者中的安全性、耐受性、药代/药效动力学特征及初步疗效的I期临床试验临床试验招募丨HRS-4508单药在HER2通路异常的晚期或转移性实体瘤患者中的安全性、耐受性及药代动力学的开放、多中心的I期临床研究临床试验招募丨一项在携带有KRAS G12C突变的晚期非小细胞肺癌患者中进行的Adagrasib单药或联合帕博利珠单抗的II期临床试验和一项Adagrasib联合帕博利珠单抗对比帕博利珠单抗的III期临床试验临床试验招募丨RX108-A片在晚期恶性实体肿瘤患者中的安全性、耐受性、药代动力学和初步疗效的I期研究临床试验招募丨评价GTA182在MTAPnull/lost的晚期实体瘤受试者中的安全性、耐受性的剂量递增和剂量扩展的I期临床研究临床试验招募丨一项在晚期实体瘤患者中评估ACR246安全性、耐受性、药代动力学 特征与有效性的开放、多中心、剂量递增和队列扩展的I/IIa期临床研究临床试验招募丨一项评价蛋白质降解剂注射用RNK05047在晚期实体瘤和淋巴瘤受试者中的安全性、耐受性、药代动力学、药效学和疗效的开放、多中心的Ⅰ期临床研究(CHAMP-1)临床试验招募丨一项评价SY-5933片在携带KRAS(G12C)突变的晚期实体瘤受试者中安全性、耐受性、药代动力学特征和初步疗效的剂量递增和剂量扩展的I期研究临床试验招募丨评价ASKC202片或联合ASK120067片在晚期实体瘤患者中的安全性、耐受性、药代动力学特征及初步有效性的开放、多中心、I期临床试验临床试验招募丨一项评估HB0052治疗晚期实体肿瘤受试者的安全性、耐受性、药代动力学特征以及初步疗效的开放性、多中心I/II期临床研究临床试验招募丨一项评价RNK08954在中国KRAS G12D突变晚期实体瘤受试者中的安全性、耐受性、药代动力学特征以及初步抗肿瘤疗效的多中心、开放的I期临床研究临床试验招募丨一项评估GFH375治疗KRAS G12D突变型晚期实体瘤患者的安全性、耐受性、药代动力学和疗效的开放标签、I/II期临床研究临床试验招募丨一项评估CT-3505胶囊对比克唑替尼胶囊在ALK阳性非小细胞肺癌患者中的有效性和安全性的随机、对照、多中心III期临床试验临床试验招募丨一项在晚期实体瘤患者中评估IBR822安全性、耐受性、药代动力学特征与有效性的开放、多中心、剂量递增和队列扩展的I/IIa期临床试验招募丨HRS-7058单药在KRAS G12C突变晚期实体瘤患者中的安全性、耐受性、药代动力学及疗效的开放、多中心的I期临床研究临床试验招募丨SHR-4849注射液在晚期恶性实体瘤患者中的安全性、耐受性、药代动力学和有效性的多中心、开放的I期临床研究临床试验招募丨一项评估HC010治疗晚期实体瘤患者的安全性、药代动力学和抗肿瘤活性的多中心、开放、剂量递增和剂量扩展I期临床研究临床试验招募丨3HP-2827在FGFR2异常的手术不可切除/转移性实体瘤患者中的安全性、耐受性、药代动力学和疗效的开放、多中心1/2期临床研究临床试验招募丨HRS-4642联合抗肿瘤药物在晚期实体瘤受试者中的安全性、耐受性及有效性的IB/II期临床研究临床试验招募丨注射用SHR-7631在晚期实体瘤患者中的安全性、耐受性、药代动力学及疗效的开放、多中心I期临床研究临床试验招募一项在晚期实体瘤患者中评估ABP1019A片的安全性、耐受性、药代动力学特征和初步疗效的剂量递增及扩展队列的开放性I/II期临床研究临床试验招募丨注射用AST2169脂质体在KRAS G12D突变晚期实体瘤患者中安全性、耐受性、药代动力学及初步疗效的I期临床研究临床试验招募丨评价HJ-002-03片在表皮生长因子受体(EGFR)突变的晚期非小细胞肺癌患者中的安全性、耐受性、药代动力学特征和初步抗肿瘤疗效的I期临床研究临床试验招募丨注射用FZ-AD005抗体偶联剂在晚期实体瘤患者中的安全性、耐受性、药代动力学和抗肿瘤疗效的I期临床研究临床试验招募丨HC006在实体瘤患者中的安全性耐受性药代动力学免疫原性和有效性研究:一项多中心开放单药剂量递增的I期临床试验临床试验招募丨评价ASKC202片或联合ASK120067片在晚期实体瘤患者中的安全性、耐受性、药代动力学特征及初步有效性的开放、多中心、I期临床试验临床试验招募丨评价GH21胶囊联合D-1553片治疗携带KRAS G12C突变的局部晚期或转移性实体瘤受试者的安全性、耐受性、药代动力学和疗效的多中心、开放、剂量递增及扩展的Ib/II期临床研究临床试验招募丨注射用SHR-A2102在晚期实体瘤患者中的安全性、耐受性、疗效及药代动力学的开放、单臂、多中心的 I 期临床研究临床试验招募丨一项评估JMT101注射液联合注射用多西他赛(白蛋白结合型)治疗局部晚期/复发或远处转移的鳞状细胞非小细胞肺癌患者的安全性、耐受性和初步疗效的随机、对照、开放II/III期临床研究临床试验招募丨一项在局部晚期或转移性实体瘤受试者中评价ANS014004单药治疗的安全性、耐受性、药代动力学和初步疗效的I期研究临床试验招募丨注射用SHR-9839联合抗肿瘤治疗在晚期实体瘤患者中的安全性、耐受性及有效性的开放、多中心IB/II期临床研究临床试验招募丨注射用LN005在恶性肿瘤患者中安全性、耐受性、药代动力学特征和有效性的I期临床研究临床试验招募丨一项在晚期实体瘤患者中评价AMT-116(一种抗CD44v9抗体偶联药物)的I/II期研究临床试验招募丨评价AMX3009治疗局部晚期或转移性非小细胞肺癌患者(罕见EGFR突变)的安全性、有效性、耐受性及药代动力学特征的Ib/II期临床试验临床试验招募丨一项评价BAT8008联合BAT1308在晚期实体瘤患者中的安全性、耐受性、药代动力学特征和初步疗效的多中心、开放的IB-II期临床研究临床试验招募丨BPI-442096片在晚期实体瘤患者中的I期临床研究临床试验招募丨注射用FZ-AD004抗体偶联剂在晚期实体瘤患者中的安全性、耐受性、药代动力学和抗肿瘤疗效的I期临床研究临床试验招募丨注射用ZGGS15在晚期实体瘤患者中的剂量递增、耐受性、安全性、药代动力学的I期临床研究临床试验招募丨盐酸ZG0895治疗晚期实体瘤患者的耐受性、安全性、药代动力学/药效学和初步疗效的开放、多中心、I期剂量递增和扩展试验临床试验招募丨Axl抑制剂FC084CSA片在晚期恶性实体肿瘤患者中的安全性、耐受性、药代动力学特征及初步疗效的I期临床研究临床试验招募丨评估QLP2117治疗晚期实体瘤患者的安全性、耐受性、药代动力学、免疫原性和有效性的开放、多中心I期临床研究临床试验招募丨评价SYS6010在晚期实体瘤患者中的安全性、耐受性、药代动力学特征和初步疗效的I期临床试验临床试验招募丨一项在HER2异常的晚期或转移性实体瘤患者中进行的BI 1810631单药治疗的开放性、I期、剂量递增(含剂量确证和扩展)试验临床试验招募丨注射用SHR-A1904在晚期实体瘤患者中的安全性、耐受性、药代动力学及疗效的开放、单臂、多中心的I期临床研究临床试验招募丨一项在携带KRAS p.G12C突变的晚期实体瘤受试者中评估D3S-001单药治疗的安全性、耐受性、药代动力学、药效学和初步疗效的开放性标签、剂量递增和剂量扩展的I期研究临床试验招募丨一项评价用注射用BAT8008在晚期实体瘤患者中的安全性、耐受性、药代动力学和初步疗效性的多中心、开放性I期临床研究临床试验招募丨注射用SG1408在晚期恶性实体瘤受试者中安全性、耐受性和初步有效性的I期临床研究临床试验招募丨一项评价注射用BAT8007治疗晚期实体瘤患者的安全性、耐受性和药代动力学特征的多中心、开放性I期临床研究临床试验招募丨XCCS605B在中国晚期实体瘤患者中的安全性、耐受性、药代动力学特征和疗效研究:一项多中心、开放、单药及联药剂量递增和联药剂量扩展的I/II期临床试验临床试验招募丨评估注射用TQB2103在晚期恶性肿瘤受试者中安全性、耐受性、药代动力学和初步有效性的I期临床试验临床试验招募丨评估TQB2223注射液联合派安普利单抗在晚期恶性肿瘤受试者中耐受性和药代动力学的I期临床试验临床试验招募丨一项评价GH2616片在晚期实体瘤受试者中的安全性、耐受性、药代动力学、药效动力学和初步疗效的剂量递增、剂量扩展的Ia/Ib期临床研究临床试验招募丨注射用重组抗EpCAM和CD3人鼠嵌合双特异性抗体(M701)胸腔输注联合全身治疗在晚期非小细胞肺癌合并恶性胸水患者中的药代/药效学特征、安全性、耐受性和初步疗效的多中心、开放的II期临床研究临床试验招募丨一项评价H002在表皮生长因子受体突变阳性的局部晚期或转移性非小细胞肺癌患者中的安全性、耐受性、药代动力学和初步抗肿瘤活性的开放性、剂量递增和扩展的I/IIa期临床研究临床试验招募丨注射用SHR-1826在晚期恶性实体瘤患者中的安全性、耐受性、药代动力学和有效性的多中心、开放的I期临床研究临床试验招募丨一项评估注射用DR30206治疗晚期实体瘤患者的安全性、耐受性和药代动力学特征的多中心、开放性I期临床研究临床试验招募丨注射用SHR-A1921联合抗肿瘤疗法治疗晚期实体肿瘤的开放、多中心II期临床研究临床试验招募丨一项在NTRK或ROS1基因融合成人晚期实体瘤患者中评估HG030片的安全性、耐药性、药代动力学特征与有效性的开放、多中心、剂量递增和扩大入组的I期临床研究临床试验招募丨评价7MW3711在晚期实体瘤患者中的安全性、耐受性、药代动力学特征和初步疗效的I/II期临床研究临床试验招募丨一项评价CTS2190胶囊在实体瘤患者中的安全性、耐受性、药代动力学和初步抗肿瘤活性的多中心、开放、剂量递增/扩展的临床研究临床试验招募丨评价注射用SKB410用于治疗晚期实体瘤受试者的安全性、耐受性、药代动力学和初步疗效的I期临床研究临床试验招募丨一项评估D-1553联合IN1008治疗KRAS G12C突变阳性的局部晚期或转移性肠癌受试者的安全性、耐受性、药代动力学和有效性的1b/2期研究临床试验招募丨注射用SHR-A1811联合SHR-1316在HER2突变、扩增或者过表达的晚期非小细胞肺癌受试者中的安全性、耐受性、药代动力学及有效性的IB/II期研究临床试验招募丨一项注射用SHR-9839在晚期实体瘤患者中的安全性、耐受性、药代动力学及疗效的开放、多中心I期临床研究临床试验招募丨一项评价FWD1509 MsOH在晚期非小细胞肺癌患者中的安全性耐受性药代动力学和抗肿瘤活性的开放性Ia期研究临床试验招募丨HRS2300单药及分别联合SHR-1316、SHR-1701、曲美替尼和阿美替尼在晚期恶性肿瘤患者中的剂量递增和剂量拓展的多中心、开放的I期临床研究临床试验招募丨一项评估重组人源化抗HER2单抗-Tub114偶联剂(DX126-262)在HER2阳性或HER2基因突变的不可切除的局部晚期或转移性非鳞非小细胞肺癌患者中的安全性、有效性及药代动力学特征的Ib/II期临床研究临床试验招募丨一项评估PRJ1-3024在晚期实体瘤患者中的安全性、耐受性、药代动力学、药效学和初步疗效的I/II期临床研究临床试验招募丨注射用LM-108单药或联合特瑞普利单抗在晚期实体瘤患者中的安全性、耐受性、药代动力学特征以及初步疗效的开放标签、剂量递增和剂量扩展的I/II期临床研究临床试验招募丨LVGN7409单药治疗局部晚期、转移性或复发难治恶性肿瘤的开放标签、I期试验临床试验招募丨HRS-4642注射液在携带KRAS G12D突变的晚期实体瘤受试者中的I期临床试验临床试验招募丨注射用QLF31907在晚期恶性肿瘤患者中的耐受性、安全性、药代动力学以及初步抗肿瘤活性的开放、剂量递增的Ia期临床研究临床试验招募丨ES014用于局部晚期或转移性实体瘤患者的开放标签、多中心、剂量递增和队列扩展的I期临床研究临床试验招募丨一项评价重组人源抗BTLA单克隆抗体(JS004)注射液联合特瑞普利单抗以及联合标准化疗在晚期肺癌患者中安全性、耐受性、药代动力学和初步疗效的Ib/II期临床研究临床试验招募丨一项在携带EGFR和/或HER2突变的晚期非小细胞肺癌受试者中评估BAY 2927088的开放、首次人体研究临床试验招募丨JYP0322在ROS1基因融合阳性局部晚期/转移性实体瘤患者中安全性、耐受性、药代动力学特征及初步有效性的I期临床研究临床试验招募丨一项阿美替尼联合SHR-1701等创新药物治疗EGFR突变的复发或晚期非小细胞肺癌开放、多中心的Ib/II期临床研究临床试验招募丨一项在携带RET突变或融合的不可切除、局部晚期或转移性实体瘤成人患者中研究APS03118的安全性、耐受性、药代动力学和初步疗效的I期研究临床试验招募丨一项在晚期实体瘤患者中评价QLS31904安全性、耐受性和药代动力学的I期研究临床试验招募丨评估HLX35在晚期或转移性实体瘤患者中的安全性、耐受性、药代动力学特征及初步疗效的I期临床研究临床试验招募丨M701胸腔输注联合全身治疗在晚期非小细胞肺癌合并恶性胸水患者中安全性耐受性和初步疗效的多中心Ib/II期临床研究
临床试验招募丨注射用SHR-A1811联合吡咯替尼或SHR-1316在HER2突变、扩增或者过表达的晚期非小细胞肺癌受试者中的安全性、耐受性、药代动力学及有效性的IB/II期研究
临床试验招募丨一项评估D-1553联合IN10018治疗KRAS G12C突变阳性的局部晚期或转移性实体瘤受试者的安全性、耐受性、药代动力学和有效性的1b/2期研究
临床试验招募丨一项评估D-1553联合治疗在KRAS G12C突变阳性的局部晚期或转移性非小细胞肺癌受试者中的安全性、耐受性、药代动力学和有效性的1b/2期研究
临床试验招募丨一项评估H002在表皮生长因子受体突变阳性的局部晚期或转移性非小细胞肺癌患者中的安全性、耐受性、药代动力学和初步抗肿瘤活性的开放性、剂量递增和扩展的1/IIa期临床研究临床试验招募丨一项评估MAX-402794(多靶点酪氨酸激酶抑制剂)联合KN046(抗PD-L1/CTLA-4双特异性抗体)在晚期/转移性实体瘤患者中的安全性、耐受性、药代动力学特征和有效性的I/II期临床研究临床试验招募丨评价LF0397片在晚期实体瘤患者中的安全性、耐受性、药代动力学特征及初步有效性的开放、多中心、Ia期临床试验临床试验招募丨评价注射用BL-B01D1在消化道肿瘤及其他实体瘤患者中的安全性、耐药性、药代动力学特征和初步疗效的I期临床研究临床试验招募丨一项开放标签的评价ABSK061在晚期实体瘤患者中的安全性、耐受性和药代动力学I期研究临床试验招募丨一项旨在评估D-1553在晚期或转移性实体瘤受试者中的安全性、耐受性、药代动力学和有效性的I/II期、开放研究
临床试验招募丨注射用金纳单抗单药及联合替雷利珠单抗治疗晚期恶性实体瘤的安全性、耐受性和药代动力学的I期及扩展临床研究
临床试验招募丨一项评价TGRX-326治疗ALK阳性或ROS1阳性晚期非小细胞肺癌患者中的安全性、耐受性、药代动力学及初步有效性的剂量递增及扩展的I期临床试验临床试验招募丨评价注射用RC118在Claudin18.2表达阳性的局部晚期不可切除或转移性恶性实体肿瘤患者中的开放、多中心I/IIa期临床研究临床试验招募丨注射用SHR-A1921在晚期恶性肿瘤受试者中的安全性、耐受性、药代动力学及疗效的开放、多中心的I期临床研究临床试验招募丨评价 FH-2001胶囊治疗晚期实体瘤患者的安全性、有效性、药代/药效动力学特征的开放多中心、I期临床研究临床试验招募丨评价JAB-21822用于KRAS p.G12C突变的晚期实体瘤的安全性、耐受性、药代动力学和抗肿瘤活性的多中心,开放,剂量递增及扩展的I/II期临床研究临床试验招募丨多中心、开放、剂量递增的I期临床研究评估注射用重组抗PD-L1和TGF-β双特异性抗体(Y101D)在转移性或局部晚期实体瘤受试者中的安全耐受性和药代/药效学特征
临床试验招募丨抗EGFR/c-Met双特异性抗体MCLA-129在晚期实体瘤患者中进行的评价安全性、药代动力学特征和抗肿瘤活性的I/II期临床研究
临床试验招募丨HMPL-295S1治疗晚期恶性实体瘤的I期临床研究
临床试验招募丨一项评价RET抑制剂SY-5007片在晚期实体瘤受试者中安全性、耐受性、药代动力学特征和有效性的I期研究
临床试验招募丨MAX-10181在晚期实体瘤患者中的安全性、耐受性及药代动力学I期临床研究临床试验招募丨IMM2510治疗晚期实体肿瘤的多中心、开放、剂量递增及队列扩展的I期临床研究临床试验招募丨评价SI-B001双特异性抗体注射液在局部晚期或转移性上皮肿瘤患者中的安全性耐受性初步疗效的I期临床研究(已结束)
临床试验招募丨评价JAB-3068联合特瑞普利单抗(JS001)用于晚期实体瘤患者的安全性、耐受性、药代动力学和抗肿瘤活性的多中心,开放,剂量递增及扩展的Ib/IIa期临床研究(已结束)
临床试验招募丨一项旨在评估D-1553在晚期或转移性实体瘤受试者中的安全性、耐受性、药代动力学和有效性的1期、开放研究
临床试验招募丨一项采用Durvalumab(MEDI4736)联合化疗和立体定向放疗(SBRT)治疗寡转移非小细胞肺癌(NSCLC)患者的研究(已结束)
临床试验招募丨评估金妥昔单抗注射液联合甲磺酸阿美替尼治疗表皮生长因子受体基因突变的IV期非小细胞肺癌患者的安全性、耐受性和有效性的Ib期及Ib期扩展研究(已结束)临床试验招募丨SH3809片在晚期实体瘤患者中的单、多次剂量递增的安全性、耐受性和药代动力学的I期临床研究临床试验招募丨APL-102胶囊治疗晚期实体瘤患者的安全性,耐受性及药代动力学的I期临床研究临床试验招募丨XZP-3621治疗中国ALK重排或ROS1重排的晚期非小细胞肺癌受试者的多中心、开放、剂量递增和剂量扩展的Ⅰ期临床研究(已结束)临床试验招募丨注射用SHR-A1811在HER2过表达、扩增或突变的晚期非小细胞肺癌患者中的安全性、耐受性、药代动力学及有效性的I/II期临床研究(SHR-A1811-I-103)临床试验招募丨一项多中心、开放、单臂 I 期剂量探索和 II 期扩展研究,评价FCN-011在晚期实体瘤(I 期)和 NTRK 融合阳性晚期实体瘤(II 期)患者中的安全性、耐受性、药代动力学特征和初步抗肿瘤活性
临床试验招募丨评价在中国晚期实体瘤患者中给予注射用Lurbinectedin(PM01183)单药的安全性、耐受性、 药代动力学特征及初步有效性的单臂、开放、剂量递增及扩展的I期临床研究(已结束)临床试验招募丨注射用RC108治疗c-MET阳性晚期恶性实体肿瘤患者的安全性、药代动力学和有效性的I期临床研究
临床试验招募丨金妥利珠单抗注射液治疗晚期恶性实体瘤和淋巴瘤的安全性、耐受性和药代动力学的Ia期临床研究(已结束)
临床试验招募丨评价 LZM009 治疗至少接受过一线化疗失败的复发或转移性胸腺癌患者的有效性和安全性的单臂、多中心、Ⅱ期研究
临床试验招募丨APG-2449口服治疗晚期实体瘤患者的安全性、药代动力学和药效动力学的I期临床研究
临床试验招募丨重组人源化抗BTLA单克隆抗体(JS004)在晚期实体瘤患者的I期临床研究
临床试验招募丨评价JMT101联合阿法替尼或奥希替尼治疗EGFR 20号外显子插入突变的ⅢB或Ⅳ期非小细胞肺癌患者的安全性和有效性的Ⅰb期临床研究
临床试验招募丨TQB3558片耐受性、有效性和药代动力学I期临床试验
临床试验招募丨评估口服谷美替尼(SCC244,一种高度选择性 c-Met 抑制剂)在具有 c-MET 改变的晚期非小细胞肺癌患者中有效性和安全性的多国、多中心、开放 Ib/II 期临床研究(已结束)
临床试验招募丨多中心、开放、非随机I期临床研究,评价口服HS-10340在晚期实体瘤患者中的安全性、耐受性和药代动力学
临床试验招募丨多中心、开放、单臂的剂量爬坡和扩展的I期临床研究:评价CYH33在晚期实体瘤患者中的安全性、耐受性、药代动力学特征和初步疗效
临床试验招募丨在EGFR突变型的晚期非小细胞肺癌患者中评估TY-9591片安全性、耐受性、药代动力学和药效学的单臂、开放、剂量爬坡和剂量扩展I期临床研究(已结束)
临床试验招募丨一项在胸腺癌受试者中评估KN046的疗效、安全性和耐受性的II期、开放性、多中心研究(已结束)
临床试验招募丨一项在不可切除的局部晚期或转移性实体瘤患者中评估抗TIGIT单克隆抗体BGB-A1217与抗PD-1单克隆抗体替雷利珠单抗(BGB-A317)联合用药治疗的安全性、耐受性、药代动力学和抗肿瘤活性的1/1b期研究
临床试验招募丨评价QL1706在晚期恶性实体肿瘤患者中有效性和安全性的多中心、开放的Ⅰb期研究(已结束)
临床试验招募丨抗LAG3抗体SHR-1802在晚期恶性肿瘤患者中的耐受性、安全性及药代动力学特征的I期临床研究(已结束)
临床试验招募丨评估 HL-085联合维莫非尼治疗BRAF V600突变的晚期实体瘤患者的安全性和药代动力学的单臂、剂量爬坡和扩展的I期临床研究
临床试验招募丨评价注射用重组人源化双功能单克隆抗体MBS301治疗HER2阳性复发或转移性恶性实体肿瘤的安全性、耐受性和药代动力学的开放、剂量递增的I期临床研究
临床试验招募丨评价SIM1803-1A在NTRK、ROS1或ALK基因融合的局部晚期/转移性实体瘤患者中的安全性、有效性及药代动力学特征的开放、多中心I期临床研究(已结束)
临床试验招募丨评价JAB-3312用于晚期实体瘤患者的安全性、耐受性、药代动力学和抗肿瘤活性的多中心开放,剂量递增的1期临床研究(已结束)
临床试验招募丨重组人源化抗PD-1单克隆抗体注射液在晚期实体瘤患者中的安全性、耐受性及药代动力学的Ia期临床试验
临床试验招募丨评价全人源抗PD-L1抗体注射液(LDP)在晚期恶性肿瘤患者中的安全性、 耐受性和药代动力学特征的I期临床试验
思维导图
科室简介
I期病房于2018年12月正式组建,2020年7月作为独立病房运行
主要从事抗肿瘤药物的I期临床试验,是浙江省省内唯一、国内较早开设的专业肿瘤I期临床试验病房,也是国内最重要的I期临床研究中心之一,立项数量和入组患者数均位居国内前列。2021年度出院2072人次,新立项I期项目72项。
目前有固定床位14张,加床4张,抢救床位1张,并设有医学检查室、活动室、沟通室、样本处理和储存室、药物储存和准备室、CRA/CRC办公室、备餐间、档案室、抢救室等区域,为参加各项新药临床试验的受试者提供周到的医疗服务。
科室设有专业的临床研究团队,拥有专职研究医生6人,主任医师1人,副主任医师1人,住院医师3人,医生助理1人,其中博士学历2人,硕士学历3人。研究生导师2名,在读研究生5名;拥有专职研究护士11人,均为主管护师,所有人员均接受GCP相关培训和专业技术培训,具备丰富的I期临床试验经验。
科室目前开展业务包括:项目立项审核,研究方案设计、项目阶段讨论,患者筛选入组,临床研究医生培训,临床科研训练等,是国内为数不多的集研究项目设计、立项审核、患者筛选和临床管理、科研培训于一体的临床研究中心。
科室自组建以来,已承接和完成抗肿瘤药物I期临床研究近百项,包括新药的首次人体试验(FIH)、临床药代动力学(PK)、药物相互作用(DDI)等。研究领域涵盖常见实体肿瘤和血液肿瘤,包括免疫治疗、靶向治疗、化疗、抗体药物等。在I期临床研究方面积累了丰富的经验。
走进中国科学院大学附属肿瘤医院(浙江省肿瘤医院)I期临床试验病房【所言医语】冲分能手!一名奔跑在科研路上的临床医生猎药人系列 | 浙江省肿瘤医院宋正波:3年诊治患者超千人,把好新药进临床的首道关口浙江省肿瘤医院宋正波主任:用“浙商精神”推动临床研究 | 创新100人35under35星光故事繁星篇02|从“六张床”到“领头羊”:宋正波教授的I期病房“创业”记中国科学院大学附属肿瘤医院(浙江省肿瘤医院)I期临床试验病房进修人员招聘公告中国科学院基础医学与肿瘤研究所/中国科学院大学附属肿瘤医院(浙江省肿瘤医院)宋正波课题组博士后招聘公告
推动行业前行的力量-医学创新专家
他是创新药进入临床的第一道“把关人”。目前我国创新药物发展迅速,亟待更多的创新模式进行快速临床转化,他自担任中国科学院大学附属肿瘤医院I期临床试验病房主任后,积极创新发展模式,目前病房开展肿瘤I期临床研究数量位居国内前列,已经成为中国重要的肿瘤创新药物临床研究基地,推动了中国肿瘤创新药物临床研究的开展。
——宋正波
推动行业前行的力量-十大医学创新专家
临床试验病房医生谈肿瘤早期临床研究长三角肺癌与药物临床试验质量管理规范(GCP)的故事
I期浙肿行
I期浙肿行(第一期)(2023.3.24)
战略合作
战略合作签约
第四届中国杭州临床试验高峰论坛上中国科学院大学附属肿瘤医院院长、中国科学院基础医学与肿瘤研究所所长谭蔚泓院士代表医院和阿斯利康、百济神州、恒瑞、罗氏、齐鲁、正大天晴等6家单位进行战略合作签约。
陈明常务副院长表示,此次签约的6家企业,都有重大临床试验的主要研究者PI落户中国科学院大学附属肿瘤医院,希望以此为样板,把科学家、医务人员、企业家汇聚在一起,共同建设和发展中国的医药事业。
历届杭州临床试验发展论坛汇总
2021年第五届中国杭州临床试验半山论坛(2021.11.13)
2020年第四届中国杭州临床试验高峰论坛(2020.12.11-2020.12.13)
2019年第三届中国杭州临床试验发展论坛(2019.12.13-2019.12.15)
2018年第二届中国杭州临床试验发展论坛(2018.12.7-2018.12.9)
2017年首届中国杭州临床试验发展论坛(2017.12.8-2017.12.9)
ECLUNG新生代共识/指南系列
由ECLUNG新生代主导执笔的国际/全国专家共识/指南系列,相继在Innovation,Cancer,Thorac Cancer等国际知名期刊发表。
临床研究
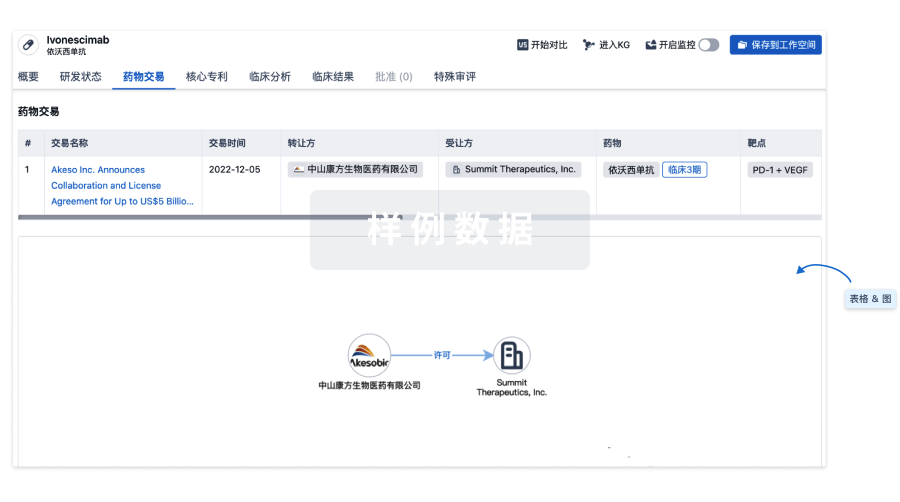
100 项与 JAB-3068 相关的药物交易
登录后查看更多信息
研发状态
10 条进展最快的记录, 后查看更多信息
登录
| 适应症 | 最高研发状态 | 国家/地区 | 公司 | 日期 |
|---|---|---|---|---|
| 实体瘤 | 临床2期 | 美国 | 2022-07-14 | |
| 胼胝体发育不全 | 临床2期 | 中国 | 2022-01-30 | |
| 鳞状细胞癌 | 临床2期 | 中国 | 2021-07-23 | |
| 头颈部鳞状细胞癌 | 临床2期 | 中国 | 2021-04-16 | |
| 食管癌 | 临床2期 | 中国 | 2018-11-20 | |
| 食管鳞状细胞癌 | 临床2期 | 中国 | 2018-11-20 | |
| 转移性实体瘤 | 临床2期 | 中国 | 2018-11-20 | |
| 非小细胞肺癌 | 临床2期 | 中国 | 2018-11-20 | |
| 非小细胞肺癌 | 临床2期 | 中国 | 2018-11-20 | |
| 晚期恶性实体瘤 | 临床2期 | 中国 | 2018-11-06 |
登录后查看更多信息
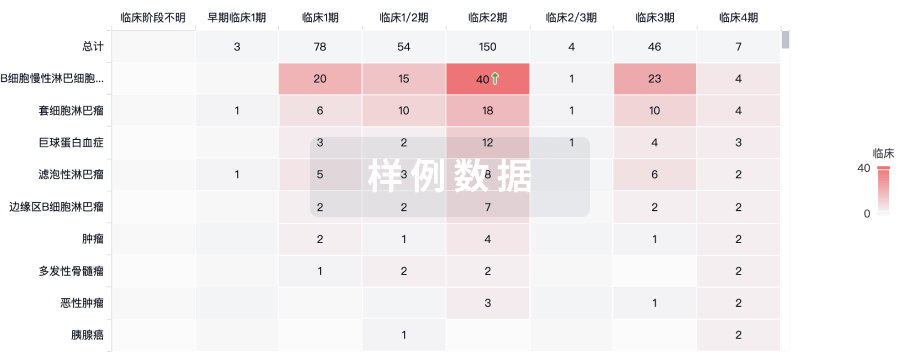
临床结果
临床结果
适应症
分期
评价
查看全部结果
| 研究 | 分期 | 人群特征 | 评价人数 | 分组 | 结果 | 评价 | 发布日期 |
|---|
No Data | |||||||
登录后查看更多信息
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
核心专利
使用我们的核心专利数据促进您的研究。
登录
或
临床分析
紧跟全球注册中心的最新临床试验。
登录
或
批准
利用最新的监管批准信息加速您的研究。
登录
或
特殊审评
只需点击几下即可了解关键药物信息。
登录
或
生物医药百科问答
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用