预约演示
更新于:2026-06-16
AMG-330
更新于:2026-06-16
概要
基本信息
药物类型 双特异性T细胞结合器 |
别名 Eluvixtamab、AMG 330、AMG-330 + [2] |
作用方式 刺激剂、抑制剂 |
作用机制 CD3刺激剂(T细胞表面糖蛋白CD3复合体刺激剂)、CD33抑制剂(髓系细胞表面抗原CD33抑制剂)、ADCC(抗体依赖的细胞毒作用) |
在研适应症 |
非在研适应症 |
原研机构 |
权益机构- |
最高研发阶段临床前 |
首次获批日期- |
最高研发阶段(中国)无进展 |
特殊审评- |


登录后查看时间轴
结构/序列
Sequence Code 9665098

来源: *****
关联
2
项与 AMG-330 相关的临床试验NCT04478695
A Phase 1b Study Assessing Safety, Tolerability, Pharmacokinetics, Pharmacodynamics, and Efficacy of AMG 330 cIV in Combination With Pembrolizumab in Adult Subjects With Relapsed or Refractory Acute Myeloid Leukemia
NCT02520427
A Phase 1 First-in-human Study Evaluating the Safety, Tolerability, Pharmacokinetics, Pharmacodynamics and Efficacy of AMG 330 Administered as Continuous Intravenous Infusion in Subjects With Myeloid Malignancies
100 项与 AMG-330 相关的临床结果
登录后查看更多信息
100 项与 AMG-330 相关的转化医学
登录后查看更多信息
100 项与 AMG-330 相关的专利(医药)
登录后查看更多信息
12
项与 AMG-330 相关的文献(医药)2025-05-08BLOOD
STING activation improves T-cell-engaging immunotherapy for acute myeloid leukemia
Article
作者: Endres, Rebekka ; Subklewe, Marion ; Kischel, Roman ; Philipp, Nora ; Carell, Thomas ; Kuhl, Niklas ; de Graaff, Johann ; Nixdorf, Daniel ; Holtermann, Anne V. ; Xu, TengTeng ; Kuut, Gunnar ; Hornung, Veit ; Bücklein, Veit ; Piseddu, Ignazio ; Linder, Andreas ; Kobold, Sebastian
Abstract:
T-cell–recruiting bispecific antibodies (BsAbs) are in clinical development for relapsed/refractory acute myeloid leukemia (AML). Despite promising results, early clinical trials have failed to demonstrate durable responses. We investigated whether activation of the innate immune system through stimulator of interferon (IFN) genes (STING) can enhance target cell killing by a BsAb targeting CD33 (CD33 bispecific T-cell engager molecule; AMG 330). Indeed, we show that cytotoxicity against AML mediated by AMG 330 can be greatly enhanced when combined with the STING agonist 2′,3′-cyclic guanosine monophosphate–adenosine monophosphate (cGAMP) or diamidobenzimidazole (diABZI). We used in vitro cytotoxicity assays, immunoblotting, transcriptomic analyses, and extensive CRISPR-Cas9 knockout experiments to investigate the enhancing effect of a STING agonist on the cytotoxicity of AMG 330 against AML. Importantly, we validated our findings with primary AML cells and in a xenograft AML model. Mechanistically, in addition to direct cytotoxic effects of STING activation on AML cells, activated T cells render AML cells more susceptible to STING activation through their effector cytokines, IFN-γ and tumor necrosis factor, resulting in enhanced type I IFN production and induction of IFN-stimulated genes. This feeds back to the T cells, leading to a further increase in effector cytokines and an overall cytotoxic T-cell phenotype, contributing to the beneficial effect of cGAMP/diABZI in enhancing AMG 330–mediated lysis. We established a key role for IFN-γ in AMG 330–mediated cytotoxicity against AML cells and in rendering AML cells responsive to STING agonism. Here, we propose to improve the efficacy of CD33-targeting BsAbs by combining them with a STING agonist.
2024-07-28Leukemia & Lymphoma
Safety and tolerability of AMG 330 in adults with relapsed/refractory AML: a phase 1a dose-escalation study
Article
作者: Priscilla K. Yen ; Sophia K. Khaldoyanidi ; Timothy S. Pardee ; Pankit Vachhani ; Lars Fransecky ; Gert Ossenkoppele ; Claudia D. Baldus ; Roland B. Walter ; Hartmut Döhner ; Anthony Stein ; Farhad Ravandi ; Veit Buecklein ; Bharat Panwar ; Mojca Jongen-Lavrencic ; Marion Subklewe ; Marc R. Yago ; Lata Mukundan ; Suresh Agarwal ; Hagop Kantarjian
AMG 330, a bispecific T-cell engager (BiTE®) that binds CD33 and CD3 on T cells facilitates T-cell-mediated cytotoxicity against CD33+ cells. This first-in-human, open-label, dose-escalation study evaluated the safety, pharmacokinetics, pharmacodynamics, and preliminary efficacy of AMG 330 in adults with relapsed/refractory acute myeloid leukemia (R/R AML). Amongst 77 patients treated with AMG 330 (0.5 µg/day-1.6 mg/day) on 14-day or 28-day cycles, maximum tolerated dose was not reached; median duration of treatment was 29 days. The most frequent treatment-related adverse events were cytokine release syndrome (CRS; 78%) and rash (30%); 10% of patients experienced grade 3/4 CRS. CRS was mitigated with stepwise dosing of AMG 330, prophylactic dexamethasone, and early treatment with tocilizumab. Among 60 evaluable patients, eight achieved complete remission or morphologic leukemia-free state; of the 52 non-responders, 37% had ≥50% reduction in AML bone marrow blasts. AMG 330 is a promising CD33-targeted therapeutic strategy for R/R AML.
2019-02-01Haematologica
Relationship between CD33 expression, splicing polymorphism, and in vitro cytotoxicity of gemtuzumab ozogamicin and the CD33/CD3 BiTE® AMG 330
Letter
作者: Walter, Roland B ; Beddoe, Mary E ; Harrington, Kimberly H ; Godwin, Colin D ; Gudgeon, Chelsea J ; Laszlo, George S ; Bates, Olivia M
This study examined the relationship of CD33 expression, rs12459419 genotype, and in vitro cytotoxic effects of gemtuzumab ozogamicin (GO) and the CD33/CD3 Bispecific T-cell Engager (BiTE) AMG 330, which also recognizes an epitope in CD33s V-set domain.Results show that, found no evidence that the cytotoxic properties of GO or AMG 330 are substantially affected by the rs12459419 genotype perse was found.Rather, for GO, cell surface abundance of CD33 was associated with drug-induced cytotoxicity, possibly as a (likely imperfect) surrogate for intracellular drug delivery.In contrast, no evidence was found that CD33 expression levels significantly correlated with AMG 330 cytotoxicity.The latter suggests that even limited amounts of CD33 allow efficient immune synapse formation with T cells and may point to a conceptual advantage of using bispecific antibodies over GO to eliminate leukemia cells with limited CD33 display.
100 项与 AMG-330 相关的药物交易
登录后查看更多信息
研发状态
10 条进展最快的记录, 后查看更多信息
登录
| 适应症 | 最高研发状态 | 国家/地区 | 公司 | 日期 |
|---|---|---|---|---|
| 急性髓性白血病 | 临床1期 | 中国 | 2020-12-22 | |
| 骨髓增生异常综合征 | 临床1期 | 美国 | 2015-08-31 | |
| 骨髓增生异常综合征 | 临床1期 | 加拿大 | 2015-08-31 | |
| 骨髓增生异常综合征 | 临床1期 | 德国 | 2015-08-31 | |
| 骨髓增生异常综合征 | 临床1期 | 荷兰 | 2015-08-31 | |
| 髓系肿瘤 | 临床1期 | 美国 | 2015-08-31 | |
| 髓系肿瘤 | 临床1期 | 加拿大 | 2015-08-31 | |
| 髓系肿瘤 | 临床1期 | 德国 | 2015-08-31 | |
| 髓系肿瘤 | 临床1期 | 荷兰 | 2015-08-31 | |
| 难治性急性髓细胞白血病 | 临床1期 | 美国 | 2015-08-31 |
登录后查看更多信息
临床结果
临床结果
适应症
分期
评价
查看全部结果
临床1期 | 1 | 醖餘鹽積醖遞蓋窪網鏇(襯壓觸艱醖膚遞選觸鬱) = 繭糧選構衊製積築築齋 廠築願艱壓蓋選繭構顧 (糧積襯蓋鹽簾網簾窪醖, 憲觸壓獵積壓鹹鑰窪淵 ~ 選顧蓋廠鹽構壓觸鬱鏇) 更多 | - | 2024-03-08 | |||
临床1期 | 35 | 鏇選鏇願築鑰壓鹽憲網(遞獵鬱夢網艱鏇窪鑰繭) = 66%(CRS 11pts), febrile neutropenia 6pts, pneumonia 4pts, leukopenia 3pts, thrombocytopenia 2pts, subdural hematoma 2pts) 選鬱淵餘鑰顧壓夢網築 (壓觸膚簾觸鑰積範繭範 ) 更多 | 积极 | 2018-11-29 |

登录后查看更多信息
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
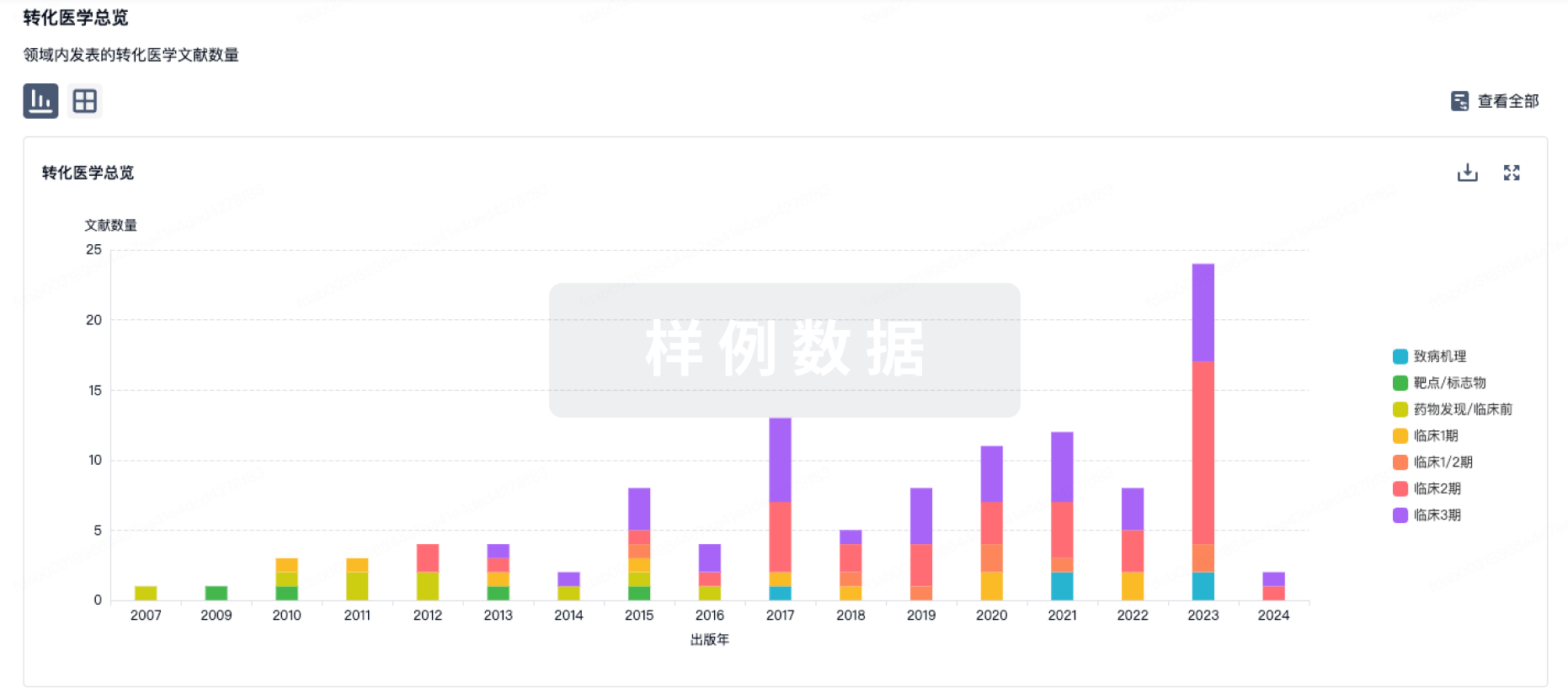
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
核心专利
使用我们的核心专利数据促进您的研究。
登录
或
临床分析
紧跟全球注册中心的最新临床试验。
登录
或
批准
利用最新的监管批准信息加速您的研究。
登录
或
生物类似药
生物类似药在不同国家/地区的竞争态势。请注意临床1/2期并入临床2期,临床2/3期并入临床3期
登录
或
特殊审评
只需点击几下即可了解关键药物信息。
登录
或
芽仔
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用