预约演示
更新于:2025-05-07
中国科学院上海有机化学研究所
教育|中国上海市
教育|中国上海市
更新于:2025-05-07
概览
标签
肿瘤
皮肤和肌肉骨骼疾病
呼吸系统疾病
小分子化药
蛋白水解靶向嵌合体(PROTAC)
疾病领域得分
一眼洞穿机构专注的疾病领域

暂无数据
技术平台
公司药物应用最多的技术
暂无数据
靶点
公司最常开发的靶点
暂无数据
| 排名前五的药物类型 | 数量 |
|---|---|
| 小分子化药 | 6 |
| 蛋白水解靶向嵌合体(PROTAC) | 3 |
关联
13
项与 中国科学院上海有机化学研究所 相关的药物作用机制 AXL抑制剂 [+1] |
最高研发阶段临床2期 |
首次获批国家/地区- |
首次获批日期- |

靶点 |
作用机制 FGFR2拮抗剂 |
在研机构 |
在研适应症 |
非在研适应症- |
最高研发阶段临床前 |
首次获批国家/地区- |
首次获批日期- |
作用机制 SARS-CoV-2 PLpro抑制剂 |
原研机构 |
在研适应症 |
非在研适应症- |
最高研发阶段临床前 |
首次获批国家/地区- |
首次获批日期- |
100 项与 中国科学院上海有机化学研究所 相关的临床结果
登录后查看更多信息
0 项与 中国科学院上海有机化学研究所 相关的专利(医药)
登录后查看更多信息
142
项与 中国科学院上海有机化学研究所 相关的新闻(医药)2025-04-30
·知识分子
4.30知识分子The Intellectual 导读当地时间4月30日,美国国家科学院(National Academy of Sciences)公布了2025年新当选院士名单。今年共有120位院士和30位外籍院士入选,其中包括中国中医科学院青蒿素研究中心研究员屠呦呦。现年94岁的她,因发现青蒿素治疗疟疾的突破性贡献,于2015年获得诺贝尔生理学或医学奖。她还获得过国家最高科学技术奖、并被授予共和国勋章。不过,她仍未当选中国科学院或中国工程院院士。2015年诺奖揭晓后,公众对屠呦呦未能入选院士的质疑随之而来。获奖后接受《纽约时报》采访时,屠呦呦表示自己过去曾多次申请中国科学院院士,但未能成功。“卫生部长部长曾亲自推荐我当院士。但是,有许多因素在起作用。情况很复杂”。青蒿素发现于大型研究抗疟疾药物的“523任务”中,这个项目规模大、参与人员众多,关于屠呦呦能否成为这个项目成果的代表性人物,一直存在争议。饶毅与合作者曾在《中国科学》发表文章,基于原始中文论文、文件和访谈资料,他们肯定屠呦呦为“发现青蒿素的代表性人物”,在青蒿素的发现过程中起了“关键作用”,青蒿素发现是“中药的科学研究丰碑”。尽管作出如此原创性且具有全球影响的贡献,屠呦呦在很长一段时间内并没有获得国内重要奖项或是类似院士等重要荣誉头衔。诺奖之后,屠呦呦依然未能入选两院院士。年龄或许是一个限制因素。屠呦呦获奖时已84岁,而中国科学院和中国工程院对院士增选设有一定的年龄门槛。根据2014年修订的《中国科学院院士增选工作实施细则》,65周岁以上的候选人需由6名或以上院士推荐,且其中至少4人须来自候选人申请学部,推荐方为有效。2023年改革后对这一标准有所调整,但还是要求至少两位院士推荐。尽管这一规定并未将候选人年龄严格限定在65岁以下,但也为高龄候选人增选院士制造了障碍。以2023年为例,中国工程院院士增选的有效候选人中,最年长者77岁。中国科学院方面,最年长的候选人为68岁。而美国国家科学院院士的入选没有年龄限制。纵观屠呦呦的荣誉历程,国际科学界的认可始终走在国内前面。她先是在2011年、2015年先后获得拉斯克奖与诺贝尔奖,随后才在2016年,数度无缘之后终于获颁国家最高科学技术奖。如今,获得诺奖十年后,先授予这位中国科学家院士荣誉的,仍然是美国国家科学院。2009年4月21日,正在北京大学医学部读硕士学位研究生的黎润红(现任职于北京大学医学人文研究院),在饶毅和张大庆教授指导下,以青蒿素为科学史研究课题,曾在中国中医科学院中药研究所对屠呦呦进行了访谈。访谈中,屠呦呦详细回顾了青蒿素发现的过程,《知识分子》获黎润红授权刊发访谈实录。撰文 | 黎润红● ● ●黎=黎润红屠=屠呦呦黎:请问您当时是怎么参加“523任务”的?主要做了哪些方面的工作?屠:青蒿素研究时间长、牵涉面宽,好在我们编了一本《青蒿及青蒿素类药物》,刚刚出版,现送你们三本,一本请留给饶毅和你、另外两本请转给我母校,你可回去看看,书里有比较详细的叙述,现只能概要地聊聊。我们中药所1967年没有参加“523任务”。早期,中医研究院广安门医院和针灸所搞针灸的同事们参加了。1969年,“523办公室”的领导到中医研究院,我们领导负责接待工作。“523办公室”的领导说:中央领导十分重视抗疟防治的研究工作,为解决问题,希望你们能参加此项任务。院领导当即表示,虽然处于文化大革命,中国中医研究院科研工作全部停顿,但我们不能推辞,要尽最大努力承担工作。后来就指定我为课题组长。当时就是这样一个情况。1969年2月,我先开始收集历代的文献资料、收集单秘验方,从中挑选出要研究的中药,提取后送236部队(军事医学科学院)做药效。当时,我们还没动物试验,到1970年年中,筛了百余种,包括青蒿在内,都没有理想结果。后来,236因其他任务也不再协助我们了,我们这个组也就停了。直到1971年5月广州会议以后,我们这个组经过调整和加强,才重新开展工作。1971年下半年,开始筛了一些中草药、也复筛了一些,都没有大的进展,后在《肘后方》“青蒿一握、绞汁服”的启示下,将青蒿用低沸点溶剂提取,得到“醚中干”[1](乙醚提取的中性部分)的化学有效部位,当年10月,鼠疟试验全部转阴,年底猴疟也得相同效果;1972年8、9月在海南和北京用青蒿“醚中干”治疗30例间日疟、恶性疟患者,获满意效果。1972年11月,在“523”的全国性会议上做了汇报。与此同时,我们对“醚中干”进行活性成分分离,1972年11月从中获得几个结晶,12月初,鼠疟确认其Ⅱ号结晶是活性成分,曾称“青蒿素Ⅱ”,后定名为“青蒿素”,应用至今。几十毫克剂量就使疟原虫全部转阴。1972年11月得到青蒿素后,即着手其化学结构鉴定研究。经确定其分子式分子量等,并证实其不含氮元素,又结合四大光谱数据,推定为倍半萜类化合物,属于新结构类型抗疟药。为争取时间确定其分子结构,就想找条件好的单位协作,了解到中科院上海有机所刘铸晋对倍半萜类化合物的研究有较多经验,后又了解到中科院生物物理所有先进的X线衍射,先后请他们合作研究青蒿素的结构,到1975年底,确定了青蒿素的立体构型。当然,不是什么事情都会一帆风顺的。有人说我们1973年做的青蒿素不成功,临床效果不好。1973年我们开展对青蒿素化学研究以外,还从北京产的青蒿中大量提取青蒿素。因为我们剂型室没恢复工作,所以提出来的青蒿素是拿出去外加工做的片剂。同年9月到海南疟区对恶性疟患者进行临床试用,结果5例患者,仅1例有效,效果不确切。经研究发现这些片子敲都敲不碎,片剂的崩解度有问题,又赶着将青蒿素原粉直接装胶囊,改用胶囊剂。副所长章国镇同志亲自带着胶囊赴海南昌江现场,做了几例。因为季节比较晚了,海南地区工作即将收尾,只做了3例间日疟,效果都是很好的。证明青蒿素抗疟有效。后来我院医疗队用我们提取的青蒿素继续在全国做了380多例,全部有效。附注一:1973年北京中药所提取的青蒿素II临床试验情况在进行青蒿素Ⅱ的临床试用前,由于急性动物试验的结果存在一定的问题,所内部对青蒿素Ⅱ是否可以进行临床使用产生了分歧。后经3人进行临床试服,未发现明显问题后于当年9月份赴海南开展临床试用。以下为当年的临床试验结果:“1973年9-10月,北京中药所用提取出的青蒿素在海南昌江对外地人口间日疟及恶性疟共8例进行了临床观察,其中外来人口间日疟3例。胶囊总剂量3-3.5g,平均退热时间30小时,复查3周,2例治愈,1例有效(13天原虫再现)。外来人口恶性疟5例,1例有效(原虫7万以上/mm3,片剂用药量4.5g,37小时退热,65小时原虫转阴,第6天后原虫再现);2例因心脏出现期前收缩而停药(其中1例首次发病,原虫3万以上/mm3,服药3g后32小时退热,停药1天后原虫再现,体温升高),2例无效”。屠呦呦口述的“后来我院医疗队用我们提取的青蒿素继续在全国做了380多例,全部有效。”指的是从1975年成都会议之后,全国进行大协作对青蒿素及其各种制剂进行临床验证时,中医研究院医疗队所做的总病例数。1973年临床试验回来以后就向“523办公室”汇报了。汇报以后,1974年1月,他们就说要开一个中西医结合寻找新药的会议。这个“523办公室”当时起到组织领导作用,因为当时文化大革命全部瘫痪了,只有部队来抓,而且任务期间领导关系也有变化,成为三部一院的领导,后来参加的单位越来越多。中医中药组实际上开始重点抓的是针灸,后来才是中药,中药抗疟传统多用常山,这个常山生物碱有效是有效,但有呕吐副作用,不好用到临床去,所以曾经做过好多改造结构的衍生物。西方1820年从金鸡纳树皮中提取出奎宁,后又研发好多衍生物,如阿的平、乙胺嘧啶,伯氨喹啉,最后的王牌氯喹,都是含氮的喹啉类药物,用了一百多年,已经产生抗药性了。我去了两次海南岛现场,感染疟疾的患者40多度高烧不退,又是恶性疟,后来转为脑型疟。好多年轻同志身体挺棒的,得了疟疾40度以上高烧,两天以后嘴唇都紫了,要不给他药吃,那不就送命了。当时大家都期望搞出一个全新结构的、疗效也很好的药物,来解决氯喹抗药性问题。国外,据报道美国在华盛顿陆军研究所以改造结构为主,有一个比较好的甲氟喹,但还是喹啉类的化合物。国内来说,实际上这个项目60年代中期,就是一个军工项目,军事医学科学院已经做了不少工作。1967年成立“523办公室”,全国七大省市都来做这个项目。据报道已经筛了化合物、中草药等4万多种。我从北医毕业后正好中医研究院建院,就来了中医研究院中药研究所工作。当年毛主席提出来要西学中,所以那时候办了一个西学中班,希望有西医工作经验的参加学习。我从北医毕业后,来到中医研究院中药所,学了两年半的西学中[2]。确实,学完了之后还是很有收获的。当时一些中医老大夫从全国各地请来的,像蒲辅周[3],他后来成为周总理的保健医生。青蒿素是一个原始性创新,因为它是一个全新的结构,而且疗效也很好。解决了抗性疟疾的治疗问题。所以大家比较重视,也是因为这个道理。但工作也不是一帆风顺的,从我的思路来说,中国有5000年的悠久历史,对医学也是比较重视的。我就开始搜集古代的书籍,地方的经验,我这有一本册子(疟疾单秘验方集)给你看,这是我69年做的。那时候我做了200多张卡片。黎:是否还有卡片?屠:你看看时间你就知道了,1月份开始做的,4月份就做出来了。你看这个第5页就是青蒿了,我编了这个本子,然后给“523办公室”送去,我说,我整理的这个,你们可以看看。七个省市嘛,我拿去了8本,一本给“523办公室”,每个地区一本,请大家来发掘。从青蒿中发掘出青蒿素牵涉到几个问题,一个品种问题,菊科蒿属是个很大的属,市场调查品种也不少,1963年和1977年的中国药典上就写了两种品种[4]。还有药用部位问题,梗没有效果。我就想到各种因素,后来就跑到门头沟药库,找头年新进的青蒿药材,经研究还真跟采收季节有关。早期采集的根本就没有青蒿素,因为植物体内根本就还没生物合成这些有效物质,只在花前叶茂时才合用。所以,后来药厂提取青蒿素都用那个季节的青蒿叶子为原料。还有一个提取方法问题。从青蒿中得到具有100%疟原虫抑制率的提取物是很关键的问题。由于其抗疟活性基团是过氧,在青蒿原药材中含有很多其他化学成分,在一些常用的水、乙醇做溶剂提取时,温度超过60℃就破坏了具有抗疟活性的过氧基团。后经我们实验,其实青蒿素本身是很稳定的,用水煮沸半小时或乙醇回流4小时,其抗疟活性稳定不变。在做青蒿素化学结构研究时,为确认羰基的存在,我们做了还原衍生物,得到双氢青蒿素。由于在青蒿素中引进了羟基,方便做更多的衍生物。通过青蒿素构效关系的研究,我们明确了过氧是抗疟活性基团,也认识到双氢青蒿素的抗疟活性更高,安全性也比较好。 经研究开发临床药效提高了10倍,剂量就可以减少到1/10,总剂量480mg就解决问题。完成双氢青蒿素研发后,中药所终于将1973年9月发现的双氢青蒿素于1992年获得新药证书。1981年WHO(世界卫生组织)来中国开会,安排我做青蒿素化学研究的报告。会议对于青蒿素是一片赞扬。后来,一位法国记者问,当时是为了战争,现在的话世界都在用青蒿素,你什么感想?我说,我很高兴,作为一个医药科技人员,工作就是要为人类健康服务的。国家鼓励原始性创新,加强原始性创新。我觉得要谈论的问题恐怕已经不是发明的问题了,就是一个原始性创新出来以后怎么办的问题。那么这个原始性创新不是说的,是要很多的艰苦的工作。要营造好的气氛,让学术不浮躁,科学和学术要实事求是。所以我们就一直在说,能不能以青蒿素做一个解剖,从里面吸取一些有意义的经验和教训。中药领域还有一个问题,有的人不太重视青蒿素,觉得单药单一成分,他们一下子就把它推到西药里去了。21世纪,现代科学是与时俱进的、不断发展的,你拿现在科学的观点、手段再来挖掘中医药的宝库,你就能发现一些更好的东西出来。我想在自己的书上提出这个观点,要大家多做些原始创新。另外,原始创新出来以后,应该有些政策出来保护它们。青蒿素出来多少年了,整体成果是大协作的产物,当年的协作单位和参与者从不同方面做出了各自的贡献。在北医就有一位赵燏黄[5]专门搞本草研究的。我们是学了本草的,有这个基础又系统学了中医药,“西学中”给了我思路,从祖先古代经验来找,经验还是很多的。77年在《科学通报》上发表了青蒿素结构,这是第一篇,我们打的报告给卫生部,经批准后发表的,马上就被CA(注:美国化学文摘)收载了。我们总共从青蒿中分离鉴定了17种化学成分,以及几个混乱品种的20来个化学成分,在81年后陆续发表。黎:这个国家发明奖的证书当时是发了6张还是只是这一张?屠:这个我就不知道了。我们也看到过网络上出现过与我们持有的不一样的国家发明奖证书,不知是什么道理。出现有关青蒿素成果的不同认识也不奇怪,但客观事实不会是多版本的。首先要认定事实,认识才能正确,不然只能浪费时间和精力做无谓的争议。我们需要向前看,国家需要大力促进科学发展,实现科教兴国。科技工作者要有责任感,要努力创新。附注二:有关国家发明奖的争议一、国家发明奖名单与鉴定会名单的差异1978年在高邮举办青蒿素鉴定会,会上确定的主要研究单位为卫生部中医研究院、山东省中西医结合研究院、云南省药物研究所、广州中医学院、四川省中药研究所、江苏省高邮县卫生局六家,其他单位为协作单位,1979年9月18日、20日、21日,国家发明奖评选委员会召开第三次会议,审定发明项目及相应的奖励等级,审查批准了十五项发明创造,并分别评定为二、三、四等发明奖。其中抗疟新药——青蒿素的制造获国家发明奖(二等)。10月15日《人民日报》第四版报道了卫生部中医研究院中药研究所、山东省中医药研究所、云南省药物研究所、中国科学院生物物理所、上海有机化学研究所,以及广州中医学院发明的抗疟新药——青蒿素的制造获得国家二等发明奖。由于对外公开报道的就是这六家,至于其他的协作单位包括之前在鉴定会上确定在主要研究单位中的四川省中药研究所、江苏省高邮县卫生局两个单位完全没有提到。在《迟到的报告》一书出版后,四川省中药研究所曾经参加过“523任务”的研究人员还给科技部成果局写过相应的文件要求“还历史于本来面目”。他们认为:“当时的评奖是不公正的,尤其是在鉴定书上他们作为第5家主研单位,为青蒿研究做了大量的工作,但是当青蒿素研究获得奖励时,四川一线的科技工作者却无一人授奖。在青蒿素获国家发明证书时,他们所的“523”科技人员均不知道,未将他们排入获奖单位并没有征求他们的意见。”[6]为什么1978年的鉴定会上提出的6家研究单位到了1979年获奖的时候变了,《迟到的报告》一书上认为:鉴定会由于是把青蒿简易制剂和青蒿素的研究两个项目作为一个大项,合并进行鉴定的,因此,青蒿素的研究部分排了4个单位;青蒿简易制剂的研究排了2个单位。后来国家科委发明奖评选委员会在评审中,认为青蒿素作为一个新药的发明,确认药物的化学结构必不可少。一个新药若不清楚其化学结构,就不可能确定为新药。因此,提出负担青蒿素化学结构研究的中国科学院生物物理研究所和中国科学院上海有机化学研究所应列入主要研究单位。四川省中药研究所和江苏省高邮县卫生局,研制的主要成果是青蒿生药片,对于青蒿素成果的研究发明,应列入主要协作单位。在当时的发明奖励条例中有规定:第十三条对发明项目如有争议,可向上级机关反映,上级机关应认真调查审理。第十四条各部门和各单位对群众的发明,应当给予鼓励,采取严肃认真和实事求是的科学态度。在贯彻执行奖励制度时,必须加强思想政治工作,提出社会主义大协作精神,反对本位主义、个人主义互不协作等不良倾向。对打击、压制发明和在发明尚弄虚作假,剽窃他人劳动成果的行为,应当批评教育,加以纠正,情节恶劣者,应给以处分,甚至依法惩办。[7]对于发明权的争议和处理:凡国家科委发明评选委员会通过的奖励项目,自公布之日起三个月内,任何人如有意义,均可向国家科委发明评选委员会办公室提出。公布三个月后如无人提出异议,即行授奖,超过三个月提出的异议,一般不予受理。凡国家科委发明评选委员会通过奖励的项目,自公布之日起三个月内,如有争议,一般由发明申报部门负责处理。为及时解决争议问题。正以任何一方,均应按照争议处理部门的要求,及时地如实的提供有关争议的补充材料和旁证文件。如在六个月内不作答复,即判为弃权,由争议处理部门裁决。[8]四川省中药研究所并没有在1979年公布结果的时候依据发明奖条例上的规定向国家科委提出意见而是到2005年才提出意见。此时国家科委并没有给出明确的解释。对此,笔者认为,这是文革后国家发明奖首次评选,在没有一个十分明确、细致的评审标准的情况下,国家发明奖评选委员会在评审时认为:作为一个新药的发明,确认药物的化学结构必不可少;一个新药若不清楚其化学结构,就不可能确定为新药。这种依据,也符合当时的历史事实。不过,无论是国家发明奖评奖委员会还是“523办公室”在1979年并未对四川和江苏这两家单位排除在获奖名单之外做出公开说明或者说并未告知他们,这说明当时在荣誉分配的问题上处理的不是很好,也从一个侧面反映了“523任务”在快要结束时的收尾工作做的不够完善。虽然后来《迟到的报告》一书中对四川省中药研究所和江苏省高邮县卫生局未能出现在获奖名单名单上做出了解释,也肯定了四川省中药研究所在“523任务”开展过程中的大量工作,但是这样的说明似乎有点晚而且权威性并不够。二、国家发明奖证书编号引发的误解我们在最初拥有的资料中发现《迟到的报告》一书与《青蒿素研究》一书中所使用的青蒿素的发明证书主要内容相同,只有其中的证书号码与排版方式略有不同,《迟到的报告》一书中编号为A00011,而《青蒿素研究》一书中的编号为A00612。为此,笔者在访谈不同的老科学家时也提到这个疑问,但是他们都不清楚这是为什么,有人说可能有真有假,也有人说是所里面弄丢了后来补发的,但是具体什么时候补发的就不知道。随着我们深入了解与资料搜集的完善,终于使该疑问有了一个比较清晰的答案。目前收集到的关于青蒿素的国家发明奖证书有几个不同编号的证书,第一个是中医研究院目前保存的编号为00011的发明证书[9],其他为保存在其余获奖单位的证书,编号为别为A00609(山东省中医药研究院)[10]、A00610(云南省药物所)[11]、A00612(中国科学院上海有机化学研究所)[12]。这几个单位拥有的证书原件如下:由于条件所限,我们未能获得全部的证书原件,根据证书的编排判断还有两张编号不同的证书,其中之一为中国科学院生物物理所的,证书编号应该为A00611,另一张为广州中医学院的证书编号为A00613,这些证书是连号的。根据发明奖规定的奖金分配原则:集体发明(包括协作单位),所得奖金按照发明者贡献大小,合理分配。几个单位共同完成的发明,其奖金由申报部门负责进行合理分配。针对青蒿素是卫生部作为申报部门申报的,因此奖金分配也由卫生部进行。有关发明奖证书、奖章以及奖金的分配情况,有如下记载[13]:图5青蒿素发明奖奖章(屠呦呦提供)图6卫生部科技局下发青蒿素发明奖奖金分配的通知图7中医研究院中药研究所向其他单位写的通知信函根据卫生部当时的文件指明:国家科委颁发的青蒿素发明证书(见中医研究院的发明证书)、奖章(只有一份,如上图为复制件)[14],建议由中医研究院照相复制给其他五个单位,原件由中医研究院中药研究所保存。根据中医研究院的信函以及相关单位的档案显示,当时中医研究院中药研究所将影印件给了其他单位。因此,当时虽然是六家单位为发明单位,但是拥有证书和奖章原件的只有中医研究院中药研究所一家。其实卫生部的这份通知为后来的多张证书的出现既提出的异议又为后面的争议埋下的伏笔。既然当时只有一张证书,现在为什么出现了6张,而且编号相差很多,但是后5张却是连号?为此笔者询问了原国家科委的一位对此有所了解的工作人员,据他所述:“青蒿素是1979年获得的国家发明二等奖,但是后来因为获奖单位和发明人的排名问题一直存有争议,后经较长时间的调解,达成较为一致的排名顺序(但还是有些单位和人员不服),重新打印了获奖证书。这就是二个证书的来由,应该是以第二张证书为准。”[15]其实前后的证书,内容都是一样,不同的只是证书编号和排版方式。因此,经过上述的说明,笔者认为,两种版本的4张证书以及还可能出现的两张不同编号的证书从根本上来说都是真的。根据科委工作人员所说的对排名问题一直存在争议,说明这个问题一直未得到很好的解决,就算是重新打印了获奖证书之后,很多研究人员并不知情,有关争议还是存在。注释:(上下滑动可浏览)[1]醚中干,指的是在研究植物药有效成分时用乙醚提取,先将植物中的混合成分按其极性大小粗略地分开,乙醚提取液中含有的主要是极性小的脂溶性混合物,再用不同强度的碱溶液(碳酸氢钠、碳酸钠或氢氧化钠溶液)依次提取,将得到强度不同的酸性成分;经过碱提取的残余液可以再用酸溶液(盐酸或硫酸溶液)提取,得到碱性成分(一般是生物碱或其它碱性成分),乙醚残余液中剩下的就只有中性混合成分。这是一种经典的系统分离方法。屠呦呦在分离过程中使用的是2%的氢氧化钠溶液提取,并省略了用酸溶液提取这一步,得到的乙醚中性部分(简称“醚中干”)[2]屠呦呦与余亚纲同为中医研究院西医离职学习中医班第三期学生,学习时间为1959年10月至1962年2月。[3]蒲辅周(1888-1975),四川省梓潼县人。现代中医学家,长期从事中医临床、教学和科研工作,精于内、妇、儿科,尤擅治热病。伤寒、温病学说熔于一炉,经方、时方合宜而施。1955-1975年历任中医研究院广安门医院内科医师、内科主任,中医研究院副院长等职务。曾任全国政协第三、四届委员。[4]《中华人民共和国药典》(以下简称《药典》)1977 版中药青蒿的原植物为黄花蒿(Artemisia annua L.)或青蒿(ArtemisiaapiaceaHance)二种植物,在解释性状时也分为黄花蒿和青蒿两种。不过从《药典》1985版开始,中药青蒿的原植物只有黄花蒿(Artemisia annua L.)一种植物。[5]赵燏黄(1883-1960),江苏省武进县,本草学家和中国生药学先驱者。1909年毕业于日本东京药学专门学校。他毕生致力于本草学和生药学的教学和科学研究,应用现代科学方法整理本草,研究中药,考订本草药品名实,为编修中国新本草做了大量开拓性工作,在澄清中药品种混乱方面做出了贡献,并为中国培育了几代药学科技人才。先后担任过任上海国立中央研究院化学研究所研究员,北京大学医学院药学系教授,中央卫生研究院中医(祖国医学)研究所生药室顾问、中国药学会北平分会副理事长、中医研究院中药研究所研究员等职务。[6]齐尚斌、万尧徳.还历史于本来面目——青蒿素研制中鲜为人知的故事.2005-11-1[7]中华人民共和国发明奖励条例(1978年12月28日国务院发布)[8]中华人民共和国发明奖励条例(1978年12月28日国务院发布,1984年4月25日国务院修订)[9]屠呦呦提供[10]山东省中医药研究院档案处提供[11]罗泽渊提供[12]中国科学院上海有机化学研究所档案处提供[13]中国科学院档案馆.青蒿素的结构和反应.档案号D144-357[14]屠呦呦提供[15]有关获奖证书的回忆.北京大学医学史研究中心.523任务相关档案20101119[16]https://cn.nytimes.com/china/20151010/c10sino-tuyouyou/亲爱的读者们,不星标《知识分子》公众号,会错过每日科学新知!星标《知识分子》,紧跟前沿科学,一起探索科学的奥秘吧! 请戳上图卡片添加星标关注《知识分子》视频号get更多有趣、有料的科普内容END
临床研究
2025-04-29
·漫游药化
在读博五年多,历经一次又一次无功而返的科研旅程之后,来自中国科学院上海有机化学研究所的阮林新博士终于以一篇《科学》文章证明了自己。在科研的江湖上,结果为王,好在阮林新笑到了最后。国家自然科学基金委员会在其官网上以“我国学者在(动态)动力学调控不对称酮加成研究方面取得进展”为题关注了上述文章,文章的题目是《酮的(动态)动力学不对称芳基化和烯基化》。专家们认可这项成果,因为其利用自主研发的手性卡宾-镍催化剂,通过手性η2羰基活化新模式,解决了传统不对称羰基加成活化模式单一,反应活性低的问题,实现了非活化酮的动态动力学不对称芳基化和烯基化反应。如何理解?阮林新的解读是,这项研究是为了一款近40年来出现的治疗肺结核的重要药物而进行的;在理论上,该成果可以实现原料的高效利用,提高生产效率,大幅降低百姓的用药成本。从基础研究到工业应用,中间要经历“爬雪山、过草地”基础研究是整个科学体系的源头,是解决所有技术问题的总机关。但从基础研究到应用,中间隔了二万五千里长征路,“爬雪山、过草地”在所难免。在阮林新看来,从目前他做的基础研究走到工业上的放大生产,最大的难点就是解决放大反应过程中所需要的关键性金属催化所导致的金属残留问题,另一个难点就是产物的立体选择性还不够高。“如何改进当下的催化剂来提高反应选择性和反应效率,同时降低金属用量,去除金属残余,是走向产业化最艰辛的一步。”阮林新说。药物研发是人类发展过程中跟疾病斗争最有效的手段。一个新药,大概要经历十年时间才能从想象走向成药的过程。为什么要花这么长时间?中国科学院院士、中国科学院副院长、中国科学院大学校长周琪曾这样回答,第一,要知道治疗一个疾病主要针对的靶点是什么,就是有的放矢,要知道治什么东西。第二,当确定了药物靶点,要选到所有的候选化合物和候选药物,不是每一种药都可以成药的,同样还要经过药物的各种成药的相关试验。对于自己的研究成果要经历十年乃至数个十年才能面世,阮林新淡然却认真地对待这一过程。“人生短暂却也漫长,做好一件事便是成功。”阮林新。但他也做好了另一种准备——失败。因为很多科学家在持续了几十年的研发之后,只得到一个结果:此路不通。对于失败,阮林新有刻骨铭心的记忆。从2017年在有机化学研究所硕博连读到2023年博士毕业的5年半时间里,他曾经历过一次又一次的失败。记忆中,身边的博士同学在二年级或三年级都已经在发表文章,而阮林新却在临近毕业的最后半年里还没有成果,甚至直到答辩前夕,他的科研成果都没被接受。他不知道自己的成果何时能成功,甚至能不能成功,更重要的是,作为一个全新的体系,是否能被更多国际专家认可,一切都是未知数。只有在不断失败中坚持目标,才能拿到打开科研大门的钥匙“更大的挑战,就是在心理上能不能承受日复一日的失败,是否能在不断失败里仍然坚持科研目标,坚持自己的初心是非常难的。”好在苦心人,天不负,他最终拿到了打开科研大门的钥匙。阮林新总结:“只要坚持到最后,往往会有峰回路转的可能性,说不定就有奇迹发生。在这期间,心态很重要,不能放弃,不能自暴自弃。”记者了解到,阮林新是从国家级贫困县走出来的学生,自小学起父母就在外打工。在他家周围只有一所小学,十里八村的孩子都在这一所小学读书,冬冷夏热,教学设施和交通都很艰苦。读中学时,他住在二三十人的大通铺宿舍。为了节省开支,每周一去学校上学时,他会带够一周的菜量。天气热的时候,便只能带咸菜才能放得住。熬到周五实在没有东西吃,用热水冲一碗速食紫菜汤就一碗米饭吃饱。考上省重点高中,最让他开心的是,可以拥有一张单独的床,而不用睡通铺。在艰苦条件下,家人支持给了他莫大的力量。哪怕在他临近博士毕业时还没有出成果,甚至可能面对延后毕业一年或更久,家人总是打电话劝慰:“你不要有任何压力,大不了回家也没有关系的。”从国家级贫困县走到大城市,成为人人尊崇的科研者,阮林新有了更多使命感。他告诫自己,决不允许半途而废,“困难肯定是有,但是也会有一些美好的回忆,支撑我一步步走过来。”如今,他拿到了2023年度京博科技奖-化学化工与材料京博优秀博士奖金奖,对未来有了新的规划,“我需要通过自己的努力去做一些有用的发现,造福社会,造福人民。哪怕以后的学生在查找这方面的材料文献时,能找到我的名字,这也是一件非常自豪的事情。” (受访者供图)来源:科技日报 记者 王延斌,原题:《阮林新:在枯燥和失败中耐住寂寞,才能抓住一丝成功的机会》
2025-04-17
·智药邦
帕金森病(Parkinson’s Disease,PD),是一种影响运动和认知功能的神经退行性疾病,也是世界上第二常见的神经退行性疾病,仅次于阿尔茨海默病(AD),影响着约 1%-2%的65岁及以上老人。随着全球人口老龄化,帕金森病患病率还将大幅增加。病理性α-突触核蛋白(α-Syn)在不同脑区聚集介导了帕金森病的疾病进展。聚集的α-Syn蛋白形成纤维,以朊病毒样(Prion-like)方式发挥作用,在大脑的不同区域传播α-Syn病理,然而,其在神经元之间传递的分子机制,目前仍不清楚。之前的研究显示,α-Syn通过不同受体的内吞作用被内化,然而,其在神经元上的特异性受体仍有待完全鉴定。郁金泰团队曾于2020年10月在Science Advances期刊发表论文,首次发现了FAM171A2基因的单核苷酸多态性(rs708384)与帕金森病等神经退行性疾病的风险增加有关。帕金森病患者的脑脊液(CSF)中FAM171A2蛋白水平增加,并且与脑内α-Syn高聚集的指征相关。2025年2月20日,复旦大学附属华山医院郁金泰教授团队联合复旦大学脑科学转化研究院袁鹏教授及中国科学院上海有机化学研究所刘聪教授(博士后吴凯敏为第一作者),在国际顶尖学术期刊Science上发表了题为:Neuronal FAM171A2 mediates α-synuclein fibril uptake and drives Parkinson’s disease的研究论文。该研究首次发现了帕金森病(PD)的全新治疗靶点——FAM171A2,并利用基于人工智能的蛋白质结构预测和虚拟筛选技术,成功找到了具有潜在治疗作用的小分子化合物。这项研究发现的全新治疗靶点和前在治疗药物有望从疾病早期对帕金森病进行干预,延缓疾病进展,结合现有的对症治疗手段,有望实现帕金森病病因治疗与症状缓解的双重突破。在这项最新研究中,研究团队通过对超过100万样本的大规模全基因组关联分析(GWAS),发现FAM171A2基因的5个突变(例如rs850738、rs708384)与帕金森病风险显著相关。帕金森病患者的脑脊液(CSF)中FAM171A2蛋白水平升高,且与α-Syn病理标志物呈负相关,提示其参与了α-Syn的异常聚集。进一步实验验证显示,过表达FAM171A2会促进α-Syn纤维的被神经元内吞,加剧病理扩散(例如向黑质、纹状体的扩散)和运动功能障碍。而敲低FAM171A2能够减少α-Syn纤维的内吞,保护多巴胺能神经元,从而改善小鼠的运动表现。研究团队在小鼠神经元中特异性敲除FAM171A2,进一步验证了神经元来源的FAM171A2在α-Syn病理传播中是必要的。从机制上来说,FAM171A2蛋白的胞外结构域1通过静电力与α-Syn的C端相互作用,且高度选择性地结合病理性α-Syn(纤维形式),其对纤维形式α-Syn的选择性结合是单体形式α-Syn的超1000倍。也就是说,位于神经元上的FAM171A2蛋白能够高度选择性的结合病理性α-Syn,使其通过内吞作用进入神经元内,诱导神经元内的正常的单体形式的α-Syn发生错误折叠,转变为病理性α-Syn,进而诱导神经元死亡以及在神经元之间传播。接下来,研究团队使用AlphaFold-Multitimer预测了FAM171A2-α-Syn复合物结构,并利用虚拟筛选技术,从7173种化合物中筛选出了一种小分子药物——Bemcentinib,其能够竞争性结合α-Syn的C端,从而阻断FAM171A2与病理性α-Syn结合,抑制FAM171A2介导的神经元对病理性α-Syn的内吞。总的来说,该研究首次阐明了FAM171A2作为病理性α-Syn的神经元受体,揭示了其驱动帕金森病病理传播的分子机制,并成功筛选到了潜在抑制剂,为帕金森病的精准治疗提供了新方向。未来研究可聚焦于靶向FAM171A2的药物开发及多维度验证其临床价值。相关阅读:2024年11月22日,复旦大学附属华山医院郁金泰、毛颖团队与复旦大学类脑智能科学与技术研究院程炜、冯建峰团队合作,在国际顶尖学术期刊Cell在线发表了题为:Atlas of the plasma proteome in health and disease in 53,026 adults的研究论文。这项研究全面绘制了人类健康与疾病蛋白质组图谱,并结合人工智能大数据分析方法,构建了疾病诊断预测模型以及发现了26个药物治疗新靶点,为精准医学的实施提供了重要科学依据。该研究于2025年1月9日,登上了Cell期刊封面,该封面描绘了一个由蛋白质构成的人类,正在借助人工智能(由芯片和二进制代码代表)的帮助来检查自己的健康状况(通过心电图监测界面来体现)。参考资料:1.https://www.science.org/doi/10.1126/sciadv.abb30632.https://www.science.org/doi/10.1126/science.adp36453.https://www.cell.com/cell/fulltext/S0092-8674(24)01268-6--------- End ---------感兴趣的读者,可以添加小邦微信加入读者实名讨论微信群。添加时请主动注明姓名-企业-职位/岗位或姓名-学校-职务/研究方向。
申请上市临床研究
100 项与 中国科学院上海有机化学研究所 相关的药物交易
登录后查看更多信息
100 项与 中国科学院上海有机化学研究所 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2026年07月28日管线快照
管线布局中药物为当前组织机构及其子机构作为药物机构进行统计,早期临床1期并入临床1期,临床1/2期并入临床2期,临床2/3期并入临床3期
药物发现
1
8
临床前
其他
4
登录后查看更多信息
当前项目
登录后查看更多信息
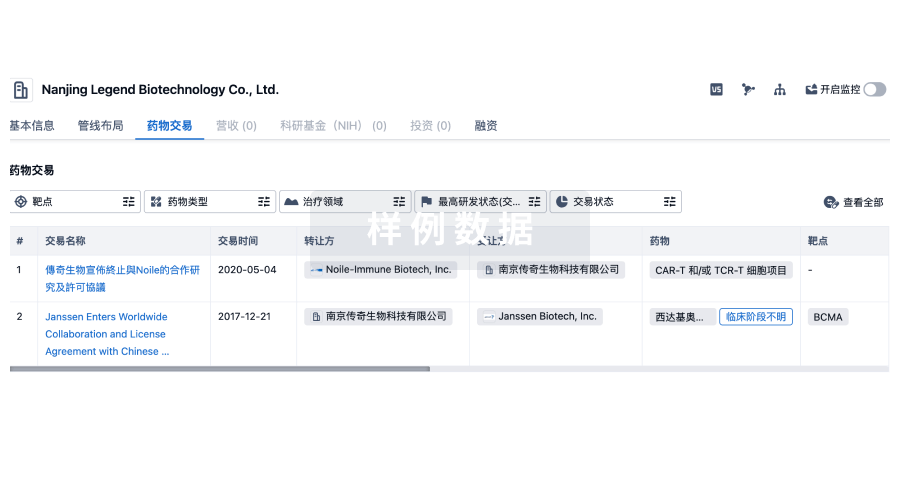
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
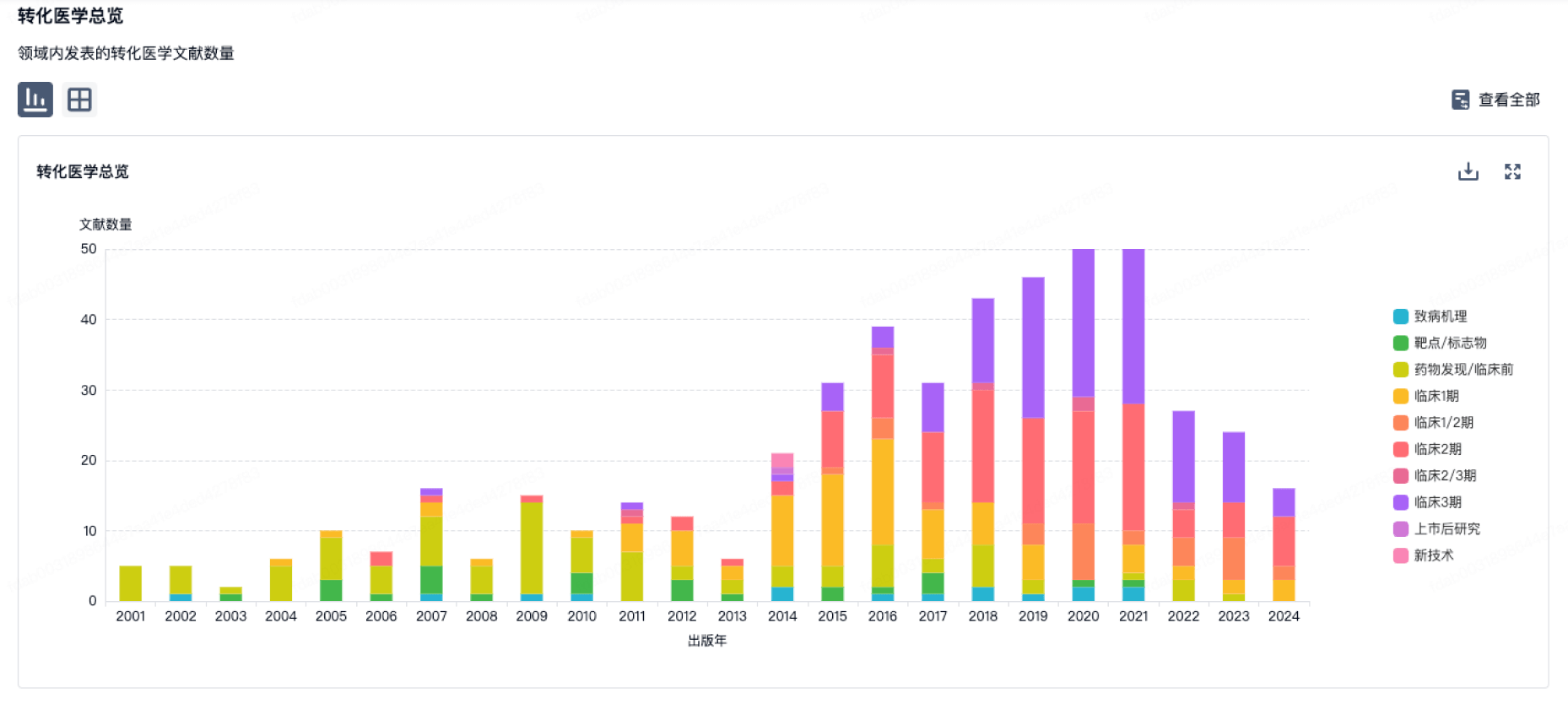
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
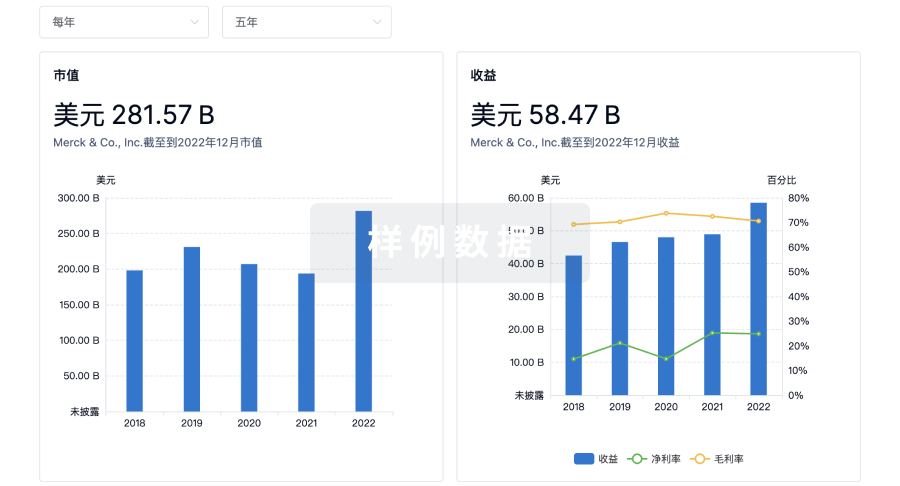
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
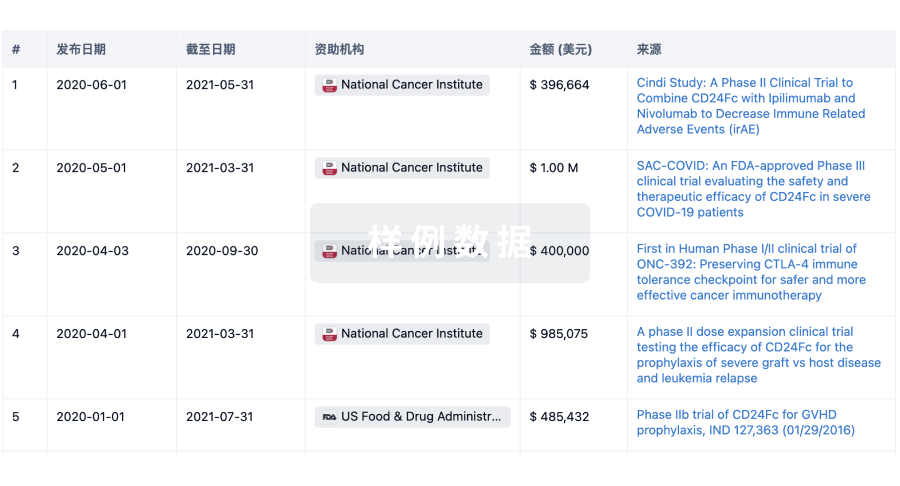
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
芽仔
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用