预约演示
更新于:2026-05-10
Sinovac Biotech Ltd.
更新于:2026-05-10
概览
标签
感染
消化系统疾病
眼部疾病
预防性疫苗
减毒活疫苗
灭活疫苗
疾病领域得分
一眼洞穿机构专注的疾病领域

技术平台
公司药物应用最多的技术
靶点
公司最常开发的靶点
关联
靶点 |
作用机制 |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症 |
最高研发阶段 |
首次获批国家/地区 |
首次获批日期 |
靶点 |
作用机制 |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症 |
最高研发阶段 |
首次获批国家/地区 |
首次获批日期 |
靶点 |
作用机制 |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症 |
最高研发阶段 |
首次获批国家/地区 |
首次获批日期 |
NCT07473427
Immunogenicity and Safety of Varicella Vaccine, Live in Healthy Vietnamese Children Aged 1~12 Years: A Single-armed Bridging Clinical Trial
NCT07505810
A Randomized, Double-Blind, Active Comparator-Controlled Phase Ⅲ Trial to Evaluate the Immunogenicity and Safety of the Group ACYW135 Meningococcal Conjugate Vaccine in Population Aged 3 to 5 Months
NCT07505888
A Randomized, Double-Blind, Active Comparator-Controlled Phase Ⅲ Trial to Evaluate the Immunogenicity and Safety of Group ACYW135 Meningococcal Conjugate Vaccine in Population Aged 2-6 Years
100 项与 科兴控股生物技术有限公司 相关的临床结果
登录后查看更多信息
登录后查看更多信息
2026-12-31Human Vaccines & Immunotherapeutics
Standardized neutralizing antibody assay for Coxsackievirus A10 in clinical trial: A framework for multivalent hand, foot, and mouth disease vaccine evaluation
Article
作者: Xie, Zhongping ; Zhang, Zhongyang ; Wang, Wenhui ; Li, Yajing ; Chen, Xiaoqi ; Ma, Xiao ; Wang, Ying ; He, Qian ; Liu, Qingwei ; Gao, Junying ; Liu, Mingchen ; Han, Jinle ; Wang, Qian ; Gao, Fan ; Guo, Jing ; Li, Lu ; Liang, Zhenglun ; Mao, Qunying ; Gu, Meirong ; Wu, Xing
Enteroviruses (EVs) are the major etiological agents of hand, foot, and mouth disease (HFMD), which could lead to severe complications and even fatalities. Owing to the alternating or co-circulation of multiple EV serotypes and the lack of cross-protection, global institutions are pursuing the development of multivalent HFMD vaccines. Notably, the advancement of innovative multivalent HFMD vaccines into the clinical stage highlights the urgent need for a standardized platform to assess immunogenicity. In accordance with international regulatory guidelines, a standardized neutralizing antibody (NtAb) assay for CV-A10 is established through rigorous selection and systematic optimization of key assay components, representing the first such assay tailored for clinical immunogenicity evaluation of CV-A10-based vaccines. By integrating the 1st Chinese National Standard (NS) for CV-A10 NtAb and a standard detection virus, this assay exhibits enhanced detection sensitivity and significant reduction in the inter-laboratory geometric coefficient of variation (GCV), thereby providing a robust, reliable, and transferable platform. This assay facilitates accurate evaluation of multivalent HFMD vaccine efficacy, improves comparability across different vaccine products and provides genuine efficacy in real-world, serving as a valuable reference for global regulatory authorities.
2025-12-31Human Vaccines & Immunotherapeutics
Lot-to-lot consistency, immunogenicity, and safety of a 23-valent pneumococcal polysaccharide vaccine (PPV23) in healthy adults aged 40–65 years: A randomized, double-blind, controlled, phase IV clinical trial
Article
作者: Geng, Jiaxu ; Zhao, Haiyan ; Chen, Zhuozhuo ; Zhou, Siliang ; Zhang, Yunong ; Shi, Gang ; Xu, Li ; Li, Gaimei ; Zhao, Zhifeng ; Li, Yun ; Wang, Lianhao ; Li, Guohua ; Zhao, Weimin ; Jiao, Peng ; Wang, Bin ; Li, Jidong
This randomized, double-blind, phase IV study (NCT05731115) assessed the lot-to-lot consistency, immunogenicity, and safety of three manufacturing lots of a 23-valent pneumococcal polysaccharide vaccine (PPV23) in 1800 healthy Chinese adults aged 40-65 y. The participants were enrolled and randomized into three groups at a 1:1:1 ratio, with each group receiving one dose of PPV23 from one of three manufacturing lots. Blood samples were collected pre-vaccination and at 30 d post-vaccination for serotype-specific IgG antibody testing. The primary objective was to demonstrate lot-to-lot consistency via equivalence evaluation in 40-65 y old Chinese adults, assessed by geometric mean concentration (GMC) of IgG antibodies at 30 d post-vaccination. Safety data were collected for up to 30 d after vaccination. The results showed that GMC ratio 95% confidence intervals (CIs) in all 23 serotypes were within the predefined equivalence criteria of (0.5, 2). Although the incidence of adverse reactions (AR) may have varied among the three lots, the clinical impact is likely minimal, given that the vast majority (93.2%) were mild in severity. No serious adverse events (SAEs) related to vaccination were reported during the study. This study confirmed that the three manufacturing lots of PPV23 (Sinovac Biotech Co. Ltd.) demonstrated manufacturing consistency and can elicit satisfactory immune responses for all 23 serotypes and no safety concerns were identified.Trial registration: ClinicalTrial.gov identifier: NCT05731115.
2025-12-31Emerging Microbes & Infections
Structural basis for broad neutralization of rabies virus by an antibody cocktail SYN023
Article
作者: Jinyue Wang ; Xiangxi Wang ; Panayampalli Subbian Satheshkumar ; Michael Niezgoda ; Chuziyue Zhang ; Jiagui Qu ; Eric Tsao ; Christine Fehlner-Gardiner ; Ruihong Chen ; Lei Cao
2026-05-10
·今日头条
2026-05-09
100 项与 科兴控股生物技术有限公司 相关的药物交易
登录后查看更多信息
100 项与 科兴控股生物技术有限公司 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2026年07月21日管线快照
管线布局中药物为当前组织机构及其子机构作为药物机构进行统计,早期临床1期并入临床1期,临床1/2期并入临床2期,临床2/3期并入临床3期
药物发现
1
7
临床前
临床申请
5
2
临床申请批准
临床1期
6
5
临床2期
临床3期
4
1
申请上市
批准上市
14
6
其他
登录后查看更多信息
当前项目
登录后查看更多信息
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
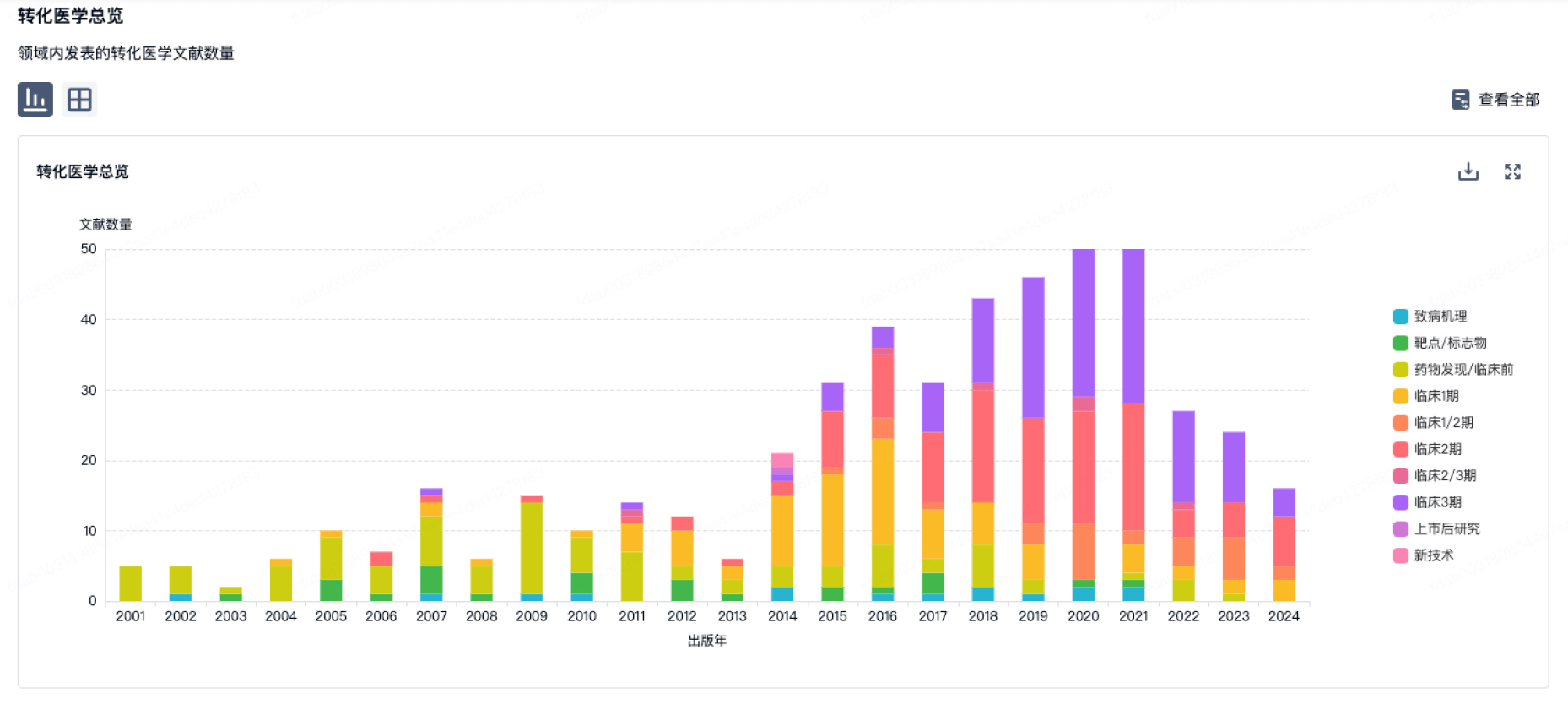
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
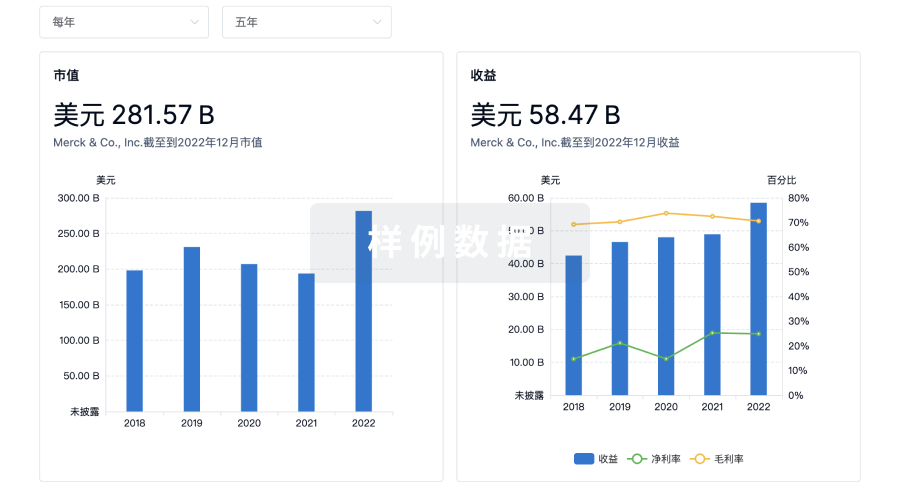
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
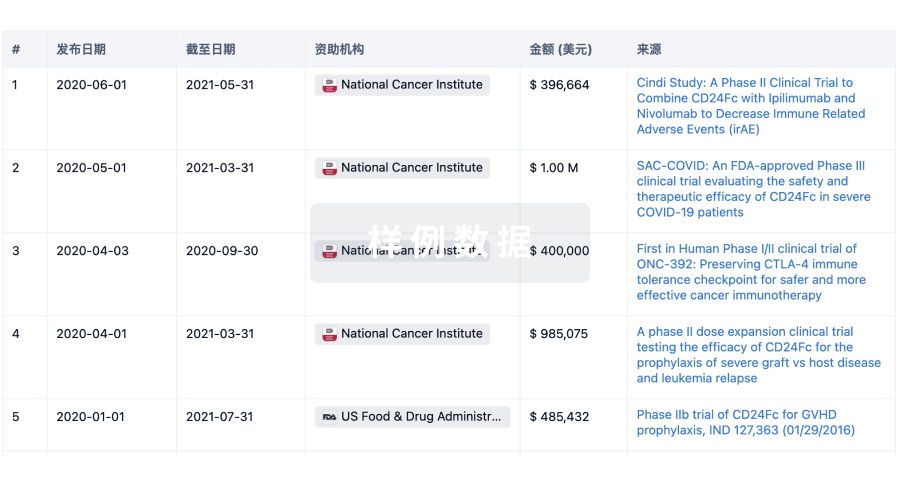
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
芽仔
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用