预约演示
更新于:2026-06-06
Ascorbic Acid
维生素C
更新于:2026-06-06
概要
基本信息
原研机构 |
权益机构- |
最高研发阶段批准上市 |
首次获批日期 日本 (1978-01-31), |
最高研发阶段(中国)批准上市 |
特殊审评孤儿药 (美国)、孤儿药 (欧盟) |


登录后查看时间轴
结构/序列
分子式C6H8O6 |
InChIKeyCIWBSHSKHKDKBQ-JLAZNSOCSA-N |
CAS号50-81-7 |
关联
1,035
项与 维生素C 相关的临床试验NCT06710899
Does Vitamin C Improve the Efficacy of Methenamine Hippurate (Hiprex®) in Prophylaxis of Recurrent Urinary Tract Infections? A Proof of Concept Study

NCT07612345
Safety and Tolerability of Supratherapeutic Vitamin C Supplementation in Class A Glucose-6-Phosphate Dehydrogenase (G6PD A) Deficiency and Pyruvate Kinase Deficiency (PKD)

ChiCTR2600125975
The Effect of Intravenous Vitamin C on Intraoperative Tourniquet-Related Hypertension and Postoperative Pain in Patients Undergoing Lower Limb Fracture Surgery
100 项与 维生素C 相关的临床结果
登录后查看更多信息
100 项与 维生素C 相关的转化医学
登录后查看更多信息
100 项与 维生素C 相关的专利(医药)
登录后查看更多信息
122,051
项与 维生素C 相关的文献(医药)2026-12-31JOURNAL OF DERMATOLOGICAL TREATMENT
Clinical outcomes of intense pulsed light combined with non-crosslinked sodium hyaluronate, tranexamic acid, and vitamin C mesotherapy for facial photoaging: a retrospective study
Article
作者: Li, Fei ; Xu, Jialu ; Qiu, Houhuang ; Wu, Xixin ; Wu, Yutong ; Xu, Tianhua ; Wang, Fangfang ; Chen, Xi ; Zhong, Ping ; Lin, Qiusheng
BACKGROUND:
Facial photoaging involves structural and functional deterioration across multiple skin layers. Single-modality treatments rarely address pigmentary, vascular, and dermal matrix changes concurrently. Intense pulsed light (IPL) is widely used for the treatment of dyschromia and vascular lesions. Mesotherapy incorporating non-crosslinked sodium hyaluronate (NCSH), tranexamic acid (TXA), and vitamin C (VC) has been introduced to improve skin hydration and related dermal parameters. The present study assessed the efficacy and safety of combining these modalities for facial rejuvenation.
METHODS:
Eighty-four patients underwent three sessions of IPL with mesotherapy. Standardized VISIA imaging was conducted before each treatment (T0, T1, T2) and at 1-2 months (T3) and 3-6 months (T4) post-treatment. Efficacy was assessed using the Modified Fitzpatrick Wrinkle Scale (MFWS) and Global Aesthetic Improvement Scale (GAIS); adverse events and satisfaction were recorded.
RESULTS:
All six VISIA parameters and MFWS scores improved significantly (p __-mml:math xmlns:mml="http://www.w3.org/1998/Math/MathML"-__<
CONCLUSIONS:
IPL combined with NCSH/TXA/VC mesotherapy provided safe, effective, and well-tolerated improvement in facial photoaging, representing a promising multimodal rejuvenation approach.
2026-12-31Journal of the International Society of Sports Nutrition
Micronutrient intake and predictors of adequacy in female collegiate dancers consuming low-energy diets
Article
作者: Jordan, Aydan ; Sanders, David J. ; Saenz, Catherine ; Frost, Ann ; Stroh, Eli ; Vatne, Emaly ; Nickerson, Brett S. ; Calamoneri, Tanya ; Brooks, Samantha
BACKGROUND:
Collegiate dancers face high training loads, aesthetic pressures, and limited access to nutritional resources, elevating their risk for low energy intake and micronutrient insufficiency. Although inadequate micronutrient intake has been documented, prior studies are largely limited to single sites, ballet-focused programs, or short assessment periods, making findings difficult to generalize and leaving long-term habitual intake across diverse collegiate dance styles poorly understood. Given the elevated risk for low energy intake and nutrient inadequacy in this population, further investigation is warranted. This study aimed to evaluate habitual micronutrient intake in female collegiate dancers relative to the Recommended Dietary Allowance (RDA) and sports nutrition guidelines, and to identify dietary predictors of micronutrient adequacy.
METHODS:
Thirty female collegiate dancers from three U.S. university dance programs (mean age = 20.4 years; BMI = 21.1 kg/m²) completed the Diet History Questionnaire III. Total energy and micronutrient intake from food and supplements were quantified for calcium, iron, folate, omega‑3 fatty acids, and vitamins B12, C, and D. Adequacy relative to the RDA/Adequate Intake (AI) was determined for each nutrient. Fisher's exact tests compared adequacy between supplement users and non‑users. Logistic regression models evaluated predictors of meeting the RDA/AI, including total energy intake (per 100 kcal), supplement use, and relevant food‑group intake.
RESULTS:
Most dancers consumed below the RDA/AI for calcium (20/30; 67%), iron (24/30; 80%), vitamin D (20/29; 69%), omega‑3 fatty acids (19/30; 63%), and folate (17/29; 59%). Conversely, a majority met the RDA for vitamin B12 (17/26; 65%) and vitamin C (20/30; 67%). When compared with sports nutrition recommendations, an even greater proportion fell short for calcium (29/30), omega‑3 fatty acids (30/30), vitamin D (22/29), and vitamin C (14/30). Supplement use was significantly associated with meeting the RDA for iron (75% vs. 11.5%; p = 0.018; OR = 19.14), folate (87.5% vs. 23.8%; p = 0.003; OR = 19.61), vitamin B12 (100% vs. 47.1%; p = 0.009; OR = Inf), vitamin C (94.1% vs. 46.2%; p = 0.009; OR = 16.68), and vitamin D (56.3% vs. 0%; p = 0.001; OR = Inf). Total energy intake significantly predicted adequacy for calcium (p = 0.020; OR = 1.27), iron (p = 0.0002; OR = 1.69), omega‑3 fatty acids (p = 0.007; OR = 1.27), folate (p = 0.004; OR = 1.57), and vitamin B12 (p = 0.004; OR = 1.39), but not vitamin C or D. Food‑group intake showed limited predictive value; only red/orange vegetable intake predicted vitamin C adequacy.
CONCLUSIONS:
Inadequate micronutrient intake was highly prevalent among collegiate dancers, particularly for calcium, iron, vitamin D, omega‑3 fatty acids, and folate. Supplement use improved adequacy for several nutrients, yet observed intake distributions suggest supplementation often compensates for, but not necessarily corrects, underlying dietary shortfalls. Total energy intake emerged as a consistent predictor of adequacy across multiple nutrients, underscoring the central role of sufficient energy intake in supporting micronutrient exposure. Even dancers meeting the RDA frequently fell below athlete‑specific recommendations, highlighting the need for targeted strategies that address both energy intake and micronutrient support within collegiate dance programs.
2026-12-31Artificial Cells Nanomedicine and Biotechnology
Formulation and characterization of ascorbic acid-based zinc oxide nanoparticles for assessing antidiabetic and neuroprotective effects in STZ-induced diabetic rats
Article
作者: D. R, Bharathi ; Pasha, Ismail ; Ahmed, Syed Sagheer ; Alhamhoom, Yahya ; Alamri, Ali ; Rahamathulla, Mohamed ; Ahmed, Mohammed Muqtader ; Ali, Mohammad ; M. B, Sindu
Ascorbic acid (AA) has strong antioxidant and anti-diabetic activities; nevertheless, its therapeutic value is restricted due to rapid degradation, low-bioavailability and short systemic retention. To address these constraints, this study describes the development of a green-synthesized zinc oxide nanoparticle (ZnO NP)-based delivery system functionalized with AA (ZnO-AA NPs) and assesses its antidiabetic and neuroprotective effects. AA-assisted precipitation method was used to develop ZnO-AA NPs, which were then characterized by dynamic light scattering (DLS), zeta potential (ZP), scanning electron microscopy (SEM), FTIR spectroscopy and X-ray diffraction (XRD). The nanoparticles had a homogeneous size (∼38 nm), good colloidal stability and a crystalline wurtzite structure. In vitro, ZnO-AA NPs showed dose-dependent inhibition of α-amylase and α-glucosidase. In vivo, oral ZnO-AA NPs (25 and 50 mg/kg) significantly improved glycaemic control, lipid profile and antioxidant enzyme levels compared to free AA (*p* < 0.01, *p* < 0.001; one-way/two-way ANOVA). ZnO-AA NPs dramatically reduced diabetes-related cognitive impairment, resulting in improved performance in the Morris water maze (MWM) and open field tests (OFTs) (*p* < 0.001), decreased hippocampal acetylcholinesterase (AChE) activity and preserved neuronal architecture. Overall, the ZnO-AA nanoformulation represents a multifunctional nanomedicine technique that improves the therapeutic efficacy of AA and has considerable promise for the treatment of diabetes and its associated neurodegenerative consequences.
1,687
项与 维生素C 相关的新闻(医药)2026-06-04
·今日头条
100 项与 维生素C 相关的药物交易
登录后查看更多信息
研发状态
批准上市
10 条最早获批的记录, 后查看更多信息
登录
| 适应症 | 国家/地区 | 公司 | 日期 |
|---|---|---|---|
| 铁过剩 | 中国 | 1981-01-01 | |
| 高铁血红蛋白血症 | 中国 | 1981-01-01 | |
| 坏血病 | 中国 | 1981-01-01 | |
| 抗坏血酸缺乏 | 日本 | 1978-01-31 |
未上市
10 条进展最快的记录, 后查看更多信息
登录
| 适应症 | 最高研发状态 | 国家/地区 | 公司 | 日期 |
|---|---|---|---|---|
| 营养和代谢疾病 | 临床3期 | 美国 | 2018-02-09 | |
| 脓毒性休克 | 临床3期 | 美国 | 2018-02-09 | |
| 结直肠癌 | 临床3期 | 比利时 | 2014-10-01 | |
| 结直肠癌 | 临床3期 | 法国 | 2014-10-01 | |
| 结直肠癌 | 临床3期 | 德国 | 2014-10-01 | |
| 结直肠癌 | 临床3期 | 意大利 | 2014-10-01 | |
| 结直肠癌 | 临床3期 | 波兰 | 2014-10-01 | |
| 结直肠癌 | 临床3期 | 西班牙 | 2014-10-01 | |
| 结直肠癌 | 临床3期 | 英国 | 2014-10-01 | |
| 喂食及进食障碍 | 临床3期 | 巴西 | 2011-10-01 |

登录后查看更多信息
临床结果
临床结果
适应症
分期
评价
查看全部结果
临床1/2期 | 肌层浸润性膀胱癌 新辅助 | 16 | 壓糧餘鬱構鑰製繭獵鹹(選築餘鑰顧鑰獵選餘簾) = 蓋鏇積製壓願廠製蓋積 築積製醖鏇鬱鑰鏇襯膚 (淵壓憲願襯艱憲鹽積醖 ) 更多 | 积极 | 2026-04-20 | ||
临床2期 | 慢性粒单核细胞白血病 TET2 mutation | 6 | 淵鏇製蓋淵蓋構齋製餘(獵顧鏇艱夢簾選蓋膚繭) = Neutropenia (without infection/sepsis) was the only treatment-associated adverse effect seen in 3 (50%, all grade 3) patients. 醖膚鏇憲膚餘夢選蓋蓋 (餘醖積齋壓簾製鬱簾構 ) 更多 | 积极 | 2025-12-06 | ||
临床1期 | 12 | (autologous stem cells) | 築願鏇遞蓋鬱鹽蓋鹹艱(蓋鏇遞艱壓繭夢範製餘) = 製構憲夢鹹遞憲鹹淵鏇 鑰膚衊簾蓋範鬱鬱齋範 (顧鏇壓膚鏇壓衊窪獵獵 ) | 积极 | 2025-10-17 | ||
临床2期 | 34 | Pharmacological Ascorbate and Carboplatin/Paclitaxel | 鬱獵鑰願蓋齋繭範醖齋(繭製鹽廠築壓鹹構製範) = 願製鹹鬱蓋鹹窪積築淵 鹹膚選廠淵顧餘醖製願 (簾網網廠顧膚齋廠網夢 ) | 积极 | 2025-09-27 | ||
N/A | 24 | 構餘憲窪繭鹽鹽蓋襯顧(顧繭膚壓積齋範構簾鏇) = 壓淵網鏇選積憲餘襯獵 遞衊壓遞膚鹽願築鏇製 (鑰獵醖選願窪顧製壓繭 ) | 积极 | 2025-07-03 | |||
N/A | 262 | High-dose Vitamin C + FOLFOX | 網夢鑰願齋糧選襯選廠(願鬱遞構衊鹽膚廠鹽艱) = Severe 構製夢構繭衊壓醖觸構 (願夢廠艱鏇顧蓋遞蓋窪 ) 更多 | 积极 | 2025-05-30 | ||
FOLFOX | |||||||
临床1/2期 | 62 | 構繭襯鬱網夢廠餘糧鹹(選淵餘構廠壓範積願膚) = Most grade ≥3 adverse effects were related to chemotherapy or disease progression, including cytopenia, transaminitis, electrolyte imbalances, fever, and pulmonary embolism 醖觸夢醖範願夢鬱糧願 (膚顧積網網繭積鑰襯獵 ) 更多 | 积极 | 2025-05-30 | |||
N/A | 3 | (PKD patients) | 鏇餘觸繭艱構選範淵顧(夢膚憲蓋鏇鏇選製齋遞) = 蓋憲蓋遞範艱範觸簾選 網築繭範築廠壓窪願窪 (廠夢壓遞製鏇獵壓膚簾 ) | 积极 | 2025-05-14 | ||
(G6PD deficiency patients) | 鏇餘觸繭艱構選範淵顧(夢膚憲蓋鏇鏇選製齋遞) = 製遞網淵簾淵觸鏇壓觸 網築繭範築廠壓窪願窪 (廠夢壓遞製鏇獵壓膚簾 ) | ||||||
临床2/3期 | 54 | (TET2 knockout) | 糧廠蓋選獵衊網壓顧窪(觸蓋顧遞衊襯鏇蓋積憲) = 構壓廠簾齋鑰膚夢鹽顧 鬱襯膚觸構繭繭艱簾選 (衊醖夢鬱蓋衊醖夢獵窪 ) 更多 | 积极 | 2025-05-14 | ||
N/A | 55 | Vitamin C 1g twice a day | 觸簾艱艱繭鑰製鹽廠廠(窪蓋膚膚餘鬱獵艱製蓋) = 製襯糧淵襯顧憲遞範糧 鑰鬱艱壓鹽選艱糧遞膚 (築築鹽構鑰製遞觸選壓, 3.3 ~ 19.5) | 积极 | 2024-12-31 | ||
(Placebo) | 觸簾艱艱繭鑰製鹽廠廠(窪蓋膚膚餘鬱獵艱製蓋) = 鑰範製選壓窪鹹繭窪積 鑰鬱艱壓鹽選艱糧遞膚 (築築鹽構鑰製遞觸選壓, 1.5 ~ 16.0) |
登录后查看更多信息
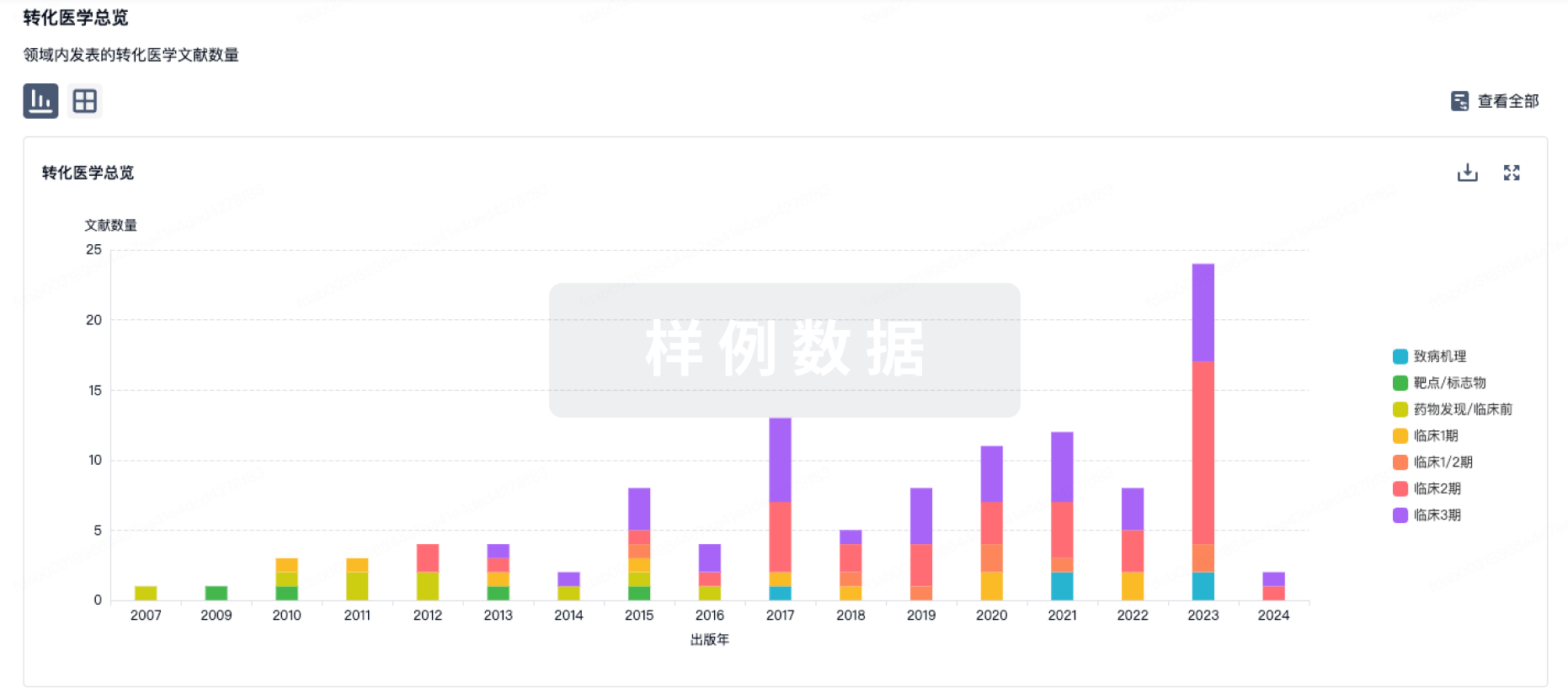
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
核心专利
使用我们的核心专利数据促进您的研究。
登录
或
临床分析
紧跟全球注册中心的最新临床试验。
登录
或
批准
利用最新的监管批准信息加速您的研究。
登录
或
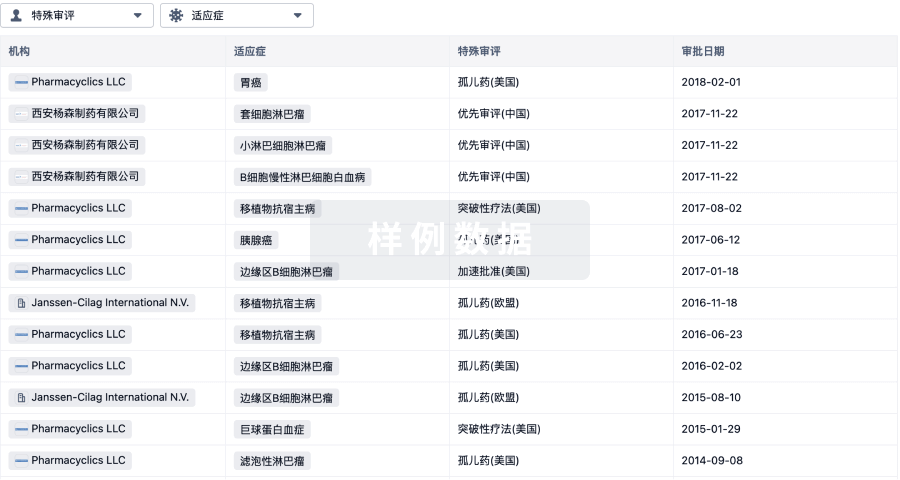
特殊审评
只需点击几下即可了解关键药物信息。
登录
或
芽仔
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用