预约演示
更新于:2026-06-17
Fudan University
更新于:2026-06-17
概览
标签
肿瘤
消化系统疾病
皮肤和肌肉骨骼疾病
小分子化药
蛋白水解靶向嵌合体(PROTAC)
诊断用放射药物
疾病领域得分
一眼洞穿机构专注的疾病领域

技术平台
公司药物应用最多的技术
靶点
公司最常开发的靶点
关联
靶点 |
作用机制 |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症 |
最高研发阶段 |
首次获批国家/地区 |
首次获批日期 |
靶点 |
作用机制 |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症 |
最高研发阶段 |
首次获批国家/地区 |
首次获批日期 |
靶点 |
作用机制 |
在研机构 |
原研机构 |
在研适应症 |
非在研适应症 |
最高研发阶段 |
首次获批国家/地区 |
首次获批日期 |


NCT06196671
Oncolytic Virus Plus PD-1 Inhibitor to Patients With Advanced Pancreatic Cancer
NCT07423611
A Multicenter, Open-label, Randomized Controlled Clinical Study Comparing the Efficacy and Safety of ctDNA-guided Ribociclib Plus Endocrine Therapy Versus Chemotherapy Followed by Ribociclib Plus Endocrine Therapy as Adjuvant Treatment in Patients With HR+/HER2- Early Breast Cancer
NCT07619911
SHR-1701 for Induction and Consolidation Therapy in Unresectable Stage III Non-Small Cell Lung Cancer: A Randomized, Phase II Clinical Trial
100 项与 复旦大学 相关的临床结果
登录后查看更多信息
登录后查看更多信息
2026-12-31Future Science OA
Clinical efficacy of 2-week and 3-week albumin-bound paclitaxel therapy for advanced pancreatic cancer
Article
作者: Su, Zhimin ; Li, Zhiyong ; Shen, Feng ; Jiang, Jiana
OBJECTIVE:
We aim to compare the clinical efficacy and safety of albumin-bound paclitaxel (nab-PTX) (125 mg/m2, q2w) for two weeks and (125 mg/m2, d1, d8, q3w) for three weeks in the first-line treatment of advanced pancreatic cancer.
METHODS:
The medical records of patients with advanced pancreatic cancer who received nab-PTX for 2 weeks and 3 weeks, combined with gemcitabine, from July 2018 to January 2023, were retrospectively analyzed. The efficacy and adverse reactions of the two groups of patients were compared.
RESULT:
A total of 64 patients were included. The median progression-free survival (mPFS) of the 2-week group was 5.6 months, while the 3-week group was 7.8 months. The median overall survival (mOS) of the 2-week group was 14.0 months, while the 3-week group was 14.7 months. The multivariate analysis showed that a physical fitness status score of 0-1 was an independent factor with better OS, while metastatic sites ≥ 3 were related to poor OS. The incidence of leukopenia or neutropenia, neurotoxicity, fatigue, and poor appetite in the 2-week group was lower than that in the 3-week group.
CONCLUSION:
The clinical efficacy of nab-PTX in the 2-week and dose intensive 3 week did not show significant difference, but the 2-week treatment group had better tolerance and safety.
2026-12-31Human Vaccines & Immunotherapeutics
Genomic comparison reveals single-nucleotide polymorphic sites and attenuation-associated site combinations specific to Chinese live attenuated varicella-zoster virus vaccines
Article
作者: Song, Minghui ; Zhang, Yuchan ; Wang, Xuanyi ; Cai, Yuanyuan ; Wang, Ziqiang ; Duan, Xuhua ; Shao, Hong ; Qiu, Tianyi ; Wang, Mingren
The live-attenuated varicella-zoster virus (VZV) vaccine, mainly based on the Oka strain, is essential for the prevention and control of chickenpox (varicella). Although genomic profiles of internationally prevalent Oka vaccine strain (vOka)-derived preparations have been reported, the comprehensive single-nucleotide polymorphism (SNP) profiles of multiple commercially available Chinese vOka-derived vaccines are still lacking. We performed next-generation sequencing (NGS) on five Chinese-manufactured VZV vaccines, comparing genomic variations against the wild-type parental Oka (pOka) strain and internationally prevalent vaccine products. Notably, among 54 key SNPs distinguishing Chinese vaccines from the wild-type pOka strain, 4 novel China-specific SNPs were identified. What's more, we identified 6 near-fixed (the vaccine allele frequency >90%) non-synonymous SNPs that consistently co-occurred across all production batches, potentially impacting protein function through amino acid substitutions. Genetic heterogeneity across manufacturers revealed manufacturer-specific signatures, whereas consistent batch production underscored manufacturing stability. By systematically mapping SNPs in Chinese VZV vaccines, this study bridges a critical knowledge gap and provides essential insights into manufacturer-specific genomic signatures.
2026-12-31Emerging Microbes & Infections
Natural history of latent brucellosis and diagnostic performance of B-SPOT: a 24-month prospective cohort study based on large-scale screening in northern China
Article
作者: Sun, Binhuang ; Zhang, Wenhong ; Wang, Hongyu ; Wang, Yan A ; Guo, Wei ; Cai, Jianpeng ; Yuan, Guanmin ; Zhao, Jingjing ; Liu, Hongyan ; Xue, Quanlin ; Chen, Yutong ; Li, Dan ; Wang, Yan O ; Liu, Xiaokun ; Sun, Yuhan ; Sun, Yulei ; Ai, Jingwen
Whether latent brucellosis exists prior to symptom onset and predisposes individuals to active disease remains to be determined. We established a novel ELISpot method for brucellosis (B-SPOT) based on host-specific immune. Diagnostic efficacy was evaluated in a testing cohort comprising brucellosis and non-brucellosis participants. A large-scale screening, using serum agglutination test (SAT) and B-SPOT for latent brucellosis, was later performed in Kangping, a brucellosis-endemic area in northern China, followed by a 24-month prospective cohort study to observe the natural development of latent brucellosis without any intervention. Among 41 brucellosis and 57 non-brucellosis participants, the sensitivity of B-SPOT for brucellosis was 95.1%, and the specificity was 98.2%, compared to the combination of blood culture and SAT. 2459 residents were screened, and the prevalence of latent brucellosis was 3.3% (81/2459). Among 63 latent brucellosis cases successfully followed, 33.3% (21/63) progressed to acute brucellosis within 2 years, while none in the control group progressed(p < 0.001). 16/21 (76.2%) progressed within the first six months, and 14/21 (66.7%) presented with joint pain as their initial symptom. B-SPOT (10.94[95%CI 2.48-48.33, p = 0.002]) outperformed SAT (3.04[95%CI 1.14-8.01, p = 0.026]) in predicting the progression of latent brucellosis. Latent brucellosis accounts for 3.3% in the endemic area, among whom 33.3% progressed to acute brucellosis within two years. The first six months are a critical window, and joint pain is the primary initial symptom. B-SPOT demonstrates high diagnostic efficacy for brucellosis disease activity and can better predict the progression of latent brucellosis.
100 项与 复旦大学 相关的药物交易
登录后查看更多信息
100 项与 复旦大学 相关的转化医学
登录后查看更多信息
组织架构
使用我们的机构树数据加速您的研究。
登录
或

管线布局
2026年07月26日管线快照
管线布局中药物为当前组织机构及其子机构作为药物机构进行统计,早期临床1期并入临床1期,临床1/2期并入临床2期,临床2/3期并入临床3期
药物发现
46
258
临床前
临床申请批准
3
9
临床1期
临床2期
9
1
临床3期
其他
41
登录后查看更多信息
当前项目
登录后查看更多信息
药物交易
使用我们的药物交易数据加速您的研究。
登录
或
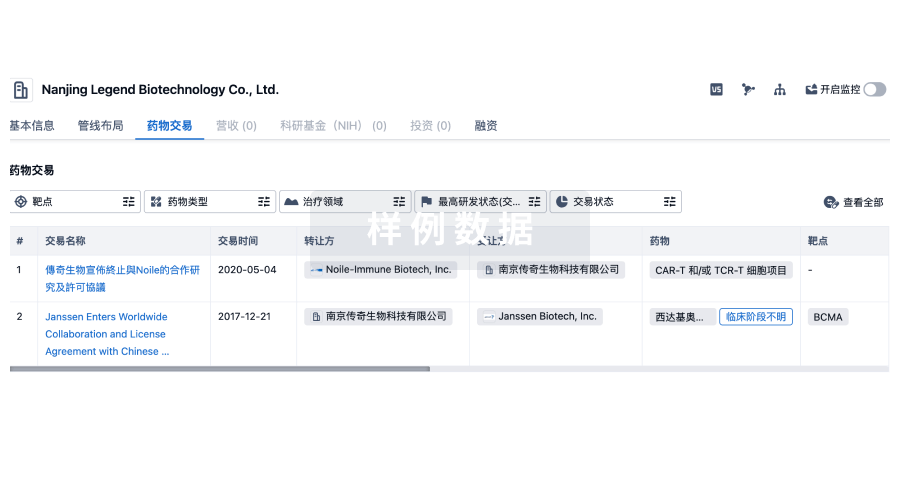
转化医学
使用我们的转化医学数据加速您的研究。
登录
或
营收
使用 Synapse 探索超过 36 万个组织的财务状况。
登录
或
科研基金(NIH)
访问超过 200 万项资助和基金信息,以提升您的研究之旅。
登录
或
投资
深入了解从初创企业到成熟企业的最新公司投资动态。
登录
或
融资
发掘融资趋势以验证和推进您的投资机会。
登录
或
芽仔
全新生物医药AI Agent 覆盖科研全链路,让突破性发现快人一步
立即开始免费试用!
智慧芽新药情报库是智慧芽专为生命科学人士构建的基于AI的创新药情报平台,助您全方位提升您的研发与决策效率。
立即开始数据试用!
智慧芽新药库数据也通过智慧芽数据服务平台,以API或者数据包形式对外开放,助您更加充分利用智慧芽新药情报信息。
生物序列数据库
生物药研发创新
免费使用
化学结构数据库
小分子化药研发创新
免费使用